遗传密码genetic code
信使核糖核酸(mRNA)中决定蛋白质的氨基酸排列的核苷酸三联体顺序。脱氧核糖核酸(DNA)分子上的遗传信息由腺嘌呤(A)、胸腺嘧啶(T)、鸟嘌呤(G)、胞嘧啶(C)四种核苷酸组成。mRNA的遗传信息从DNA双链中一条单链(编码链)转录而来,每三个核苷酸构成一个密码子。转运核糖核酸 (tRNA) 分子中有三个与mRNA的三联体密码互补配对的核苷酸称反密码子。每种tRNA具有特定的反密码子,因此蛋白质合成时每种tRNA携带一种特定的氨基酸到mRNA链上“对号入座”,即按mRNA上核苷酸顺序决定氨基酸顺序,也就是决定蛋白质种类,故DNA是通过mRNA指导蛋白质合成的。
简史 关于三联体密码的发现有以下推理及主要试验。一个核苷酸决定一个氨基酸的推理显然不能成立。两个核苷酸的可能组合数是42=16种,而已知的氨基酸种类至少有20种之多,故两个核苷酸组成的密码子也不敷应用。1961年英国学者克里克(F.H.C.Crick)等用吖啶类染料原黄素诱发大肠杆菌噬菌体T4的野生型,获得突变型rⅡ。用原黄素再处理这种突变型可使之成为具有野生型表型的回复体。将回复体再与原来的野生型杂交,子代中出现一些rⅡ突变型。由于原黄素诱发的是移码突变,即由于个别核苷酸的增加(用+表示)或缺失(用-表示)引起解读密码的错误而导致的突变,因此可把rⅡ突变型写作+(或-),而把由rⅡ突变回复正常的基因(或核苷酸)写作-(或+)。即由于一个核苷酸的增加或缺失使其位置之后的密码意义发生错误而成为突变型。当其相邻或靠近的另一核苷酸缺失或增加,则可校正后面核苷酸使解读回复正常,或解释为另一座位的突变掩盖了原来座位的突变(此即谓“基因内抑制”,见抑制基因),从而使表型回复为野生型。通过基因重组或突变可得到各种突变型,如++、--、---等(表1)。从表1说明除+-(或-+)以外的各组合中只有---或+++或------等组合是野生型,这一实验结果证明密码子由三个核苷酸组成。
表 1 T4噬菌体的rⅡ+、-突变型表型
| 基 因 型 | 表型 | 说 明 |
| +或- | 突变型 | 单个位点发生增加或缺失 突变 |
| ++或-- | 突变型 | 两个位点同时发生增加或 缺失突变 |
| ----或----- | 突变型 | 四个或五个位点同时发生 缺失突变 |
| +-或-+ | 野生型 | 增加和缺失(缺失和增加) 两个突变位点组合在一起 |
| ---或+++ | 野生型 | 三个位点缺失或三个位点 增加的突变 |
| ------ | 野生型 | 三的倍数的位点缺失(或增 加)的突变 |
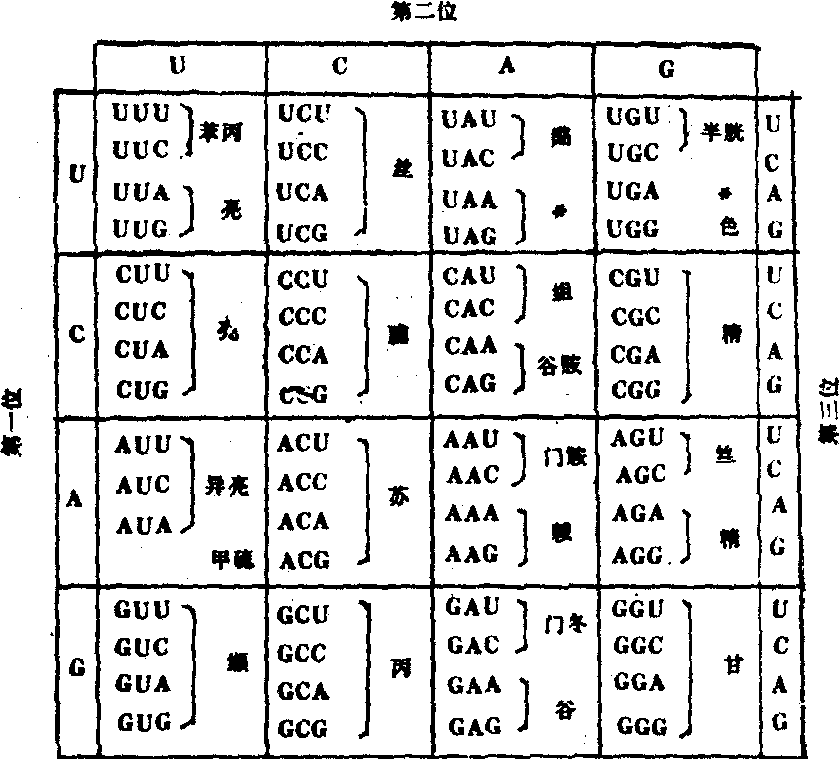
1961年美国学者尼伦伯格 (M. W. Nirenberg)等以人工合成的mRNA作模板,进行蛋白质的离体合成,随后再测定产生的多肽的氨基酸顺序。发现多聚U作为信使则合成多聚苯丙氨酸,多聚A作为信使则合成多聚赖氨酸; 多聚C产生多聚脯氨酸; 多聚G产生多聚甘氨酸等。说明苯丙氨酸、赖氨酸、脯氨酸、甘氨酸等的密码子分别是UUU、AAA、CCC、GGG等。美国学者柯拉纳(H.G.Khorana)用共聚的多核苷酸UGUGUGUG……作为信使则合成缬氨酸与半胱氨酸多聚体。以后确证缬氨酸的密码子是GUG,半胱氨酸的密码子是UGU。1964年开始,尼伦伯格等进而用64种人工合成的三核苷酸和氨基酸-tRNA(即与tRNA结合的活化氨基酸)、核糖体三者进行结合试验,证明各种三核苷酸具备信使作用。例如三核苷酸是UUU时只与苯丙氨酰-tRNA结合;AAA只与赖氨酰-tRNA结合,而二核苷酸无此作用。用这一方法最终揭示了64种密码子中61种密码子与20种氨基酸之间的对应关系,于1966年完成遗传密码表(表2)。5′端核苷酸、中间核苷酸,3′端核苷酸(或称第一碱基、第二碱基、第三碱基)依次组成一个密码子,此称三联体密码。表2中所列64个密码子中有61个编码18种氨基酸和2种酰胺。其他如胱氨酸、羟脯氨酸、羟赖氨酸等是在肽链合成后再行加工而成。还有3个不编码任何氨基酸只作为终止信号的密码子。密码子AUG既编码甲硫氨酸又有合成起始点的作用。
