转运核糖核酸(tRNA)
转运核糖核酸广布于各种细胞的胞液中,以前曾被称为可溶性RNA (sRNA),含量约占细胞总RNA的15%,以自由状态或与氨基酸结合的形式而存在。tRNA是分子量最小的一类RNA,从各种细胞中分离提纯并确定了核苷酸序列的tRNA已有一百多种,它们都是由74~94个核苷酸组成,以含稀有核苷酸较多为其特点,分子量约为2.3万~3.0万道尔顿,沉降常数为4S,故又常被称为4S RNA。tRNA的主要功能是在蛋白质生物合成中起转运氨基酸的作用,在特异的氨基酰tRNA合成酶的协助下,一种tRNA只能转运一种特异的氨基酸,但一种氨基酸却可被几种tRNA转运,这样的tRNA称为同功tRNA,例如大肠杆菌中能转运缬氨酸的同功tRNA有三种,能转运亮氨酸的同功tRNA有五种。可见用以合成蛋白质的氨基酸虽然只有20种,但转运这些氨基酸的tRNA则远不止此数; 实验证明,由一种细胞分离出的4S RNA制剂中往往含有60~80种tRNA,它们的结构和功能有共性也有特性。
一级结构 tRNA分子的组成单位主要是含腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)四种碱基的相应核苷酸,以及少量从它们修饰而来的多种稀有核苷酸。一般每分子tRNA由80个左右此等核苷酸聚合成一条多核苷酸链,链中各种核苷酸的含量没有一定的比例关系,其中稀有核苷酸之量约占5~20%;各个核苷酸之间都是通过3′,5′-磷酸二酯键而彼此相连; 至于分子中的核苷酸排列顺序,现在有多种tRNA已被搞清。Holley等于1965年用所谓片段重叠法首先测出了面包酵母丙氨酸tRNA的核苷酸序列(图1),这是世界上第一种被搞清一级结构的核酸。随后Sanger等在测定核蛋白体5S-RNA的核苷酸序列时又发展和改进了这种方法,至今仍被广泛采用,并适用于各类RNA的序列分析。此法的基本原理和步骤大致如下:
❶用两种或两种以上不同的核糖核酸酶分别将RNA或32P标记的RNA特异地完全水解成若干个小片段。最常用的酶是核糖核酸酶A及核糖核酸酶T1,前者来自牛胰,能特异地水解RNA链中嘧啶核苷酸的3′磷酸基与相邻核苷酸的5′羟基之间的磷酸酯键,产生游离的3′-嘧啶核苷酸和以它为末端基的寡核苷酸片段; 后者来自米麦曲霉菌,能特异地水解RNA链中鸟苷酸的3′磷酸基与相邻核苷酸的5′羟基之间的磷酸酯键,生成游离的3′-鸟苷酸和以它为末端的寡核苷酸片段。
