生物大分子结构与功能的示踪研究
生物大分子是聚合物,由许多小分子聚合而成,其功能不仅取决于所含的成分,而且取决于小分子的排列及整个分子的构型。所以生物大分子的研究技术与一般小分子的研究有很大不同。核素示踪技术在生物大分子结构与功能的研究中有很重要的作用,实验设计上也与小分子的研究有很大的不同。
生物大分子酶促合成基本条件的研究 核素参入试验是测定生物大分子合成速度的一种有效方法。通常是在被研究的体系(整体动物,离体细胞或无细胞培养物)中加入特定的标记小分子前身物(蛋白质用氨基酸,RNA用尿嘧啶核苷或尿嘧啶单核苷酸,DNA用胸腺嘧啶核苷或胸腺嘧啶单核苷酸),经过一定时间后分出相应的大分子,测放射性。合成速度越快,标记物参入越多,放射性越高。
利用这种参入试验,可以在试管中人为地改变各种因素,研究各种生物大分子酶促合成需要那些必不可少的基本条件。例如用无细胞体系研究DNA的合成,发现试管中必须包含DNA聚合酶、四种单核苷酸、镁离子;此外,还必须有少量DNA的存在。而单核苷酸以含三个磷酸根者最为有效,含二个磷酸根者次之。由此得出结论,DNA的合成与一切小分子化合物不同,必须先有一些DNA的存在(后来发展成“模板学说”);其直接原料则是含三个磷酸根的单核苷酸。
类似的方法曾用于研究蛋白质及RNA的合成条件;此外还曾证明,某些病毒含有一种逆转录酶,在这种酶的催化下,合成DNA不需要预先有DNA的存在,而需要RNA的存在,即所谓“逆转录”现象。
半保留模型的证实 DNA的合成需要先有一些DNA分子的存在,这些先存在的DNA分子的作用及其与新合成的DNA分子之间的关系,都是通过用15N标记的氯化铵进行示踪实验才阐明的。
在试管中培养大肠杆菌,培养基中除15NH4Cl外不含其他含氮化合物。经多代繁殖,细菌的DNA都含有大量15N,在密度梯度离心时特别重,集中在靠近试管底部的一个区带。随后将细菌移入含14NH4Cl的培养基中再繁殖一代,发现这时的DNA在密度梯度离心时其重量介于重DNA和普通DNA之间。用加热法把双链折开再离心,则这种中间密度的DNA不再存在,而出现一半是重的单链,一半是轻的单链。这表明,新形成的中间密度的DNA双链中,一条单链来自上一代的重DNA。另一条则是以14N为原料新合成的,故称之为杂化DNA。再培养一代,离心分析可见到杂化DNA和轻DNA各占一半。
上述实验证明,DNA的合成是“半保留”性质,即双链折开,各自进入一个新的DNA分子。也就是说,DNA分子的复制是保留一半,合成一半,保留的一半在好多代中都保持完整而不断裂。
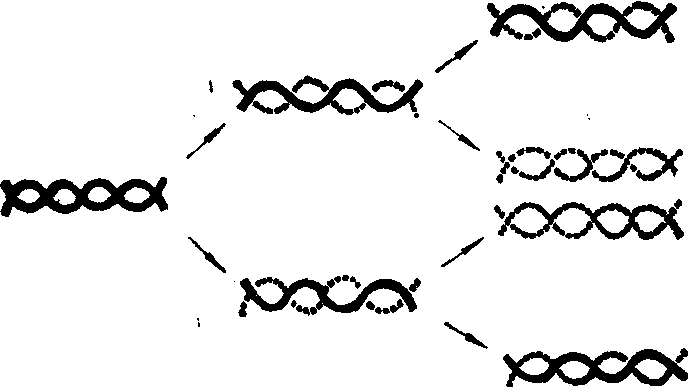
图1 DNA的半保留复制模型
实线为含15N的重链 虚线为不含15N的轻链
