真核细胞的起源
多年来人们一直把动物与植物两大类视为生物界最基本的划分。虽然早在细胞学说建立时人们就已经认识到动物和植物同样都是由细胞构成的,但是人们在很长时间内仍然把动植物的划分当作是最基本的,只是由于本世纪50年代以后细胞生物学和分子生物学的巨大发展,才使人们终于承认真核生物与原核生物的划分是更为重要的。
“细胞”一词原来是专指具有真正的细胞核、线粒体和内质网等一系列细胞器的真核细胞而言的,但是后来细胞的概念把原核生物也包括在内,特称为原核细胞,本来意义的细胞则被称为真核细胞。真核生物泛指一切由真核细胞及其衍生物所构成的生物,原核生物则包括两大类,即直到70年代后期方才真正有所认识的原细菌类(Archaebacteria) 和与之相对立的真细菌类 (Eubac-teria)。后者包括绝大多数的细菌、蓝藻、放线菌、支原体、衣原体等等。
真核细胞的特征 对比真核细胞与原核细胞可以看到这样一些差异:
❶大小的不同。
❷有无真正的细胞核。
❸分裂方式的不同。
❹核糖体的不同。
❺有无线粒体、高尔基体等细胞器。
❻鞭毛结构的不同。
❼有无胞质环流现象。
还有一些方面过去也曾被认为是真核细胞与原核细胞的重大差别,但是后来却被科学的发展所否定了。例如过去曾长期认为原核细胞的染色质中不含碱性蛋白,并认为这是它与真核细胞根本的不同。但是后来发现在真细菌类的染色质中普遍含有一种被称为Hu蛋白的碱性蛋白与DNA结合在一起。以后在各种原细菌体内也发现有不同的碱性蛋白与DNA相结合着。过去曾把壁中有无含二氨基庚二酸与胞壁酸的肽聚糖视为原核细胞的根本差异。但是后来发现原细菌也像真核细胞一样不含肽聚糖,肽聚糖的存在其实只是真细菌类的标志。又如过去曾以为在蛋白质合成的起始上真核细胞与原核细胞是不同的,原核细胞用的是甲酰蛋氨酰tRNAimet,而真核细胞用的是蛋氨酰tRNAimet。但是后来发现其实只是真细菌类用甲酰蛋氨酰tRNAimet,原细菌类所用的实际也是蛋氨酰tRNAimet。类似的事例还可以举出许多。
在上面已经肯定了的真核细胞与原核细胞之间的差别中有些其实也只是相对的。例如有些细菌就达到了原生动物的大小,如大卵硫细菌(Thiovulummajus)长11—8μm,粗8—14μm;草酸无色菌 (Achromatiumoxali-ferum)长33—35μm,粗25—28μm。原细菌的核糖体虽然在大小上与真细菌的相似而与真核细胞的显著不同,但是在它们的细微形态上,在rRNA分子和核糖体蛋白分子的一级结构上,却往往更近似于真核细胞的。线粒体、高尔基体、叶绿体等细胞器也并不是所有的真核细胞都有的,有些寄生在消化道中的原生动物就没有线粒体和高尔基体。反之,大卵硫细菌却具有类似于内质网的结构。看起来,真核细胞不同于原核细胞的最根本点在于具有真正的细胞核。
真核细胞的祖先问题 真核细胞与原核细胞在基本的生命活动机制上是高度一致的,它们具有共同的遗传密码系统,在DNA、RNA和蛋白质的合成方式上也是相同的。因此它们的同源性是没有疑问的。从它们的复杂程度和在地球上出现的先后来看,也只能是真核细胞起源于古代的某种原核生物,而不是相反。