表2 遗传密码表
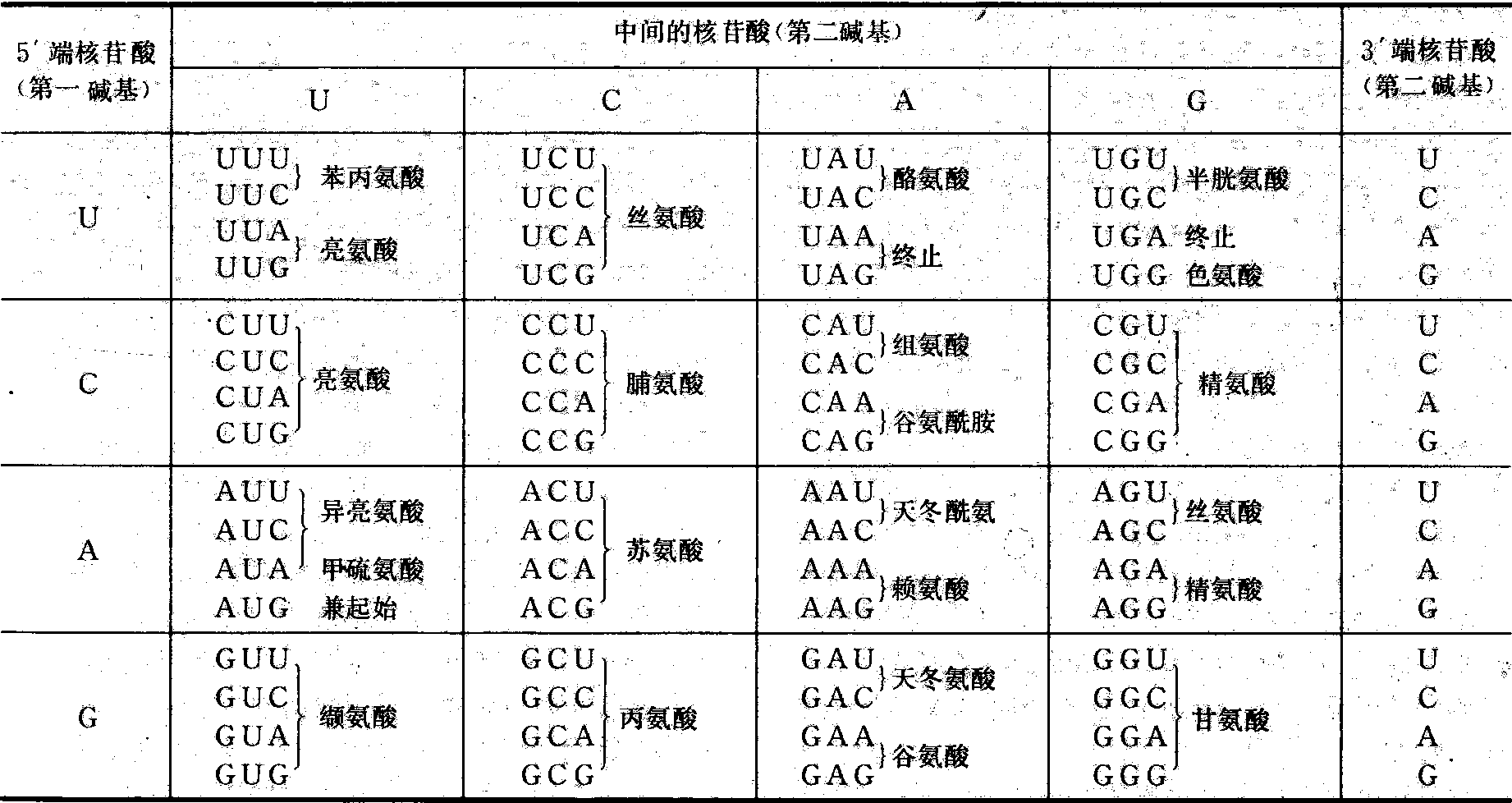
特性 ❶密码子相互之间没有 “逗号”,没有 “间隙”或“闲置”的核苷酸,即除了终止密码子外,各密码子之间没有任何不编码的核苷酸。从起始密码子(AUG)开始,沿mRNA多核苷酸链由5′到3′的方向进行转译,直至遇到终止信号UAA、UAG或UGA时便停止转译。
❷密码子之间没有重叠性,即mRNA多核苷酸链上的任何两个相邻的密码子不会共有同一个核苷酸。如多核苷酸链是ACUGAUGUG,则这三个密码子分别编码苏氨酸、天冬氨酸、缬氨酸,不可能交叉重叠组合成三个以上的密码子,编码三个以上的氨基酸。
❸简并性。大多数氨基酸可由2个以上密码子识别。例如赖氨酸有两个密码子、丙氨酸有四个密码子; 精氨酸、亮氨酸分别有六个密码子。识别同一氨基酸的密码子称同义密码,也称简密码。克里克认为三联体密码中第一、第二核苷酸的顺序是严格的,第三个可以变动为任何核苷酸,但仍决定同一氨基酸。简并现象对生物遗传性的稳定具有重要意义。密码子的简并性也反映在反密码子中,即反映在tRNA分子中,某些氨基酸能被一个以上的tRNA分子携带到mRNA特定位置上。识别同一氨基酸的不同tRNA称同功tRNA。
应用 查明决定各种蛋白质的遗传密码,就可人工合成有用的蛋白质分子直接应用于医疗和日常生活;并进而利用遗传工程手段在农业动植物生产与创新、医药卫生、工业生产、环境保护等方面造福人类。现已证明,从病毒到人类的遗传密码是通用的,故密码研究可作为进一步探索生命奥秘的手段。
遗传密码
DNA链上每三个碱基(也即三个核苷酸)顺序,作为决定一个氨基酸的密码,叫遗传密码或称三联密码。人类的一切遗传性状,都是在发育过程中,经过一系列的新陈代谢而形成,是蛋白质活动的一种体现。也就是说,所有的代谢过程都是在酶蛋白质的控制下进行,并形成一定的遗传性状。已知遗传密码存在于细胞核的DNA中,而蛋白质的合成则是在细胞质中进行的,遗传密码是怎样指令合成蛋白质而表现出遗传性状的呢? DNA不直接决定蛋白质的合成,它是将DNA所携带的遗传信息先传递给mRNA(信使RNA),此过程叫转录。转录时,DNA的双链暂时脱开,其中的一条链起模板作用,按碱基互补进行碱基配对。mRNA在细胞核中合成后就转录上DNA遗传信息,通过核膜的核孔转入细胞质中附着于核蛋白体上,以其密码控制蛋白质的合成,此过程叫翻译。也就是说,密码中应用的是mRNA的核苷酸顺序,控制着蛋白质的合成。DNA含有四种碱基,在三个相邻位置上随机排列组合,可能的组合数有43=64种密码。一个密码可决定一种氨基酸,而氨基酸只有20种,所以往往有几个不同的密码决定同一种氨基酸。这些不同密码,可以称为同义密码。同义密码愈多,这种氨基酸的稳定性就愈大。对于只有一个密码的色氨酸来说,如三联密码中有一个碱基发生改变,则整个遗传信息就会改变,而使生物出现不同性状。而有六个密码的精氨酸就比较稳定。在64种密码中AUG是起始符号,三种密码UAA、UAG、UGA没有相应的氨基酸是终止符号。
遗传密码genetic code
按核苷酸三联体顺序排列的64种密码子所决定的遗传信息。其中少数是无义密码子,部分是同义密码子。在指导蛋白质合成中,每个密码子可决定相对应氨基酸在多肽链上的位置,总的氨基酸序列决定于由遗传信息规定的遗传密码排列顺序。在以脱氧核糖核酸(DNA)作为遗传物质的生物,DNA双链分子中只有一条为有义链,可将遗传信息由DNA转录到信使核糖核酸(mRNA)上。通常遗传密码就用RNA,而不是用DNA中的核苷酸碱基排列顺序表示。具有以下性质:
❶统一性。原核生物和真核生物基本相同。
❷三联体。每三个相连续的核苷酸决定一种氨基酸。
