抗药性机制mechanism of resistance to insecticide
昆虫对杀虫剂产生抗药性的机制,可分为行为机制和生理生化机制两种。行为机制是受药剂刺激后,昆虫改变原来某些行为习性,对药剂的敏感性增加,避免或在未接触足够药量前就迅速迁移他处而不致死。对于行为抗药性机制研究得很少。生理生化机制主要包括:❶穿透性降低;
❷解毒代谢增强;
❸靶标部位敏感性降低等。
穿透性降低 根据不同施药方式,杀虫剂穿透昆虫表皮、肠道或气管,经过吸收、运输、分布和贮存,最后到达靶标部位而起作用。烟芽夜蛾(Heliothisvirescens Fabricius)和家蝇(Musca domestica Vicina)等抗药性昆虫,均显示杀虫剂穿透作用降低。萨维基(R . M . Sawicki,1968)等将控制此作用的基因称为“pen”。此基因的有效性取决于昆虫表皮成分、骨化程度和杀虫剂性质。穿透性降低很少单独起作用,常与其他抗药性因子相结合而增加抗药性。如果表皮穿透性降低时,杀虫剂不易进入体内,进入的药量会少,假使昆虫的解毒代谢能力强,则到达昆虫中毒靶标部位的药量会很少,不能使昆虫致死。反之,如果药剂容易穿透,而解毒代谢又弱,则进入虫体的药量大,会使昆虫死亡。因此杀虫剂穿透作用与其他抗药性机制的关系,是受“机会因子”作用的。
解毒代谢增强 杀虫剂进入虫体到达靶标部位的同时,也因体内各种解毒作用而降低毒效。多数抗药性机制都是由于解毒代谢的增强。家蝇代谢抗药性似乎是在单基因控制下,其产物为一个受体蛋白。此蛋白与杀虫剂结合,形成蛋白杀虫剂复合体,它可诱导多种解毒酶系的合成,其他昆虫体内的代谢受体蛋白尚不清楚。解毒代谢涉及的主要酶系为: 滴滴涕-去氯化氢酶(滴滴涕-dehydrochlorinase)、谷胱甘肽S-转移酶(glutathione S-transferase)、磷酸酯酶(phosphotase)、羧酸酯酶(carboxylesterase)和微粒体多功能氧化酶(microsomal mixed function oxidase,MFO)等。