在原细菌类的种种特性尚未为人知晓的时候,曾经有人提出过真核细胞起源于真细菌类中的蓝藻的设想。其主要根据是真核细胞的叶绿体与蓝藻的明显相似。但是真核细胞的本体,即除去叶绿体和线粒体以外的部分,在分子生物学和细胞生物学的种种特性上都是距离真细菌类相当远的。至于叶绿体和线粒体与真细菌类的相似性,其实,用它们的内共生起源学说就可以给予更合理的说明(见“线粒体与叶绿体的内共生起源学说”条)。
通过70年代末以来对原细菌类所作的多方面研究,如今已经知道原细菌类在一系列的分子生物学特性上都是近似于真核细胞,而距离真细菌类却相当远。例如原细菌类的壁像真核细胞的一样,并不含有真细菌壁中普遍含有的二氧基庚二酸和胞壁酸构成的肽聚糖层。各类真核生物的核DNA中普遍含有多量的高度重复序列,这是真细菌类的DNA中所没有的,但是已经在某些原细菌类的DNA中被发现了。在分子遗传学上真核细胞与真细菌类的重大区别之一是它们结构基因中有内含子存在,这是真细菌类所没有的。现在也已经在几种原细菌的tRNA基团和23SrRNA基团中找到了内含子。现已知各类真核生物的DNA复制酶都对头孢霉(Cephalos-porium aphidicola)所产生的抗生素aphidicolin很敏感,真细菌类的则否。现在已经发现原细菌类的几种极端嗜盐细菌的DNA复制酶也是对aphidicolin非常敏感的。对原细菌类的依赖于DNA聚合酶进行的多方面研究结果,一致表明原细菌类的与真核细胞的RNA聚合酶是相当近似的,而真细菌类的酶则距离它们两者之间的全都很远。真细菌类的RNA聚合酶仅由4种亚基构成,真核细胞的则由10种或更多的亚基构成。原细菌类中的产甲烷细菌的RNA聚合酶由7至9种亚基构成,而依赖硫的嗜高温原细菌的则由10种以上的亚基构成。后者的RNA聚合酶特别相似于真核细胞中负责核糖体RNA之转录的RNA聚合酶Ⅰ。免疫化学的交叉反应表明,原细菌与酵母菌的RNA聚合酶Ⅰ的相应亚基之间的交叉反应相当强,其强度甚至显著地超过了酵母菌的酶Ⅰ与负责mRNA之转录的酶Ⅱ的相应亚基之间的反应。而原细菌的或真核细胞的与真细菌类的此种酶的相应亚基之间的交叉反应则都很弱。它一方面说明三者的酶有同源性,同时又说明原细菌的与真核细胞的相距比较近,而它们距离真细菌的则相当远。已知各种真细菌的RNA聚合酶,包括蓝藻的在内,全能被利福平或链霉溶菌素所抑制,而真核细胞的以及原细菌类的酶则都对它们不敏感。与此相对照的是,从蓟(Silybiummarianum)提取出来的蓟素会促进真核细胞的RNA聚合酶Ⅰ的转录活动,但是对真核细胞的RNA聚合酶Ⅱ以及真细菌类的RNA聚合酶则没有作用。现在已知蓟素对原细菌类的RNA聚合酶也同样有促进作用。