最近邻序列分析 DNA彻底水解及结构分析的结果表明,四种碱基(胸腺嘧啶、胞嘧啶、腺嘌呤、鸟嘌呤,分别用T、C、A、G表示)是通过脱氧核糖及磷酸互相连接的。由此可以设想,不同DNA所以有不同特性是由于碱基排列各有一定顺序。最近邻序列分析法就是为了证实碱基排列确有特殊顺序而设计的。
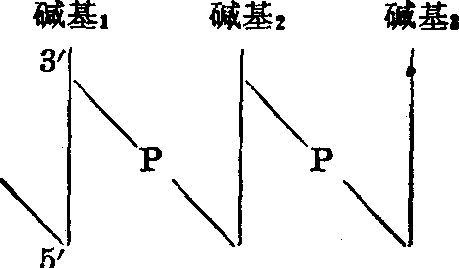
图2 DNA分子中碱基的连接方式
碱基下的垂直线代表脱氧核糖,3'、5'表示核糖的碳原
子编号
如果用四种脱氧核苷三磷酸(连于5'C)为原料,而且其中有一种用32P标记在α位,在试管中合成DNA,再用一种特殊的酶打开5’C—P键,使DNA水解成3'C上带磷酸根的单核苷酸,则22P将从原来连接的核苷酸上移到最近邻的另一单核苷酸上,以22P-ATP为前身物的情况见图3。
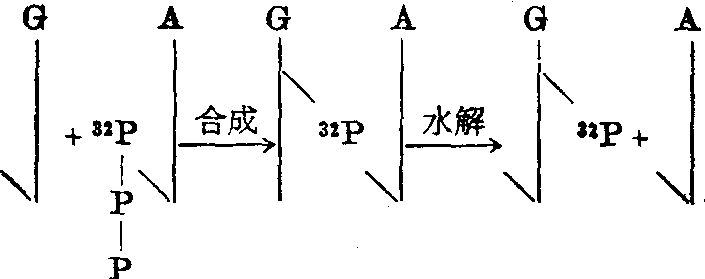
图3 最近邻序列分析中32P的移位
所以这一实验方法可以判断每种核苷酸的最近邻中T、A、C、G的比例。正式实验分四批进行,每批用一种32P标记的脱氧核苷三磷酸,总的实验结果可以知道每种单核苷酸的最近邻中T、A、C、G的比例。如果DNA分子中四种碱基的排列没有特定顺序,而是随机的,则C的最近邻为G的几率应与G的最近邻为C的几率相同,其他A-T和T-A,C-A和A-C,G-T和T-G等也应如此。但实验结果并非如此。在小牛胸腺DNA的分析中,C-G占1.6%,而G-C占4.4%;在草分枝杆菌的分析中,A-T占1.2%,T-A占3.1%。所以最近邻序列分析证明,DNA分子中四种碱基的排列并非随机的,而是有特定的顺序。此外,实验结果还表明,不同来源的DNA所得最近邻的比例不同,同一来源的DNA和以此DNA为模板复制的DNA具有相同的最近邻比例。目前,最近邻序列分析法对区别不同来源的核酸仍是一种有实用价值的方法。
分子杂交技术 参见“核酸分子杂交技术”条。
14C-氨基酸结合法 蛋白质的结构分析表明,分子中氨基酸的排列决定蛋白质的生物特性。为了阐明RNA中的遗传信息如何通过核苷酸的排列顺序决定蛋白质中氨基酸的排列顺序,Nirenberg等设计了被称为14C-氨基酸结合法的实验。先人工制备64种三核苷酸(按一定顺序排列)及20种14C标记或非标记的氨基酸-sRNA(即tRNA,转运核糖核酸)。以大肠杆菌的蛋白质合成酶为催化剂,在体外作参入试验,测定14C-氨基酸参入核朊微粒的量。根据设想,每种三核苷酸应当决定一种氨基酸参入蛋白质,亦即是该氨基酸的“密码”。所以每次实验用一种三核苷酸,用一种14C-氨基酸-sRNA及十九种非标记氨基酸-sRNA。许多次实验的结果确实证明,每种14C-氨基酸只有在某一种或几种特定三核苷酸存在时才参入核朊微粒(见表)。根据上述实验,现已基本公认,蛋白质分子上氨基酸的排列顺序是由mRNA(信使核糖核酸)上核苷酸的排列顺序决定的,而且基本上是每三个核苷酸决定一个氨基酸。