❸不重叠。任何两个相邻密码子,没有共同的核苷酸。
❹无逗号。所有三联体密码都是连续的,中间没有不作为密码的核苷酸。
❺兼并性。多数氨基酸可由几个密码子代表。
遗传密码
决定蛋白质各个氨基酸构成的核苷酸三联体顺序(密码子)。脱氧核糖核酸(DNA)分子上的遗传信息由腺嘌呤(A)、胸腺嘧啶(T)、鸟嘌呤(G)和胞嘧啶(C)4种核苷酸组成。mRNA的遗传信息从DNA双链中一条单链(编码链)转录而来,每3个核苷酸构成一个密码子。转运核糖核酸(tRNA)分子中有3个与mRNA的三联体密码互补配对的核苷酸称反密码子。20世纪60年代末期,已将由四种碱基(U.C.A.G)每3个一组而构成的64个密码子所决定的氨基酸及终止密码全部破译出来。
遗传密码yichuan mima
又称密码子或三联体密码,指DNA或其转录产物mRNA的碱基序列与蛋白质的氨基酸序列的相互对应关系。DNA转录成mR-NA,mRNA又决定所形成多肽链中的氨基酸序列。
mRNA中的主要核苷酸只有4种,而蛋白质中的氨基酸有20种。4种核苷酸怎样排列组合才足以代表20种氨基酸呢?用数学方法推算,如果每个核苷酸代表1种氨基酸,则只能代表4种氨基酸,这显然是不够的。如果两个核苷酸代表1种氨基酸,则只可能有42=16个组合,也不够为20种氨基酸编码。如果3个核苷酸为1组,可以产生43=64个不同的组合。早期的遗传学实验也证明遗传密码是由3个核苷酸组成的。但是,不同氨基酸各有什么样的遗传密码,以及它们又是如何鉴定,当时还不清楚。
1961 年尼伦伯格(Marshall Nirenberg)和马蒂(Heinrich Mattaei)所做的实验是一个突破。他们用人工合成的多聚尿苷酸与大肠杆菌提取物、GTP和20种氨基酸的混合物分别在20支试管中共同温育。每支试管中有一种不同的氨基酸是经放射性同位素标记过的。由于多聚尿苷酸可以看作是含有许多UUU三联体密码的人造信使mRNA,只有UUU编码的氨基酸才能合成放射性多肽。实验结果是:只有含放射性苯丙氨酸的那支试管中产生了放射性多肽。就是说,放射性多肽是多苯丙氨酸,是一种只含有苯丙氨酸的多肽。因此,苯丙氨酸的遗传密码是UUU。利用相同的方法,很快发现脯氨酸的遗传密码为CCC,赖氨酸的遗传密码是AAA。1964年,尼伦伯格等又获得另一突破性进展。他们发现:如存在相应的合成mRNA,离体的大肠杆菌核糖体将与特定的氨酰-tRNA(tRNA与其专一氨基酸的结合产物)结合。例如,当核糖体与多聚尿苷酸和苯丙酰-tRNAphe一起温育时,它与两者都结合;而当核糖体与多聚尿苷酸和其他氨酰-tRNA一起温育时,它就不能和后者结合。因为这些tRNA的反密码子不能识别多聚尿苷酸中的UUU三联体密码。结合的tRNA复合物可被硝基纤维素滤膜吸附,未结合的tRNA则不能被吸附,所以二者能区分开。这样,用各种已知序列的简单三联体作为mRNA,就有可能确定与不同氨酰-tRNA专一结合的那些密码子中的序列。果然,利用人工合成的密码子及其他的手段,很快阐明了20种氨基酸的全部遗传密码并用许多不同的方法检查了这些密码子的正确性。密码子的阐明被看作是60年代最伟大的科学发现。表中列出了氨基酸的遗传密码字典。
氨基酸的三联体遗传密码字典(用mRNA中的碱基序列表示)

*起始密码子或终止密码子;AUG为起始密码子,UAA,UAG和UGA为终止密码子。上表的读法是从左到右,从5'到3'。