❷用上述核糖核酸酶T1或A将RNA部分水解,产生分子较大的小片段,用DEAE—纤维素或DEAE—葡聚糖凝胶柱层析法或用醋酸纤维—DEAE纸双相电泳法很易分开;
❸经部分水解后产生的大片段可用聚丙烯酰胺凝胶电泳法分开。
❹通过碱水解、酶水解、纸电泳、纸层析以及对紫外吸收和比放射性的检测等手段,可以分析出各个小片段的相对含量、核苷酸组成及其排列顺序。
❺由于特异性不同的核糖核酸酶水解RNA时的切割点不同,产生的小片段自然也就不同,比较这些小片段的核苷酸序列,必可找到许多重叠的节段,从而可以确定各个小片段在大片段中的位置,进而推算出各大片段及整个RNA分子的核苷酸排列顺序。图1(A)为Holley等利用此法于1965年首次确定的酵母丙氨酸tRNA的77个核苷酸的序列,以及Penswick等于1975年修订后的76个核苷酸的序列,其差别仅在于后者缺少原第13及14位的鸟苷酸和胞苷酸,并在原第47和48位之间加进了一个鸟苷酸。我国科学工作者经过10年努力,已于1979年底人工合成了此tRNA由41个核苷酸组成的3′-端大半个分子;1982年初,中国科学院生物化学研究所宣布酵母tRNA的全分子的人工合成已经全部完成。除酵母丙氨酸tRNA外,现在有百多种tRNA的一级结构已被确定,从中发现凡是同功tRNA的核苷酸序列都非常近似,例如酵母丝氨酸的两种同功tRNA,在其85个核苷酸的序列中只有三个核苷酸不同(图1 (B));大肠杆菌缬氨酸的三种同功tRNA的76个核苷酸序列中只有六个核苷酸不同。但各种非同功tRNA的核苷酸组成及排列顺序则差别较大,不过一般也都只有50%左右不同。这种现象颇可显示结构与功能的一致性。
二级结构 tRNA的多核苷酸链可以自身折迭而形成三叶草形二级结构,各种tRNA共有的此种二级结构可用图2表示。它包括五个折迭区,通常称其为五个臂,即氨基酸臂、二氢尿嘧啶臂 (简称DHU臂或D臂)、反密码臂、可变臂 (又称额外臂) 和胸苷假尿苷胞苷臂(简称TψC臂)。每个臂又由两个部分组成· 一个部分中的碱基可以互补成几个碱基对(3~7对),并形成一个小双螺旋,通常称为“茎”; 另一个部分中的碱基不能互补成碱基对,而是以单链形式存在,并往往形成一个小环而与茎部相连。这样的臂实际上就是通常所谓的发夹式结构。任何tRNA分子中的D臂、反密码臂及TψC臂都是典型的发夹式结构,但氨基酸臂中单链部位则不形成环,可变臂中有的不仅不能形成环,甚至连小双螺旋也不能形成。tRNA在蛋白质生物合成中转运氨基酸时,其分子中各个臂的茎部可能与特异的氨基酰tRNA合成酶的结合有关,而各个臂的非茎部位则多具有独特的功能。