滴滴涕—去氯化氢酶 可使滴滴涕除去一分子氯化氢而降解为无毒滴滴依的酶,简称滴滴涕一酶,它对滴滴涕类似物也有催化作用。此酶须有谷胱甘肽(GSH)存在时才有活性,但是谷胱甘肽并不参与酶促反应(图1)。它可为 1,1-双(4-氯苯基)乙烷(DMC)或F-DMC所抑制。滴滴涕-酶是某些滴滴
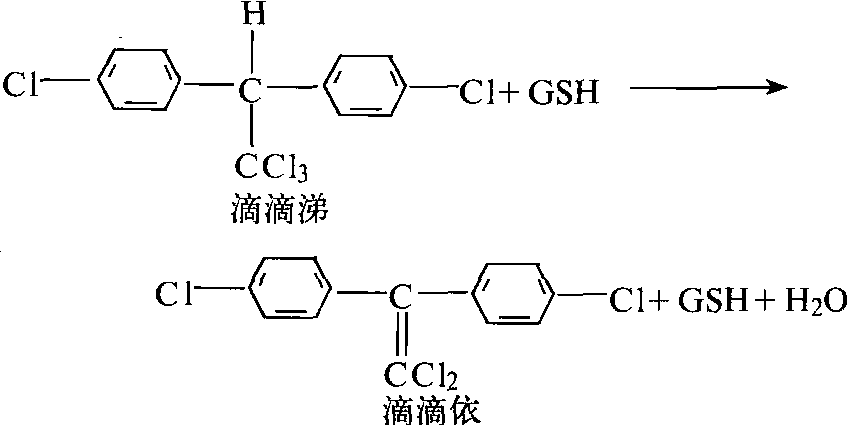
图1 滴滴涕—酶对滴滴涕的降解
涕抗药性家蝇和埃及伊蚊的抗药性机制,有些昆虫也可使滴滴涕代谢降解,但测不出滴滴依,DMC也不增效,可能为其他机制所致。谷胱甘肽S-转移酶 主要对有机磷化合物起降解作用的酶,是有机磷抗药性的重要机制。它可裂解有机磷杀虫剂的O-烷基部位(主要是对二甲基磷酸酯和二甲基硫磷酸酯),芳基和酸基等部位,而去烷基的产物比去芳基的多。此酶在谷胱甘肽参与下,可替换杀虫剂不稳定的部位,成为无毒代谢产物——有机磷底物与谷胱甘胱的复合体(图2)。
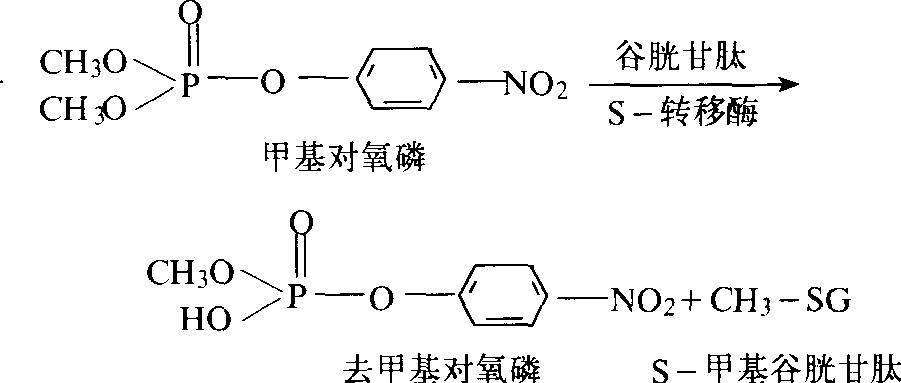
图2 谷胱甘肽转移酶对甲基对硫磷的降解
此酶对有机磷杀虫剂的催化作用,因杀虫剂底物和本身性质而不同。但谷胱甘肽S-转移酶的活性常不与抗药性程度平行,因此有时难以确定其在抗药性中的重要性。甲基碘和二乙基顺丁烯二酸(DEM)为谷胱甘肽S-转移酶的抑制剂,可用于侦测昆虫体内此酶的存在和确定其在抗药性中的作用。
磷酸酯酶 可降解对氧磷和氧二嗪磷为二乙基磷酸产物,降解马拉氧磷为二甲基磷酸的酶,对磷酸酯要比硫磷酸酯容易分解。某些有机磷抗药性家蝇品系的磷酸酯酶比感性品系有明显的高活力,这个差别可能为对磷酸酯产生抗药性的机制。磷酸酯酶的活力可被正-丙基对氧磷、对-氯汞基苯酸酯以及Hg++、Cu++所抑制。
羧酸酯酶 对降解带有羧酸酯基的二硫代磷酸酯等起重要作用的酶。例如,可将马拉硫磷代谢为α单酸和β单酸产物,二者均无毒性(图3)。但在有些情况
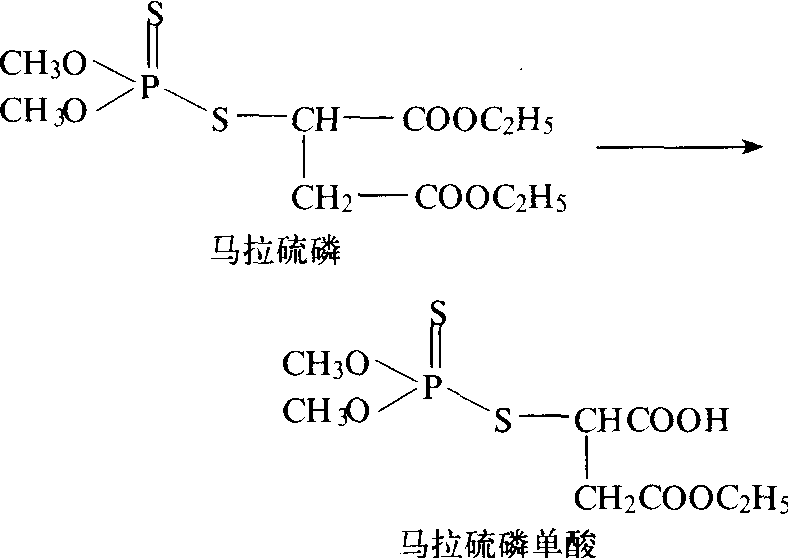
图3 羧酸酯酶将马拉硫磷代谢为单酸产物