遗传密码有以下特点:(1)无需任何标点或信号指示mRNA上一组密码子的终止和其次一组密码子的开始。因此,“可译框架”必须正确处在mRNA分子的开始处,然后从一组三联体移向另一组三联体依次转译。如果起初的可译框架缺少1或2个核苷酸或者核糖体偶然漏译了1个核苷酸,所有以后的密码子就会发生错误,导致产生氨基酸序列已经改变的、无生物活性的蛋白质。(2)有3组密码子(UAG、UAA和UGA)不为任何氨基酸编码,它们是多肽链合成终止的信号,也叫做终止密码子。AUG既是多肽链起始合成的信号(起始密码子),也为多肽链内部的甲硫氨酸编码。(3)密码子是通用的,即适用于迄今检查过的多种生物,包括人类、大肠杆菌、烟草、两栖类和病毒。但近年来发现线粒体的遗传密码有所不同。如酵母的线粒体使用AUA为甲硫氨酸而不为异亮氨酸编码;使用通常的终止密码子UGA为色氨酸编码等。不同物种线粒体的密码子也有所不同。(4)密码子有“简并性”,即一种氨基酸可能有数组密码子。除甲硫氨酸和色氨酸只有一组密码子外,多数氨基酸对应的密码子多于一组(2~6组)。简并性并不意味着密码不完善,因为每组密码子只对一种氨基酸专一。当一种氨基酸有多组密码子时,密码子之间的差异多在第3个碱基(3′端)。如丙氨酸的4组密码子为GCU、GCC、GCA和GCG,其头两个碱基GC是相同的。其他氨基酸的密码子也有类似的规律。这说明密码子的头两个核苷酸是其专一性的决定因素;第3个位置,即密码子3′端核苷酸专一性较差。DNA、mRNA与蛋白质的对应关系可简示如图。
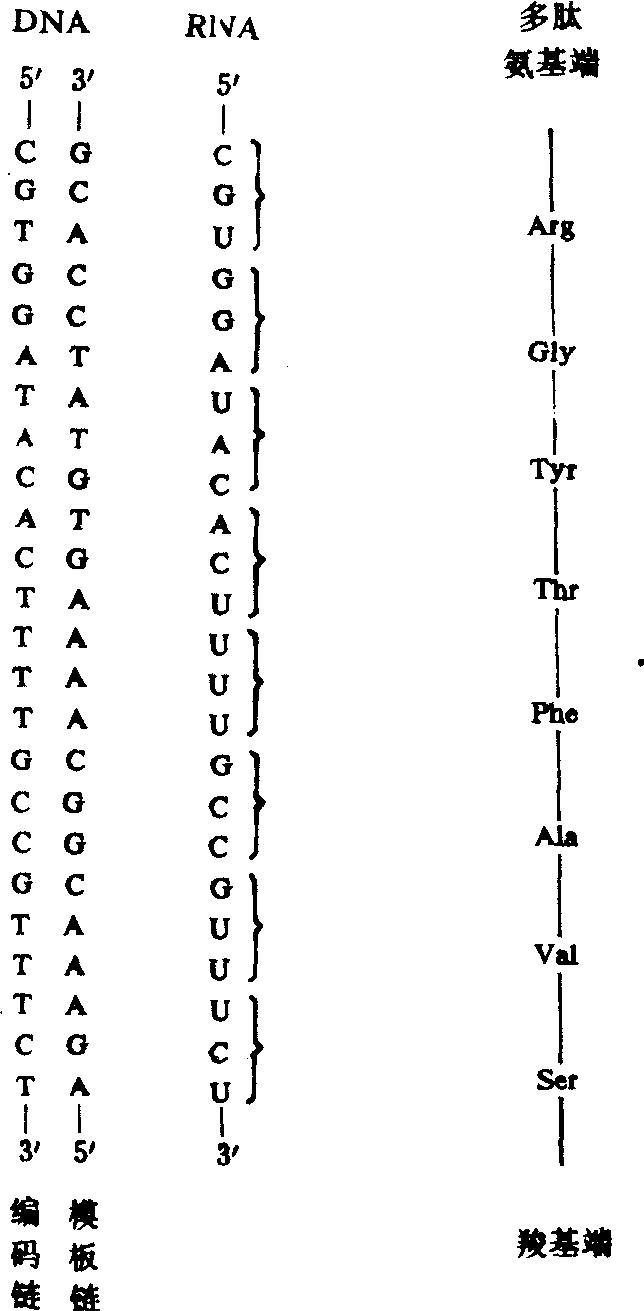
图585 DNA、mRNA与蛋白质的对应关系
尼伦伯格因发现遗传密码与合成遗传密码的柯拉纳(H.G.Khorana)同获1968年度的生理学或医学诺贝尔奖。由于自1979年以来,已先后在人、牛、酵母和链孢霉等的线粒体及枝原体、腺病毒和包括草履虫等3种原生动物中发现了变异密码子的存在,变异密码子的数目多达10个以上,有人认为:生物界可能存在另一张密码表。遗传密码genetic code
又叫遗传信息,系子代从亲代获得控制性状和发育的遗传讯号。遗传密码既存在于细胞核内又存在于细胞质内,但以细胞核内为主;既存在于脱氧核糖核酸中,又存在于核糖核酸中,但以脱氧核糖核酸为主;遗传密码的本质是3个顺序排列的碱基;机体的遗传密码数目之大难以计算。如:染色体是遗传的物质基础,而发生遗传效应的是染色体的DNA分子,每个DNA分子上又有无数个基因,而基因是由按一定顺序排列的核苷酸组成的,但DNA分子上只有4种核苷酸,关键是这4种核苷酸却组成极为复杂的排列顺序,因此存在着极其众多具有各种功能的基因。以基因控制蛋白质合成而言:它是由转运RNA与信息RNA完成的,转运RNA一端有3个碱基(为1个遗传密码)与信息RNA配对,另一端携带氨基酸,每一种运转RNA只能运转一种氨基酸,这是一个密码(3个碱基)决定的,20种氨基酸的转运密码已完全清楚。经研究证明遗传密码具有不重叠性,任何两个相邻的密码子之间没有共用的核苷酸;所有密码都是由3个按一定顺序排列的核苷酸组成的;多数氨基酸可各由几个密码代表;各种生物的遗传密码基本相同,因此各种生物的蛋白质组成、性状、生理功能等均基本相同,细胞的基本生命特征亦相同。
遗传密码
决定蛋白质中氨基酸顺序的核苷酸顺序。由3个连续的核苷酸组成的密码子构成。用mRNA中的核苷酸顺序表示。
遗传密码genetic code
是DNA分子中的四种含氮碱基,三个三个地排列组合,组成64种三联体,即为遗传密码。每一个密码与一种氨基酸相对应。
遗传密码
生命蛋白质合成的密码,是遗传信息的单位。由构成核酸的四种不同核苷酸的不同组合所代表。每一密码由核酸分子中三个相连核苷酸所组成,决定一个氨基酸。此外,还有代表遗传信息转译起点和终点的密码。
遗传密码
密码是一种信号或信息。遗传密码就是组成遗传信息的基本单位。现在已经知道,组织蛋白质的氨基酸通常不超过20种,而组成DNA或RNA的核苷酸只有4种。在蛋白质的合成过程中,DNA(或RNA)与氨基酸之间必须有一种确定的对应关系,即核苷酸的排列顺序体现为氨基酸的排列顺序。原来核苷酸是以三个作为一组,编码一个氨基酸,而成为遗传密码的。这样,用4种核苷酸(碱基)就可以组成43=64个遗传密码。因为只有20种氨基酸,所以有些氨基酸的遗传密码不止一个,有的遗传密码是蛋白质合成的终止信号。
遗传密码