例如氨基酸的非茎部位为不能形成环的……嘌—C—C—A,它是所有各种tRNA共同的3′末端核苷酸序列,在转运氨基酸时都是其中腺苷酸2′-羟基或3′-羟基与特定氨基酸的羧基缩合成酯键而生成相似的氨基酰tRNA; 此外,过碘酸法及酶解法都证实,氨基酸臂中—C—C—A的存在对于tRNA与核蛋体大亚基的结合是必不可少的。D臂中的非茎部分(D环)一般含7~10个核苷酸,此环对于维系tRNA三级结构的稳定甚为重要。反密码臂中的非茎部位(反密码环)一般都含七个核苷酸,环中央位置上的三个核苷酸称为反密码子,能识别mRNA中特异氨基酸的密码子并能与之互补结合,如此则可保证以mRNA中按特定的遗传信息翻译出具有特定氨基酸序列的蛋白质; 此外,当tRNA的反密码子与mRNA中相应的密码子互补结合后,尚可诱导引起tRNA分子的构象改变,促使tRNA通过其TψC环而与核蛋白体中5S rRNA的互补结合 (见“核蛋白体核糖核酸”条)。各种tRNA的可变臂所含核苷酸的数量相差甚远,一般为3~21个,含量多者可形成小双螺旋,含量少者则不能。可变臂中的核苷酸数与D臂的D茎中的核苷酸数之间存在一定关系,例如D茎为四对核苷酸(D4)时,可变臂往往含五个核苷酸(V5),若可变臂含13~21个核苷酸时(V13-21),D茎中一定都含三对核苷酸(D3)等。据此,所有天然存在的tRNA都可归纳为下列四类:
❶D4V5,
❷D3V5,
❸D3-4V4,
❹D3V13-21。其中以第
❶类最多,约占50%以上; 第4类其次,约占25%; 第
❷及第
❸类较少,总共约占25 %以下。这种关系的意义以及可变臂本身的功能至今尚不清楚。TψC臂的TψC环(环IV)含七个核苷酸,所有原核细胞的各种tRNA以及真核细胞中除起动tRNA外的其他各种tRNA的此臂中均含—G—T—ψ—C—序列,而原核细胞的核蛋白体大亚基的5S rRNA以及真核细胞核蛋白体大亚基的5.8S rRNA中都含—G—A—A—C—序列,此二种序列可以互补配对以实现tRNA与核蛋白体大亚基的结合,此种结合是tRNA转运氨基酸时的必要条件之一; 此外,TψC环还能维系和稳定tRNA的三级结构。真核细胞起动tRNA(tRNAimet)不同于原核细胞起动tRNA (tRNAfmet),其环IV中不含—G—T—ψ—C—而含—G—A—U—C—序列,但在真核细胞的核蛋白体大亚基的5S rRNA中也含—G—A—U—C—序列,二者可以互补配对以实现tRNAimet与核蛋白体的结合。总之,在真核细胞中,其起动tRNA与核蛋白体的5S rRNA相互结合是蛋白质合成中起动作用的重要条件,而其他tRNA与核蛋白体的5.8S rRNA相互结合则是肽链延伸作用的重要条件。但在原核细胞中,无论是起动作用或延伸作用,都仅仅与其各种tRNA和核蛋白体的5SrRNA相互结合有关。