原核细胞的核糖体虽然像真细菌类的一样在大小上是70S型的,但是它们构成核糖体的诸蛋白分子的结构却往往明显地更相似于真核细胞。在它们的核糖体中与50SrRNA相结合的蛋白以及核糖体蛋白中酸性最强的“A蛋白”等等就都是如此。这方面已经作了不少的研究。与这一点相关的是,许多能专一地结合到真细菌类的核糖体中各自特定的蛋白分子上从而抑制真细菌类蛋白合成活动的抗生素,如氯霉素、链霉素、红霉素等,不仅对真核细胞的核糖体不起作用,对原细菌类的核糖体也同样不起作用。反之,能专一地结合到真核细胞核糖体大亚基上去的茴香霉素虽然对真细菌类的蛋白合成不起作用,却能结合到原细菌类的核糖体大亚基上去,抑制它们的蛋白质合成。
有关原细菌核糖体的一项最新的重要发现是:原细菌、真细菌与真核细胞的核糖体在形态上,真细菌的与真核细胞的有明显的差异,而原细菌的与真核细胞的却是相近的,特别是依赖硫的嗜高温原细菌(图1)。后者的核糖体在蛋白组成上也比真细菌和其他原细菌类的要复杂得多,从而更近似于真核细胞。
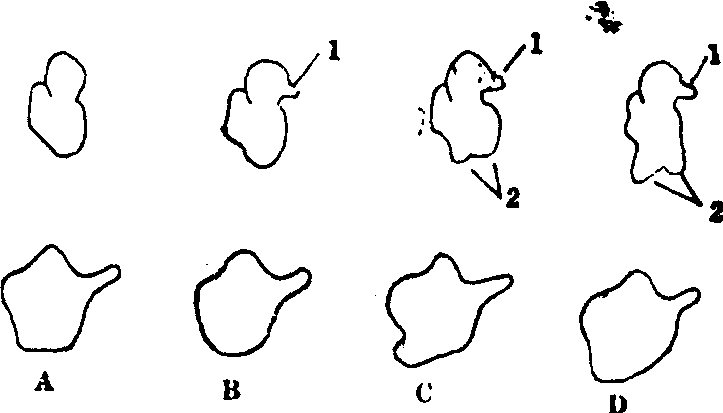
图1 核糖体的形态比较
A.真细菌,一种蓝藻;B.极端嗜盐原细菌;C.依赖硫的嗜高温原细菌; D.真核生物,一种酵母菌; 1.“喙”; 2. “叶”;
上一列示核糖体小亚基,下一列示大亚基。“喙”在两类原细菌即已出现; “叶”在依赖于硫的原细菌也开始出现,在真核生物则很发达
已知在蛋白质合成的起始端,所有的真细菌,包括蓝藻,用的都是甲酰蛋氨酰tRNAimet,而真核细胞与原细菌类所用的则都是蛋氨酰tRNAimet。真细菌类体内有一种专门为甲酰化蛋氨酰tRNA用的酶,这种酶原细菌类与真核细胞都没有。
已知白喉毒素对所有的真核生物的蛋白合成全都有强烈的抑制作用,然而对所有的真细菌类的蛋白合成则全都没有作用。这是由于白喉毒素能够作用于真核细胞的“肽链延长因子Ⅱ”分子上的一个特定段落,它在真细菌的相应分子上是没有的。用原细菌类作试验所得的结果表明,白喉毒素对它们的蛋白合成也同样有强烈的抑制作用。
有人检查了几种真细菌与真核细胞的氨基酰tRNA合成酶各自对真细菌类、原细菌类与真核生物tRNA的作用能力,结果清楚地表明:真核细胞的tRNA和原细菌类的tRNA是一类,真细菌类的则是另外一类。真核细胞的酶对真细菌类tRNA的作用能力很弱,对原细菌的作用能力要强得多;反之,真细菌类的酶对于真核生物的和原细菌tRNA的作用能力则全都很弱。
类似的证据还有许多。现存的原细菌分为两大支,产甲烷细菌和极端嗜盐细菌为一大支,依赖硫的嗜高温细菌是另一大支。后者在一系列方面都比前者更近似于真核细胞,因而可以设想真核细胞的祖先是在远古时代从这一大支中发源的; 真细菌类则有可能是从前一大支起源的(图2)。特别有意思的一点是产甲烷细菌也会受到噬菌体的危害,而危害依赖硫的嗜高温原细菌的却不是噬菌体而是病毒。真核细胞也是如此。
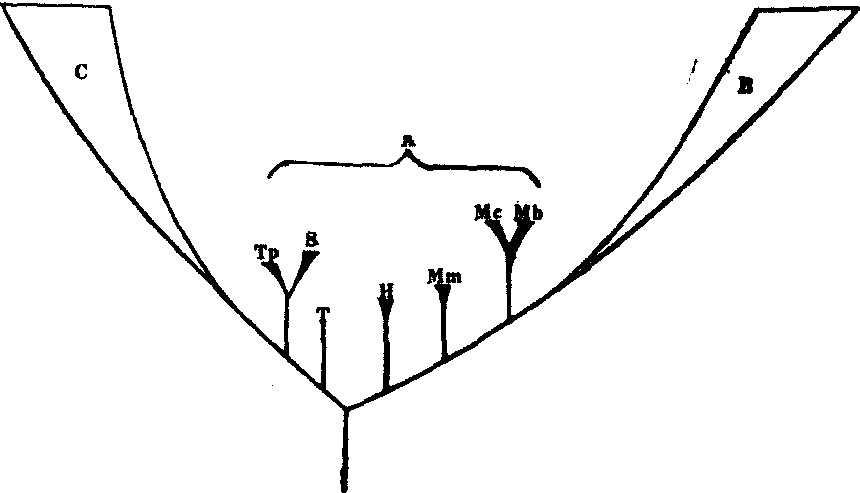
图2 一种可能的生物进化系统树
A. 原细菌;Mb. 产甲烷杆菌类;Mc. 产甲烷球菌类;Mm.产甲烷微球菌类; H. 盐细菌类; T.热原质体类; S.硫氧化菌类; Tp.热变形菌类; B. 真细菌;C.真核生物