在信使核糖核酸内,每三个相互邻近的核苷酸,其特定排列顺序,在蛋白质生物合成中可作为某种氨基酸的或蛋白质生物合成终止信号者,统称为遗传密码。有时其中的某一单个密码字,亦被称为密码子。
信使核糖核酸所含的腺嘌呤(A)、尿嘧啶 (U)、鸟嘌呤(G)、胞嘧啶 (C)四种核苷酸,三个一组相联,可排列成64种不同的三联体,所以遗传密码字,共有64个也只有64个。
位于信使核糖核酸起动部位的AUG为肽链合成的起动信号: 在真核生物中代表蛋氨酸,在原核生物中代表甲酰蛋氨酸。在原核生物中,GUG有时也可作为起动信号,此时代表甲酰蛋氨酸,而非代表缬氨酸。
密码与各种氨基酸的对应关系
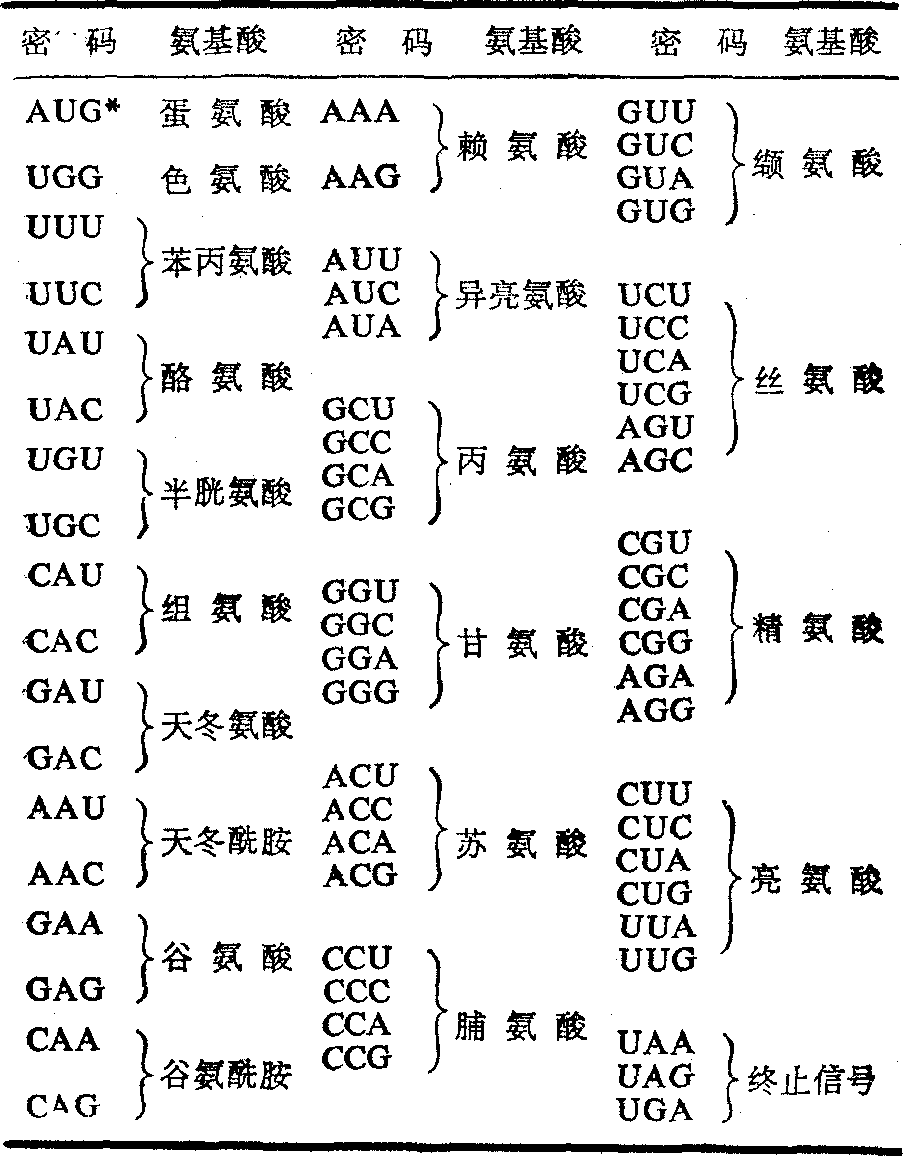
64个密码子中,61个密码子分别代表各种氨基酸,另外三个密码子(UAA,UAG,UGA)为肽链合成的终止信号,不代表任何氨基酸,故亦称为无意义密码子。每一种氨基酸少的只有一个密码子,多的可有六个,但以两个和四个居多(表)。
信使核糖核酸的这种密码子按A可与U配对,G可与C配对的原则,可被作为特定氨基酸载体的各种转运核糖核酸的相应反密码所辨认。反密码亦由三个相互邻近的核苷酸构成,因其与信使核糖核酸相应密码子的核苷酸相互补,故可藉以上的配对原则而辨认之。
上述遗传密码在生物界是通用的。有时遗传密码亦被用来泛指脱氧核糖核酸中,由相应三联体构成的遗传信息。
遗传密码
遗传密码是规定蛋白质上氨基酸序列的核酸碱基序列。DNA的遗传信息转录至RNA后再翻译成蛋白质。构成核酸的碱基有四种: DNA有腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、和胸腺嘧啶(T)或者RNA有A、G、C和U(尿嘧啶)。构成蛋白质的氨基酸有20种。四个字母的核酸语言如何翻译为20个字母的蛋白质语言就是遗传密码的内容。
单独一个碱基只能为4个氨基酸编码,两个碱基在一起也只能编码42个氨基酸,三个碱基一组可有43个编码,对于20个氨基酸已经足够。若遗传密码由碱基三联体组成,还要决定它们是重叠的或非重叠的。如AUGG-UCAAU的顺序,在密码非重叠的情况下可得AUG、G-UC、AAU三个三联体密码子; 当密码有一个碱基重叠就得AUG、GGU、UCA和AAU四个密码子; 当有两个碱基重叠,就有更多的密码子: AUG、UGG、GGU、GUC、UCA、CAA和AAU。可见同等长度的DNA,采用重叠方式就增大编码能力,但也有更多限制。如果有一个碱基重叠,则对某一给定氨基酸则只有16种氨基酸可跟随其后,因为密码子的最后一位碱基决定了下一密码子的第一位碱基。同理,如有两个碱基重叠,则只有4种氨基酸可出现在某一指定氨基酸之后,因为下一密码子的第一位和第二位碱基已由前一密码子决定了。根据对大量蛋白质序列的研究分析,并未见有如上述的氨基酸序列限制,所以密码子不是重叠的。再者,在重叠方式的密码编码法,一个碱基发生突变,相应的蛋白质会出现相连的两个或三个氨基酸残基的改变; 但对很多变异蛋白质(如各种变异血红蛋白)的分析,所观察到的只是单个氨基酸残基的改变。
非重叠性的三联体密码子在1961年经遗传学方法得到证明。吖啶可嵌入DNA双螺旋,引起碱基的插入[以(+)表示]或缺失[以(-)表示]而出现突变。噬菌体T4的rⅡ顺反子(一个顺反子是编码一条多肽链的一段DNA)经吖啶处理后,从其在大肠杆菌培养基噬斑的表现可观察到插入或缺失的影响。实验结果是插入一个碱基(+)或缺失一个碱基(-)时,T4显出突变;插入两个(+)或缺失两个(-)也显出突变,但插入三个(+)或缺失三个(-)却显出野生株性状; 一个(+)加一个(-)也显野生株性状。这正好说明密码子是非重叠的三联体。设野生株的密码子顺序是:
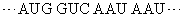
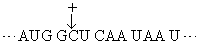
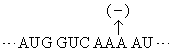