下,此酶会伴随着一个酯酶变性和底物特异性变化的现象。有机磷抗药性和以α-乙酸萘酯、β-乙酸萘酯和丁酸甲酯以及其他酯类为底物的变性羧酸酯酶(或称非特异性酯酶)活力之间的关系为: 抗药性有时与非特异性酯酶低活力有关或有时是与其高活力有关。羧酸酯酶的抑制剂为磷酸三苯酯(TPP)、三-O-甲苯磷酸酯(TOTP)、SSS-三丁苯三硫赶磷酸酯(DEF)和对硫磷、地亚农等,它们都可以克服昆虫对马拉硫磷的抗药性。昆虫对马拉硫磷产生抗药性是由于体内羧酸酯酶活性增加。此酶也可使拟除虫菊酯的羧酸酯水解: 使除虫菊Ⅱ的甲氧羰基水解为低毒物质(图4);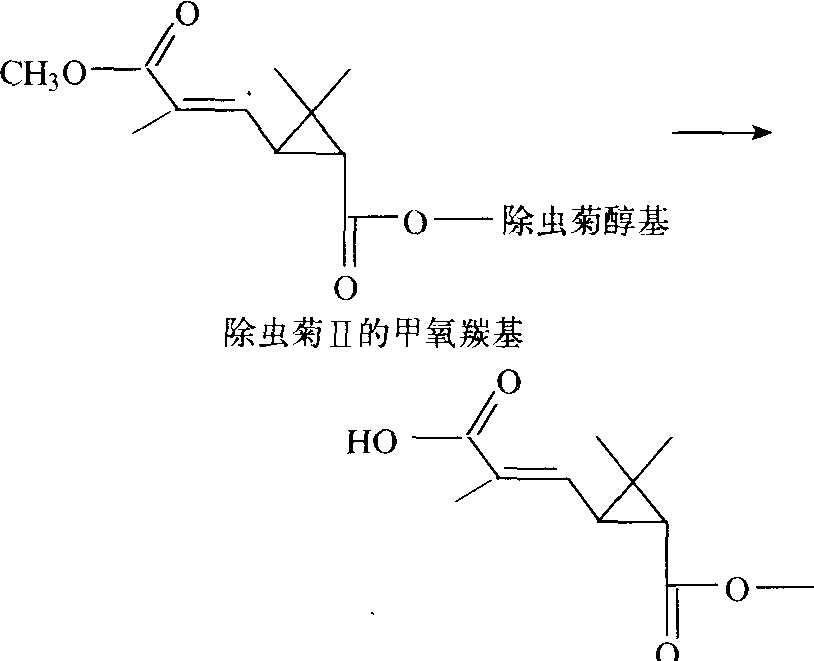
图4 羧酸酯酶对除虫菊Ⅱ的甲氧羰基水解作用
对带有反式侧链的环丙烷羧酸伯醇酯也易水解为低毒物质(图5)。由于此酶对(+)-反式异构体要比相应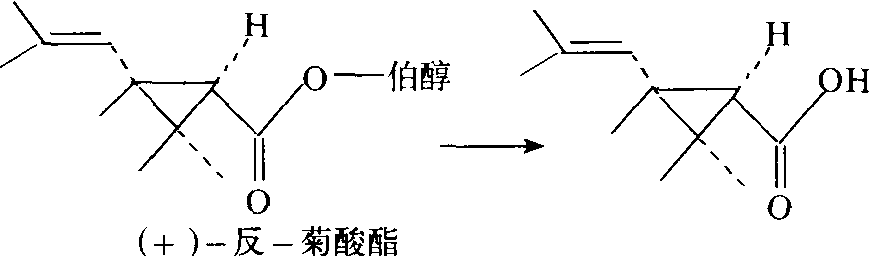
图5 羧酸酯酶对(+)-反-菊酸酯的水解作用
的(+)-顺式异构体水解快,所以昆虫对反式异构体易产生抗药性。微粒体多功能氧化酶 位于细胞中微粒体部分可催化多种类型氧化作用的酶。如硫醚和硫代磷酸酯的氧化作用、环氧化作用: 环羟基化作用、脂族侧链羟基化作用、N-甲基羟基化作用和O-和N-去烃基作用等,将有机杀虫剂或外来化合物代谢为水溶性的无毒化合物。但是在少数情况下,氧化后也增高毒性,如将艾氏剂环氧化为狄氏剂,硫磷酸酯氧化为磷酸酯。故称之为微粒体多功能氧化酶,但由于反应结果是酶的一个氧原子与底物结合,另一氧原子被还原作用生成水,也可称为单加氧化酶(mono-oxygenase)。它是有关抗药性的重要解毒酶,也是许多杀虫药剂之间产生交互抗药性的原因。可对多种杀虫剂代谢起作用。
滴滴涕代谢 在某些家蝇、德国蜚蠊、锥蝽、果蝇、库蚊和谷象等昆虫体内,滴滴涕的解毒途径主要是微粒体多功能氧化酶的羟基化作用,产物为三氯杀螨醇(图6)。昆虫对滴滴涕抗药性是与此酶的羟基化作用有关:
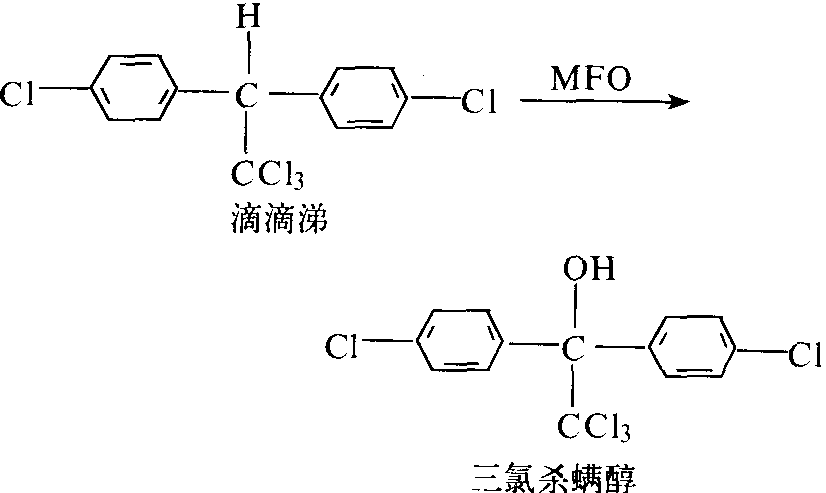
图6 MFO对滴滴涕的氧化解毒代谢