真核细胞进化途径 依据目前已知的事实,可以设想古代原细菌类中的某一小支在进化过程中长得很大,成了“前真核生物”,即最初的单细胞真核生物的直接祖先。体积加大,相对于单位体积的表面积就会缩小。如果此时相对于单位体积的原生质的细胞膜面积也随着缩小,积极的代谢就将无法保持。为此它们的细胞膜发生内褶,从加大细胞膜的总面积,保证细胞膜的相对面积不致降低,就是势所必然的了。个体长的越大,细胞膜内褶所形成的结构的作用也就越大,而这也就为与蛋白质合成相联系的原始性的内质网的形成奠立了基础。与体积长得很大相关,前真核生物应有多个重复的染色质结构。在真核细胞进化形成过程中,原始性的内质网包围住染色质结构所在的区域,而其直接邻贴此区域的部分则在功能和形态上都逐渐发生变化,以保证此区域对染色质结构的活动有着最适宜的条件。这样就形成了初始的核被膜和原始性的细胞核。与之类似的细胞核可见于现今生存的涡鞭毛虫类(甲藻)体内。在这样的细胞中,原核生物的染色质结构就成了染色体,但是后者在许多方面仍然还保留原核生物的染色质结构的特性。由原始性的细胞核后来再进化成为典型的真核。其间的一些过渡形式,也可见于现今生存的比较高等的涡鞭毛虫类。
高尔基体、溶酶体等都可以设想是起源于原始性的内质网。热原质体和硫氧化细菌等原细菌的体内已经有了接近于肌动蛋白的蛋白质,因此可以设想从原细菌演化出来的前真核生物已经具有肌动蛋白型的蛋白,能够进行胞质环流和变形运动以及吞噬活动。能够进行吞噬活动,是它们能够得到胞内共生生物的先决条件。
叶绿体(包括白色体、有色体和前质体)和线粒体看来是分别起源于在前真核生物体内(或最原始的单细胞真核生物体内)过胞内共生生活的两种不同的真细菌类生物。这两类细胞器都各有自己的染色质结构和自己的蛋白质合成系统。后者与真核细胞细胞质中的蛋白质合成系统截然不同,却是真细菌性质的。这两类细胞器都有双层的被膜,两层膜的性质是不同的,外膜相似于真核细胞的内质网膜,内膜却相似于真细菌类的细胞膜。看来,外膜在进化上是起源于宿主包围着胞内共生生物的液泡的膜,内膜才是来自于共生生物自身的细胞膜。叶绿体显然起源于胞内共生的蓝藻。目前已经发现了一些介于胞内共生蓝藻与叶绿体之间的过渡性结构。
关于线粒体的祖先,就目前已掌握的分子生物学资料来判断,应该是一种很接近于现今独立生活的反硝化副球菌 (Paracoccus denitrificans) 的胞内共生细菌。在它们转变为线粒体的过程中,身体结构也像叶绿体一样发生了很大变化,只是程度大得多。就在真核细胞已经完全形成以后,线粒体也还在继续发生变化。例如在多细胞动物体内,线粒体自身的核糖体变得很小,比真细菌类体内的核糖体小得多;反之在高等植物体内,它们却变得很大,比真细菌类体内的大得多(见“线粒体和叶绿体的内共生起源学说”条)。
曾有一些人设想线粒体是前真核生物自身分化出来的,只是后来体内只有这一小部分仍然还保留着原核生物的一系列特性,身体的其余部分全都进化成了真核细胞性的。这一类设想都有一个共同的前题,就是假定前真核生物是起源于真细菌类,否则就无从说明为什么线粒体的蛋白质合成系统是真细菌性质的。但是这一假设跟前面所列举的证明真核生物起源于古代的原细菌类的一系列分子生物学事实是全然矛盾的。