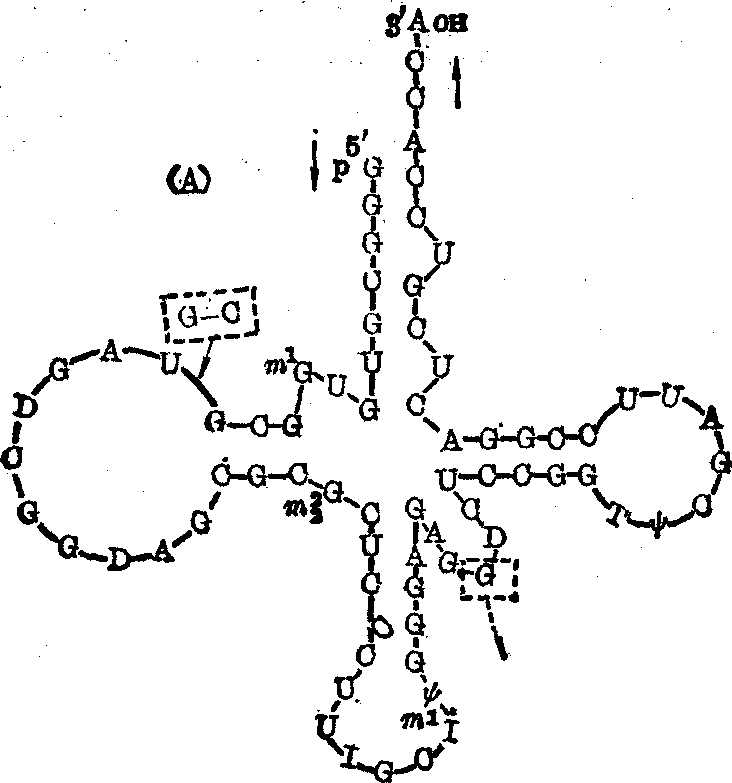
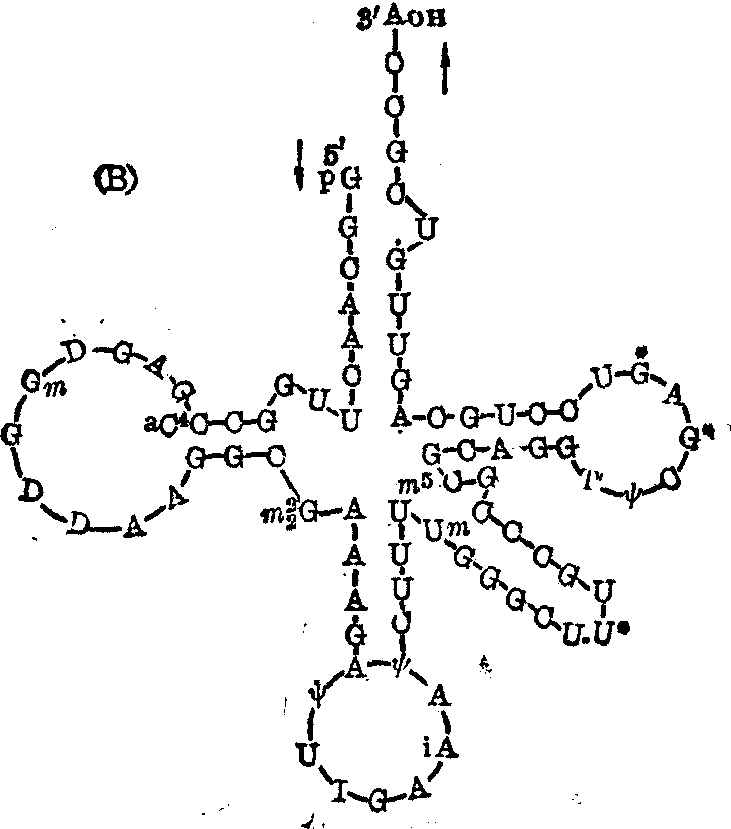
图1 tRNA的核苷酸排列顺序
A.酵母丙氨酸tRNA; B. 酵母丝氨酸tRNA2,其中带*的U改为C,G改为A后,即成为其同功tRNA1
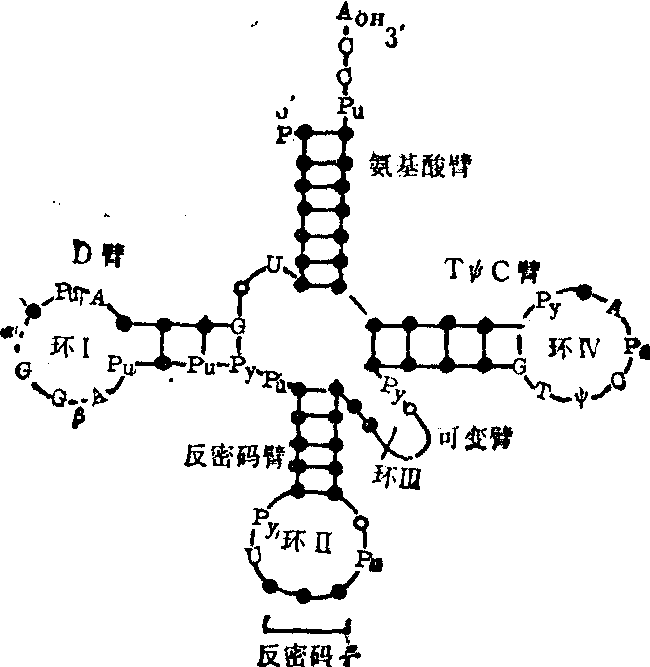
图2 tRNA的三叶草形二级结构
各部位中指出的特定核苷适用于除真核起动tRNA的其他所有tRNA。Pu及Py代表嘌呤核苷及嘧啶核苷;α及β代表1~3个核苷,其中主要是D,其他说明见正文三级结构 tRNA三叶草形二级结构的某些部位之间通过一定的空间联系可形成三级结构。1974年Kim等人和Robenrtus等人对高纯度的酵母苯丙氨酸tRNA同时分别进行了高分辨率的X线衍射分析,得到了相同的结果,一致确定了酵母tRNA苯丙的三级结构 (图3)。此结构有下列几个特点:
❶整个tRNA分子的外形如同一个倒写的L字母或近似一个字母T,L的一端为氨基酸臂,另一端为反密码环,L的拐角处则由TψC环及D环的一部分组成;
❷氨基酸臂的茎部与TψC茎相接而形成一个连续的较长的双螺旋(12个碱基对);
❸D茎和反密码茎相接而形成另一个连续的双螺旋(9个碱基对)其中D茎由于额外生成了U
 ·A14和G15·C4
·A14和G15·C4 新碱基对而增加了双螺旋的长度;
新碱基对而增加了双螺旋的长度; ❹这两个双螺旋彼此垂直;
❺二级结构中的某些原单链上的核苷酸之间可通过三级氢键的生成而形成若干个新的碱基对或碱基三联体(图4),以保持和稳定该三级结构。例如在酵母tRNA苯丙中(图4)可生成第三级新碱基对的有U
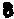 和A14、G15和C48、G1
和A14、G15和C48、G1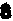 和ψ55、 G18和C56、 m2G10和G45、m22G26和A44、 T54和m1A58;可生成碱基三联体的有U12A23和A9及C13·G22和m7G46。