有机磷酸酯代谢 MFO酶可使硫磷酸酯(P=S)氧化或磷酸酯(P=O)、二硫代磷酸酯中侧键的硫醚氧化成亚砜和砜,其代谢产物的毒性更高,而表现为活化反应;对磷酸酯的烷基和芳基键氧化降解为低毒的或无毒的产物,表现为解毒反应。由于有机磷杀虫剂种类繁多,同时又为多功能氧化酶的作用而活化或降解,因此有机磷的抗药性机制是非常复杂的。其抗药性机制涉及的解毒酶系又有磷酸酯酶、羧酸酯酶、谷胱甘肽S-转移酶和MFO(图7)。由于这些解毒酶的
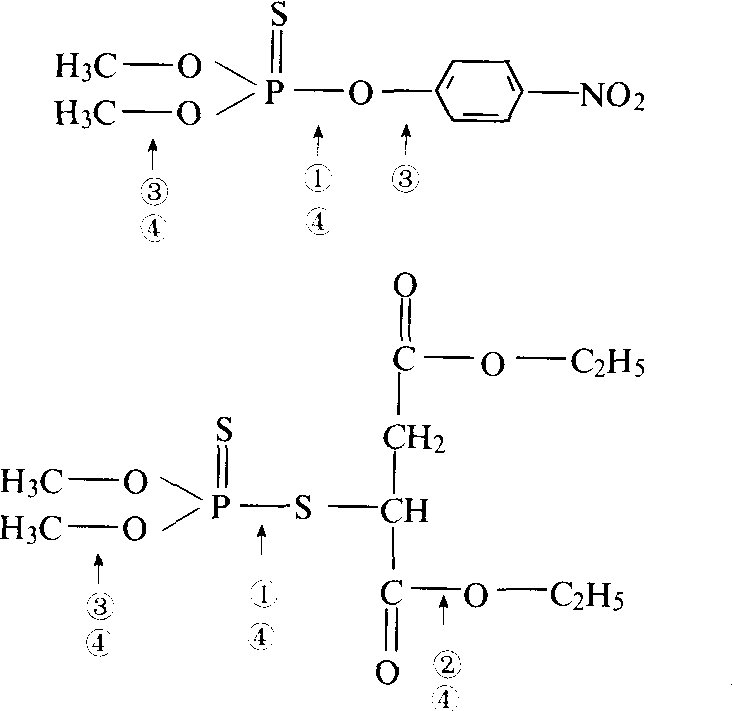
图7 有机磷化合物涉及的解毒酶系
❶磷酸酯酶;
❷羧酸酯酶;
❸谷胱甘肽S-转移酶;
❹MFO
氨基甲酸酯代谢 此类杀虫剂的主要解毒机制是多功能氧化酶,很多取代基都易被多功能氧化酶降解,甲基可被羟基化或脱甲基化。在家蝇和库蚊对残杀威的抗药性中,使残杀威氧化代谢为5-羟基残杀威、2-羟基苯基甲基氨基甲酸酯及N-羟基甲基残杀威。体内还可能由于第二步反应而产生其他产物。
除虫菊酯代谢 其降解主要是多功能氧化酶和羧酸酯酶活性的增高。如对丙烯菊酯,主要氧化代谢部位是酸部位上的异丁烯链上的两个甲基的氧化。先氧化为羟甲基衍生物,进一步为醛,再转变为羧酸,成为低毒或无毒的代谢产物。在醇部位的环烯酮的环侧链上也可以氧化降解代谢。对于二氯苯醚菊酯,氧化与水解作用几乎同样重要,可有四处氧化部位,两个是在环丙烯上的两个甲基,另两个是在间苯氧基苄醇上的氧化部位。一个异构体可以有20多个代谢产物,这些部位上的氧化代谢产物可以进一步水解。
不同昆虫种或品系,降解代谢会有所不同,同时随不同异构体也不同。一般反式异构体比顺式异构体不论在醇或酸部位上都易发生水解,也易氧化代谢。因此害虫对反式异构体较顺式易产生抗药性。昆虫微粒体多功能氧化酶对杀虫剂的解毒代谢在抗药性中的作用,可以用增效剂抑制虫体内氧化酶而显示对杀虫剂的增效作用来判断。增效剂有增效醚(piperonylbutoxide)、芝麻素(sesamex)和增效磷(SV1)等。