一般都设想前真核生物自身不会进行氧化磷酸化。在这种情况下,它们如果得到能进行氧化磷酸化的细菌作为胞内共生物,就会在生存斗争中占有巨大的优势。而这也就使得这些胞内共生物在后来变成了它们不可缺的成分,终于变成了线粒体。但前真核生物自身不能进行氧化磷酸化,并不意味着它们完全不能利用氧气。现今的真核细胞体内就存在着与线粒体无关的利用氧的系统,例如内质网上由细胞色素P450和细胞色素b5等构成的使脂肪酸羟化和不饱合化的系统。有人提出真核细胞所普遍具有的微体(过氢化氢体)就代表着前真核生物自身的一种非常低级的呼吸细胞器的残余。按照它们的很低级的呼吸方式,氧会与氢结合成为过氧化氢,而为了避免后者的毒性,生物在进化中产生出了过氧化氢酶以分解之。有迹象表明,这样利用氧的方式也存在于属于原细菌类的热原质体(thermoplasma)的体内。
- “𪯻”铜簋是什么意思
- “𫆅”铜鼎是什么意思
- :-”是什么意思
- ”-’是什么意思
- ”李膺门是什么意思
- ……gonx……是什么意思
- ……以南是什么意思
- ……如果你能…… [法国]亚默是什么意思
- ……年……月……日在……签订是什么意思
- ……年度财政预算……是什么意思
- …不几几的是什么意思
- …不几的是什么意思
- …不叽叽的是什么意思
- …不唧的是什么意思
- …不来是什么意思
- …了去了是什么意思
- …倒了…是什么意思
- …去啦…是什么意思
- …头…脑…是什么意思
- …头…脑;…是什么意思
- …头儿是什么意思
- …得慌…是什么意思
- …得过儿是什么意思
- …把……是什么意思
- …的过儿…是什么意思
- …着呢是什么意思
- …破…立… …是什么意思
- …长…短是什么意思
- …门事件…是什么意思
- :-″是什么意思
- ⅰ是什么意思
- ⅰ区效应是什么意思
- ⅰ型q波型急性心肌梗死是什么意思
- ⅰ型双向性心律是什么意思
- ⅰ型呼吸衰竭是什么意思
- ⅰ型封闭抗体是什么意思
- ⅰ型左前分支传导阻滞是什么意思
- ⅰ型左房心律是什么意思
- ⅰ型心房扑动是什么意思
- ⅰ型无q波型急性心肌梗死是什么意思
- ⅰ型空隙现象是什么意思
- ⅰ型糖尿病是什么意思
- ⅰ型补体受体是什么意思
- ⅰ型裂隙现象是什么意思
- ⅰ型超敏反应是什么意思
- ⅰ型错误是什么意思
- ⅰ导联是什么意思
- ⅰ导联轴是什么意思
- ⅰ.职官条目分类是什么意思
- ⅱ是什么意思
- ⅱ区效应是什么意思
- ⅱ和弦的应用是什么意思
- ⅱ型q波型急性心肌梗死是什么意思
- ⅱ型—后期膨胀型是什么意思
- ⅱ型双向性心律是什么意思
- ⅱ型呼吸衰竭是什么意思
- ⅱ型左前分支传导阻滞是什么意思
- ⅱ型左房心律是什么意思
- ⅱ型心房扑动是什么意思
- ⅱ型无q波型急性心肌梗死是什么意思