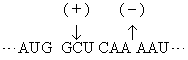
既然密码子是三联体,根据某一mRNA的碱基序列及其所指导合成的蛋白质序列就可破译遗传密码。只是当初尚不知任一mRNA的一级结构,破译只得根据其他实验。最先在无细胞的蛋白质合成体系中加入多聚尿苷酸作为信息,却合成出多聚苯丙氨酸,因为这种同聚物是UUU三联体的重复,所以可以决定苯丙氨酸的密码子是UUU。随后用多聚胞苷酸和多聚腺苷酸作实验,分别获得多聚脯氨酸和多聚赖氨酸,因此CCC和AAA分别是脯氨酸和赖氨酸的密码子。进而采用人工合成的随机共聚体作实验。如多聚AC可含AAA、AAC、CAA、ACA、ACC、CCA、CAC、ACC和CCC等9种三联体。这不同三联体各占比例随共聚体中A与C的比例而定。以A:C为3:1为例,上述各密码子依次占·42 (·75× ·75×·75)、·14 (·75×·75×·25)、·14、·14、·05 (·75×·25×·25)、·05、·05和·01 (·25×·25×·25)。以这些随机合成多聚AC作模板来指导多肽合成,观察到有赖氨酸、苏氨酸、谷氨酰胺、天冬酰胺、脯氨酸和组氨酸的参入。与已知以AAA为密码子的赖氨酸相比较,苏氨酸、谷氨酰胺、天冬酰胺各约为赖氨酸的1/3,可推断这三种氨基酸的密码子含2A1C,而脯氨酸和组氨酸的参入量只及赖氨酸的1/10,可推测其密码子含1A2C。不过依然不能得出密码子中碱基顺序。
用具有重复碱基顺序的合成多聚核苷酸可以解决密码子顺序。如用含重复二核苷酸AC的ACACACACAC…多聚核苷酸作模板,由于阅读起点不同而有两种信息,即ACA CAC ACA…或者是A CAC ACA CAC……,合成多肽是[苏-组]n和[组-苏]n。已知组氨酸密码子含1A-2C,可知CAC应是组氨酸密码子,ACA是苏氨酸密码子。还可用具有重复三核苷酸或四核苷酸的非随机共聚体去破译密码子。重复的二、三或四核苷酸顺序多核苷酸分别最多指导两种、三种或四种氨基酸参入蛋白质。对大多数核苷酸,每种可能的阅读方式都可有翻译产品。例如多聚CCA指导多聚脯氨酸、多聚组氨酸和多聚苏氨酸的合成,这一多聚核苷酸起了多聚CCA、多聚CAC和多聚ACC的作用。多聚GUA和多聚GAU只被译出可能的三种多肽中的两种,而多聚GUAA和多聚AUAG则只能译出二肽和三肽。这都符合UAA、UAG和UGA是肽链终止信号的发现。
人工合成多核苷酸促进氨基酰-tRNA结合到核糖体上,并有快速简便鉴定结合到核糖体上的氨基酰tRNA方法。核糖体吸附在硝酸纤维素微孔滤膜上,而放射性标记氨基酰-tRNA则可冲洗透过滤膜,除非和核糖体结合了。多聚尿苷酸引起苯丙酰-tRNA与核糖体结合而吸附在微孔滤膜上,而其他三种同聚核苷酸则无此作用。用共聚多核苷酸作这种氨基酰-tRNA结合测定,证明其所能促进结合到核糖体上的氨基酰-tRNA种类正是其指导多肽合成时所能使之参入的同样几种氨基酸,说明氨基酰-tRNA-共聚核苷酸-核糖体复合体的形成完全遵守蛋白质合成时的遗传密码。还发现短至含三个核苷酸的UpUpU就足以促进苯丙酰-tRNA与核糖体结合,但UpU则无效,也说明遗传密码是三联体。随后合成各种已知顺序三苷核酸作实验,可得出各种氨基酸密码子。
上表为遗传密码字典。64个密码子中有三个是无意义密码子,它们不代表任何氨基酸,却是蛋白质合成终止信号(参见“密码标点法”)。其余61个密码子分别代表20种氨基酸,不过不是平均分布的:有的氨基酸只有一个密码子,如甲硫氨酸和色氨酸,另一些又多达六个密码子,如精氨酸、丝氨酸和亮氨酸,其余的由二、三或四个密码子代表。显然,密码子的分配是非随机的。还可看到在头两个碱基相同的密码子当中,半数是为同一氨基酸编码,亦即第三位碱基无论其为A、G、U、或C,均代表同一氨基酸。因此tRNA的反密码子第一位碱基(和密码子第三位碱基配对)为次黄嘌呤而能与C以及U和A配对(与后二者的配对是非Watson-Crick的摆动),亦无害于翻译之忠实性(参见“摆动配对”)。一种氨基酸由一个以上密码子编码,在密码术语上叫做简并,所以遗传密码具有简并性(参见“简并性”)。这些密码子的破译起初是由大肠杆菌的蛋白质合成体系作出的; 现有各种确凿证据表明所有生物,从病毒、细菌直到哺乳动物,大都使用表上所列同一的密码,这就是遗传密码的通用性 (参见 “通用性”)。