保幼激素类化合物代谢 很多种对保幼激素及其类似物也产生抗药性的昆虫,是由于多功能氧化酶对底物的非特异性解毒反应,或脱甲基和羟基化作用,而引起广泛交互抗药性。
靶标部位敏感性降低 包括对有机磷和氨基甲酸酯抗药性有关的乙酰胆碱酯酶(AchE)的改变、滴滴涕和拟除虫菊酯受体蛋白靶标基因的改变和对环戊二烯杀虫剂受体蛋白靶标的改变三类。
有机磷酸酯和氨基甲酸酯主要通过抑制靶标酶而发挥它们的毒效作用。AchE的改变造成对这两类杀虫剂敏感性的变化,而不受其抑制或毒害。感药性品系昆虫的等位基因形成的酶极易被抑制,而抗药性品系的等位基因形成的酶对有机磷酸酯不敏感。这表明在两品系间的酶受抑制的差别是由于酶本身的不同,酶的改变不一定与减少执行正常生理功能有关。大部分正常昆虫中都存在着一定水平的解毒能力,而抗药性品系降低了AchE抑制速率,可使存活时间延长,其解毒速率就足以使药剂清除。
环戊二烯杀虫剂受体蛋白靶标的改变是由于昆虫体内存有对环戊二烯杀虫剂敏感的受体蛋白。对环戊二烯类杀虫剂有抗药性的蜚蠊,对植物衍生物的神经毒素(木防己苦毒素)有交互抗药性,此种蜚蠊的神经成分中,含的H3标志的α-二氢木防己苦毒素,明显少于感性蜚蠊。抗药性家蝇可降低受体蛋白的结合,主要是结合部位的数量较少。
滴滴涕和拟除虫菊酯受体蛋白的靶标基因变化,一是由于抗药性昆虫的靶标部位数量少,而不是亲合力质上的不同。数量上减少,就使药剂作用于靶标部位上的剂量少,延长致死。二是由于Kdr(抗击倒)基因存在,这是一种非代谢的抗药性机制。昆虫神经系统对滴滴涕和除虫菊酯敏感性降低,与滴滴涕酶、多功能氧化酶和酯酶无关,增效剂的使用也不显出增效。此基因不但有抗击倒作用,并且死亡率也减少。滴滴涕和除虫菊酯虽然化学结构不同,但都影响轴突的传导,神经轴突是由一个半渗透的磷脂双分子膜包围的双层磷脂结构。轴突在静止时,膜电位为-60~-70毫伏,处于极化状态。当轴突传导一个信息时,而使轴突膜电位出现上升、下降、正相和负后电位四个阶段而组成一个作用电位。在用二氯苯醚菊酯处理家蝇幼虫感觉神经时,会引起自发放电频率增加,而处理抗药性品系家蝇时,则不显示有变化。这说明抗药性系的神经膜较不敏感。Kdr基因早期由意大利一种家蝇抗药性品系中发现,它对滴滴涕和除虫菊酯有交互抗药性,但是滴滴涕酶不多,并不被滴滴涕所击倒。Kdr基因是一种隐性基因。根据联合国世界卫生组织(WHO)建议,可用增效剂测试方法来判断Kdr因子抗药性。