和ψ55、 G18和C56、 m2G10和G45、m22G26和A44、 T54和m1A58;可生成碱基三联体的有U12A23和A9及C13·G22和m7G46。稀有核苷及其功能 与其他各类RNA相比,tRNA的特点之一是其分子中含有多种稀有核苷(酸),其量约占总核苷数的5~20%。例如由76个核苷酸组成的酵母tRNA丙中有10个是稀有核苷,由85个核苷酸组成的酵母tRNA丝中有14个是稀有核苷,由85个核苷酸构成的鼠肝tRNA丝中含17个稀有核苷,由75个核苷酸组成的真核细胞起动tRNA中有8个是稀有核苷等。这些稀有核苷都是在tRNA全分子水平上由通常的四种基本核苷(A、G、C、U)经酶促修饰而成,修饰作用主要是通过甲基化、硫化以及其他某些特殊反应。修饰部位可发生在基本核苷的碱基上,生成m1A、m6A、m62A、m7G、i6A、m5S2U等; 或发生在基本核苷的核糖上,生成各种2′-o-甲基核苷,即Am、Gm、Um、Cm等; 也可同时发生在碱基及核糖之上,生成m5Um、m4Cm等; 此外,修饰作用还可能发生在碱基和核糖的连接方式上,生成ψ和ψm等。tRNA分子中的稀有核苷大多分布在其多核苷酸链的特定部位上,而且位于单链部位 (各个臂的连接处及环上)中的多于位于双螺旋(各个臂的茎)部位中的; 真核细胞tRNA中的多于原核细胞tRNA中的。就现在所知,所有tRNA的反密码3′端相邻的核苷酸(常称为“反密码3′边”核苷酸)都是嘌呤核苷酸,而且其中的核苷只有极少数是未被修饰的腺苷,绝大多数情况下是单纯甲基化的稀有核苷,如m1G、m6G、m2A、m1I等或高度修饰的稀有核苷,如i6A、ms2i6A、t6A、m6t6A、Y、Yt等。某些这类稀有核苷的结构式如图6。实验证明,tRNA反密码3′-边核苷的修饰度愈高 (例如ms2i6A>i6A>mA),则该tRNA与mRNA及核蛋白体的结合能力愈大,促进蛋白质生物合成的效能愈高。此外还发现,tRNA的反密码3′-边稀有核苷若为i6A或ms
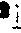 6A时,此类反密码必定都识别以U起始的密码子;若为t6A或mt6A时则必定都识别以A起始的密码子;若为m2A、m1G或A时则主要识别以G起始的密码子。这种现象似乎显示反密码3′-边稀有核苷对于监督密码子与反密码子的准确结合起一定的作用。此外tRNA反密码子本身的第二个和第三个核苷酸 (从5′端数起) 虽然都是非修饰核苷,但其第一个核苷大多为稀有核苷, 其中以次黄苷 (I)最多, 其次是m