遗传密码字典
| 5'端 碱基 | 中 间 位 碱 基 | 3'端 碱基 | |||
| U | C | A | G | ||
| U | 苯丙氨酸 苯丙氨酸 亮氨酸 亮氨酸 | 丝氨酸 丝氨酸 丝氨酸 丝氨酸 | 酪氨酸 酪氨酸 终止 终止 | 半胱氨酸 半胱氨酸 终止⊕ 色氨酸 | U C A G |
| C | 亮氨酸 亮氨酸 亮氨酸 亮氨酸 | 脯氨酸 脯氨酸 脯氨酸 脯氨酸 | 组氨酸 组氨酸 谷氨酰胺 谷氨酰胺 | 精氨酸 精氨酸 精氨酸 精氨酸 | U C A G |
| A | 异亮氨酸 异亮氨酸 异亮氨酸⊕ 甲硫氨酸* | 苏氨酸 苏氨酸 苏氨酸 苏氨酸 | 天冬酰胺 天冬酰胺 赖氨酸 赖氨酸 | 丝氨酸 丝氨酸 精氨酸⊕ 精氨酸⊕ | U C A G |
| G | 缬氨酸 缬氨酸 缬氨酸 缬氨酸* | 丙氨酸 丙氨酸 丙氨酸 丙氨酸 | 天冬氨酸 天冬氨酸 谷氨酸 谷氨酸 | 甘氨酸 甘氨酸 甘氨酸 甘氨酸 | U C A G |
*起始密码子。作为起始密码子时,GUG为甲硫氨酸编码。
 在哺乳动物线粒体,UGA是色氨酸密码子;AUA是甲硫氨酸密码子;AGA和AGG是终止信号。
在哺乳动物线粒体,UGA是色氨酸密码子;AUA是甲硫氨酸密码子;AGA和AGG是终止信号。遗传密码
遗传密码是指蛋白质生物合成中,从信使核糖核酸(mRNA)的核苷酸顺序翻译成蛋白质的氨基酸顺序所使用的密码,即遗传信息的单位。遗传密码是由DNA和mRNA中,三个相连的核苷酸组成的三联体。每一个三联体是编码一个氨基酸的密码子,譬如,编码一个有500个氨基酸的蛋白质遗传信息包含在相当于1 500个核苷酸的500个密码子中。密码子的顺序决定合成的多肽链的顺序。
已知核酸是由腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)四种不同碱基的核苷酸组成的,而蛋白质通常是由20种不同的氨基酸组成的。从组合的可能性考虑。每个氨基酸应由至少3个相连的核苷酸顺序决定。因为,在四种核苷酸中,以三个为一组的全部可能的排列为43=64,已超过了编码20种氨基酸所需的数量。Crick在60年代初期,根据对T4噬菌体移框突变的观察结果,推想密码子是相连的三个核苷酸顺序。继之,Nirenberg、Ochoa和Korana实验室利用在无细胞系统中加入人工合成的mRNA指导蛋白质合成,以及人工合成的三核苷酸和核糖体专一结合等方法,来破译遗传密码。例如,加多聚U时的翻译产物是多聚苯丙氨酸。所以苯丙氨酸的密码子是UUU。GCC只能把丙氨酰—tRNA结合到核糖体上,而不能把任何其他氨酰—tRNA结合到核糖体上,因而表明GCC是丙氨酸的密码子。利用这些方法,互相引证,终于在1966年破译了全部遗传密码(表)。70年代期间,基因之核苷酸顺序与所编码的蛋白质之氨基酸顺序的对应性的阐明,更直接证明了这个密码表的正确性。
遗传密码表(由4种氨基酸编码的64种密码子)
*是终止密码子
所有已知生物的遗传密码都具有下列共同的特性:
❶三联体密码:每一个氨基酸都是由相连的三个核苷酸编码的;
❷通用性:所有已知生物都具有相同的密码系统;
❸简并性:某些氨基酸可能不只由一个而是由几个密码子决定:
❹不重叠性:前后两个密码子互不重叠,没有兼用的核苷酸;
❺无标点:在一个蛋白质的多肽链的终点存在着终止密码,而多肽链内各个氨基酸之间则不存在作为标点的密码。蛋白质合成中,密码的阅读是沿mRNA的5′端到3′端的方向,依次进行的。