其他抗药性机制 杀虫剂进入虫体内,可部分贮藏在某些组织中,特别是脂肪体内。由于抗药性昆虫脂肪组织中药剂含量大于敏感昆虫,以使杀虫剂到达抗药性昆虫体内的靶标部位的需要量加大。此外,抗药性昆虫还会使较多杀虫剂直接排出体外,而减少中毒,增强对杀虫剂的抵抗力。
- 薅锄是什么意思
- 薅锄子是什么意思
- 薅锄锄是什么意思
- 薅除是什么意思
- 薅馬是什么意思
- 薅马是什么意思
- 薅麦是什么意思
- 薅麦子是什么意思
- 薅鼓是什么意思
- 薅(田)是什么意思
- 薆是什么意思
- 薆然是什么意思
- 薆薆是什么意思
- 薆薱是什么意思
- 薇是什么意思
- 薇䕔是什么意思
- 薇云室是什么意思
- 薇云室诗稿是什么意思
- 薇卿是什么意思
- 薇君是什么意思
- 薇堂是什么意思
- 薇客是什么意思
- 薇山是什么意思
- 薇山老人是什么意思
- 薇拉·米哈伊洛夫娜·英蓓尔是什么意思
- 薇省同声集是什么意思
- 薇省词钞是什么意思
- 薇研是什么意思
- 薇老首阳是什么意思
- 薇老首阳,芝深商谷,时遥雾拥云平。是什么意思
- 薇芜是什么意思
- 薇草是什么意思
- 薇药獐毛是什么意思
- 薇菜是什么意思
- 薇蕨集是什么意思
- 薇蕪是什么意思
- 薇薇夫人是什么意思
- 薇薇的周记是什么意思
- 薇藿是什么意思
- 薇衔是什么意思
- 薇衔茎叶是什么意思
- 薇西小舫是什么意思
- 薇轩是什么意思
- 薇采是什么意思
- 薇銜是什么意思
- 薇香是什么意思
- 薇香与果牟是什么意思
- 薇香集是什么意思
- 薈是什么意思
- 薉是什么意思
- 薉孽是什么意思
- 薊是什么意思
- 薋是什么意思
- 薋丞之印封泥是什么意思
- 薋葹是什么意思
- 薌是什么意思
- 薌合是什么意思
- 薌萁是什么意思
- 薍是什么意思
- 薎是什么意思