蛋白质的末端分析 蛋白质的羧基末端在醋酐作用下能环化,在碱性条件下又会开环,开环时需加入一分子水,如果反应物中有氚水,氚就参入到分子中。以后将蛋
决定蛋白质中氨基酸排列顺序的三核苷酸——遗传密码
| 第二核苷酸 第一核苷酸 | U | C | A | G | 第二核苷酸 第三核苷酸 |
| U | 苯丙 苯丙 亮 亮 | 丝 丝 丝 丝 | 酪 酪 结束肽链 结束肽链 | 半 胱 半 胱 结束肽链 色 | U C A G |
| C | 亮 亮 亮 亮 | 脯 脯 脯 脯 | 组 组 谷 酰 谷 酰 | 精 精 精 精 | U C A G |
| A | 异亮 异亮 异亮 蛋 | 苏 苏 苏 苏 | 门冬酰 门冬酰 赖 赖 | 丝 丝 精 精 | U C A G |
| G | 缬 缬 缬 缬 | 丙 丙 丙 丙 | 门冬 门冬 谷 谷 | 甘 甘 甘 甘 | U C A G |
注:A——腺嘌呤,C——胞嘧啶,G——鸟嘌呤,U——尿嘧
啶。丝表示丝氨酸,色表示色氨酸,余类推
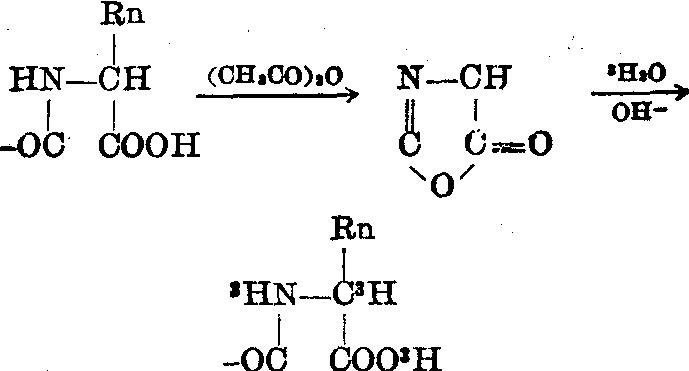
图4 用氚水作蛋白质羧基末端分析的反应历程
核糖核酸的末端分析 当核糖核酸的3′末端为核苷时,该核苷的核糖上C′2及C′3两个顺位羟基均呈游离状态,可用过碘酸钠氧化成双醛,再用NaB3H4还原成双醇,从而形成3H标记物,降解后可用层析法分离,通过测放射性作出鉴定。
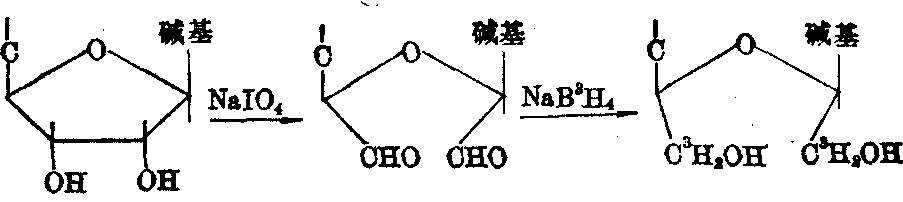
图5 核糖核酸3′末端分析的反应历程(NaB3H4标记法)
如核酸的3′末端带有一个磷酸单酯,则可用14CH2OH等进行标记,再水解鉴定。
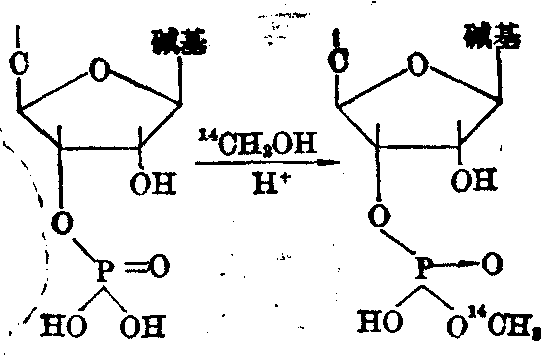
图6 用14CH3OH标记RNA 3′末端的磷酸单酯
许多RNA的末端有5′-磷酸单酯。14C-酰替吗啉磷酸甲酯能特异地标记5′-磷酸单酯,降解后分离测放射性,可判定5′末端的核苷酸。

图7 用14C-酰替吗啉磷酸甲酯标记RNA的5′末端
用“加减法”进行DNA序列分析 加减法是一种新的DNA序列分析方法,快速而简便,一次能分析多至200个碱基的顺序。