在64个密码子中,61个编码特定的氨基酸,剩余的3个密码(UAG,UAA,UGA)不编码任何氨基酸,而是作为多肽链合成的终止信号。多肽链合成的起始密码是AUG,它在原核生物编码N-甲酰甲硫氨酸,在真核生物则编码甲硫氨酸。密码有高度的简并性,即一种氨基酸可能有几种同义密码,也就是说一种tRNA可能专一地识别一种以上的密码子。简并总是出现在密码子3′端的第三个碱基位置上。这是由于tRNA的反密码子之5′端的碱基在和密码子配对时,倾向于“摇摆”的结果(见“转移核糖核酸”条)。大多数氨基酸的同义密码以及起始密码和终止密码,已从对R17,f2和MS2病毒的RNA和珠蛋白mRNA的核苷酸顺序与其翻译产物的氨基酸顺序的直接对比研究中得到证明(见“信使核糖核酸”条图3)。从原核生物到真核生物(植物、动物和人),遗传密码几乎都是通用的。例如,把大肠杆菌的各种氨酰—tRNA加入到家兔网织红细胞核糖体(含有珠蛋白mRNA)无细胞蛋白质合成系统中,就能合成珠蛋白。这表明细菌tRNA的反密码子能识别珠蛋白mRNA的密码。同样地,任何物种的tRNA和另一物种的mRNA配合的类似实验,在迄今所有试验过物种,如大肠杆菌、豚鼠、爪蟾等之间都得到成功。遗传密码的通用性证明了生物界的统一性和共同起源。然而,不同物种的一个特定氨基酸使用不同的简并密码的频率则各不相同。这一事实提示氨基酸密码的多样性可能经历过一个进化和分化的过程。遗传密码的起源和进化是生物学目前的一个重要研究课题。
有些化学物质的作用能改变遗传密码,从而诱发基因突变,因而称为诱变剂。一些诱变剂(如亚硝酸,羟胺)能使DNA的一种碱基改变成另一种碱基 (如亚硝酸能使腺嘌呤经过氧化脱氨转化为次黄嘌呤)。另一些诱变剂是碱基结构类似物(如5-溴尿嘧啶,2-氨基蝶呤,5-溴脱氧尿嘧啶和5-氟脱氧尿嘧啶),在复制时能参入DNA,造成碱基配对的差错。烷化剂(如环氧乙烷,氮芥)能引起DNA碱基脱落。某些荧光染料(如吖啶,前黄素)能嵌入DNA或引起碱基缺失,从而阻断遗传信息的阅读。由于以上这些作用,诱变剂也能抑制细胞分裂和增生。诱变剂的这些特性在医学上已应用于癌的化学治疗。
遗传密码
genetic code
遗传密码
genetic code
遗传密码
genetic code
- 孙鼎和是什么意思
- 孙鼎宜是什么意思
- 孙鼎臣是什么意思
- 孙鼐是什么意思
- 孙鼐是什么意思
- 孙龙是什么意思
- 孙龙基是什么意思
- 孙龙父是什么意思
- 孙龙父书法篆刻选是什么意思
- 孙龙花鸟草虫册是什么意思
- 孚是什么意思
- 孚中是什么意思
- 孚化志略是什么意思
- 孚尔根染色法是什么意思
- 孚念孚是什么意思
- 孚泽庙是什么意思
- 孚琪乳膏是什么意思
- 孚赫,尤利乌斯(茹尔)是什么意思
- 孚远县是什么意思
- 孚远县乡土志是什么意思
- 孚雷是什么意思
- 孛儿帖赤那是什么意思
- 孛斡勒是什么意思
- 孛星是什么意思
- 孛术鲁翀是什么意思
- 孛术鲁阿鲁罕是什么意思
- 孛来是什么意思
- 孛秃是什么意思
- 孛罗帖木儿是什么意思
- 孛罗帖木儿是什么意思
- 孛罗帖木儿是什么意思
- 孛罗贴木儿是什么意思
- 孜亚伊是什么意思
- 孜代克西比赫是什么意思
- 孜孜上进心是什么意思
- 孜孜不倦是什么意思
- 孜孜不倦是什么意思
- 孜孜不倦是什么意思
- 孜孜不倦是什么意思
- 孜孜不倦是什么意思
- 孜孜不息是什么意思
- 孜孜矻矻是什么意思
- 孜拉布扎是什么意思
- 孜然芹是什么意思
- 孜然芹是什么意思
- 孜牙墩是什么意思
- 孜牙墩是什么意思
- 孜牙敦是什么意思
- 孝是什么意思
- 孝是什么意思
- 孝是什么意思
- 孝是什么意思
- 孝是什么意思
- 孝与中国文化是什么意思
- 孝丰县志是什么意思
- 孝为百行之首是什么意思
- 孝义县志是什么意思
- 孝乎惟孝,友于兄弟是什么意思
- 孝仁皇后(董氏)是什么意思
- 孝仪纯皇后(魏佳氏)是什么意思