DNA的合成是以单链DNA为模板,以四种脱氧核苷三磷酸(dATP、dCTP、dGTP、dTTP)为原料,按碱基配对的原则,从起始点逐一接上去的。如果在系统中减去一种原料,则新合成的链将停止在需要这一原料接上去的位置。这就是减法分析的依据。实际做法是先在四种原料齐全的条件下合成一段新链,时间不同,长度也不同。四种原料中至少选一种用32P标记,使新链全部带放射性,以便最后用放射自显影定位。然后在缺一种原料的条件下继续合成过程,使不同长度的新链继续延长,直到需要该种原料的位置因无原料而中止。最后用凝胶电泳法分离不同长度的新链,并用放射自显影定位,这些位置的后面一位就是该所缺原料碱基的位置。
如果在新链合成到某一长度后转移到只有一种原料的条件下,并在试管中加一种DNA聚合酶,这种酶有两个作用,一是能使DNA链延长,一是能从末端开始逐一降解核苷酸。于是原来不同长度的新链都将逐步被降解,但降解到某种碱基时,由于有大量同种碱基的存在,就不再降解(实际上是一边降解,一边重新合成)。最后在凝胶电泳的放射自显影上显示的新链位置,都是上述一种原料碱基的位置。
四组减法系统和四组加法系统 (分别缺一种碱基或加一种碱基)的结果综合起来,就可一次得到相当长一段新链的碱基排列顺序。
生物大分子结构与功能的研究正在不断深入,新的示踪研究方法也将不断踊现。
- 冒调是什么意思
- 冒谋天光是什么意思
- 冒谋天尊是什么意思
- 冒谋打瓜是什么意思
- 冒谷井是什么意思
- 冒豆干是什么意思
- 冒货是什么意思
- 冒贿是什么意思
- 冒赈是什么意思
- 冒赏是什么意思
- 冒起胆子是什么意思
- 冒越是什么意思
- 冒路德节是什么意思
- 冒躁是什么意思
- 冒车是什么意思
- 冒辟疆是什么意思
- 冒进是什么意思
- 冒遁是什么意思
- 冒酸水是什么意思
- 冒里冒失是什么意思
- 冒里冒突是什么意思
- 冒里失砍是什么意思
- 冒金星是什么意思
- 冒金星儿是什么意思
- 冒金花是什么意思
- 冒钱是什么意思
- 冒销是什么意思
- 冒锋是什么意思
- 冒闷是什么意思
- 冒阳是什么意思
- 冒险是什么意思
- 冒险主义是什么意思
- 冒险主义者是什么意思
- 冒险借贷是什么意思
- 冒险倾向的性别差异是什么意思
- 冒险偷婚是什么意思
- 冒险公司是什么意思
- 冒险家是什么意思
- 冒险小说是什么意思
- 冒险履危是什么意思
- 冒险干坏事的人必将自食其恶果是什么意思
- 冒险性投资组合是什么意思
- 冒险投资是什么意思
- 冒险故事是什么意思
- 冒险故事作家是什么意思
- 冒险犯难是什么意思
- 冒险生活是什么意思
- 冒险的事情是什么意思
- 冒险的军事活动是什么意思
- 冒险穿越是什么意思
- 冒险精神是什么意思
- 冒险行事是什么意思
- 冒险行进是什么意思
- 冒险规谏是什么意思
- 冒险触难是什么意思
- 冒险记幸 杨绛是什么意思
- 冒险资本是什么意思
- 冒险转移是什么意思
- 冒险钻空子是什么意思
- 冒隙乘虚是什么意思