6A时,此类反密码必定都识别以U起始的密码子;若为t6A或mt6A时则必定都识别以A起始的密码子;若为m2A、m1G或A时则主要识别以G起始的密码子。这种现象似乎显示反密码3′-边稀有核苷对于监督密码子与反密码子的准确结合起一定的作用。此外tRNA反密码子本身的第二个和第三个核苷酸 (从5′端数起) 虽然都是非修饰核苷,但其第一个核苷大多为稀有核苷, 其中以次黄苷 (I)最多, 其次是m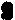 A、 m5C、Cm、Gm. N4-乙酰胞苷以及下列一些高度修饰的稀有核苷(图7)。根据Crick的摆动学说,tRNA反密码子的第一个核苷(通称摆动核苷或变偶核苷)可以通过适当摆动或扭转而与mRNA中密码子的第三个核苷进行非典型的互补配对,通称变偶碱基对。例如反密码子中的变偶核苷U既可与密码子中的第三核苷A互补结合 也可与第三核苷G配对结合; 同样,G可与C或U互补结合, I(次黄苷)可与C或U或A互补配对,核苷Q (图7) 可与U或C配对结合等。通过变偶作用,一种氨基酸tRNA的反密码子可与mRNA的几种密码子互补结
A、 m5C、Cm、Gm. N4-乙酰胞苷以及下列一些高度修饰的稀有核苷(图7)。根据Crick的摆动学说,tRNA反密码子的第一个核苷(通称摆动核苷或变偶核苷)可以通过适当摆动或扭转而与mRNA中密码子的第三个核苷进行非典型的互补配对,通称变偶碱基对。例如反密码子中的变偶核苷U既可与密码子中的第三核苷A互补结合 也可与第三核苷G配对结合; 同样,G可与C或U互补结合, I(次黄苷)可与C或U或A互补配对,核苷Q (图7) 可与U或C配对结合等。通过变偶作用,一种氨基酸tRNA的反密码子可与mRNA的几种密码子互补结合,这对于加速蛋白质合成的翻译过程显然有利。
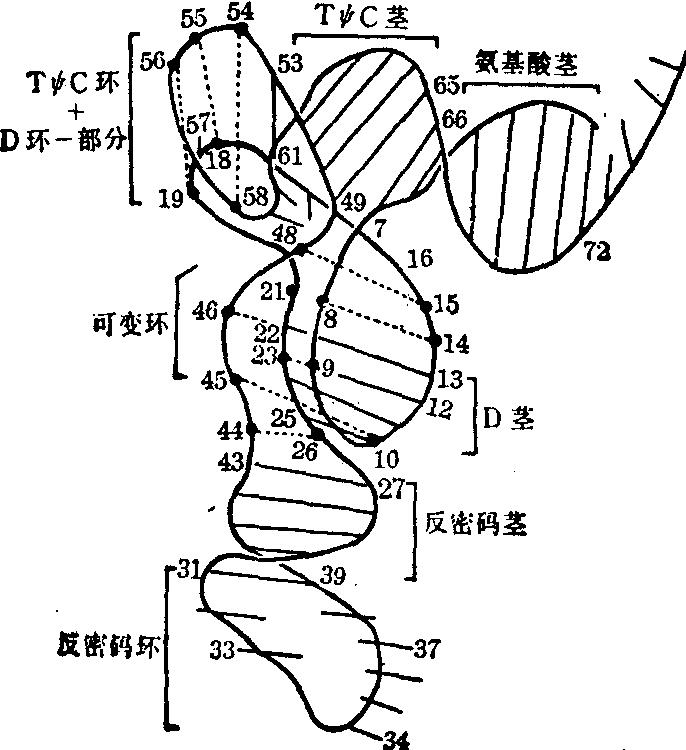
图3 酵母苯丙氨酸tRNA的三级结构
连续粗线表示磷酸核糖骨架,细长线表示双螺旋基中的碱基对,虚线表示三线结构中形成的新碱基对,短细线代表不配对碱基
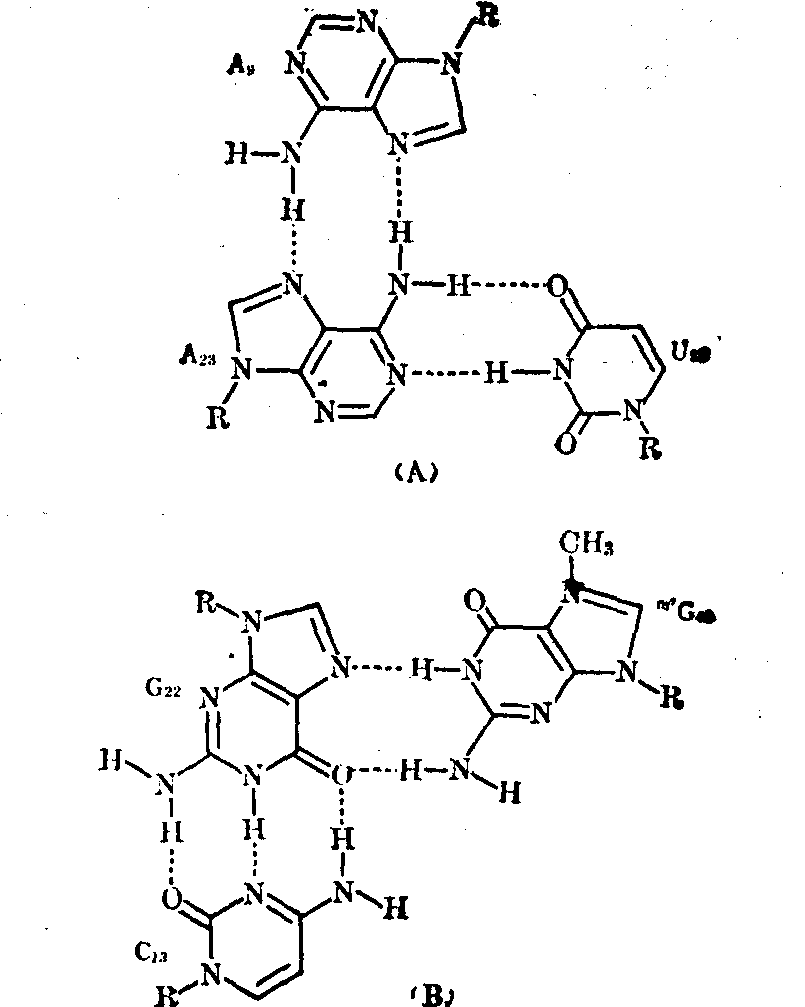
图4 酵母苯丙氨酸tRNA三级结构中的碱基三联体A.U12·A23·A B.C15·G2
B.C15·G2 ·m,G4
·m,G4
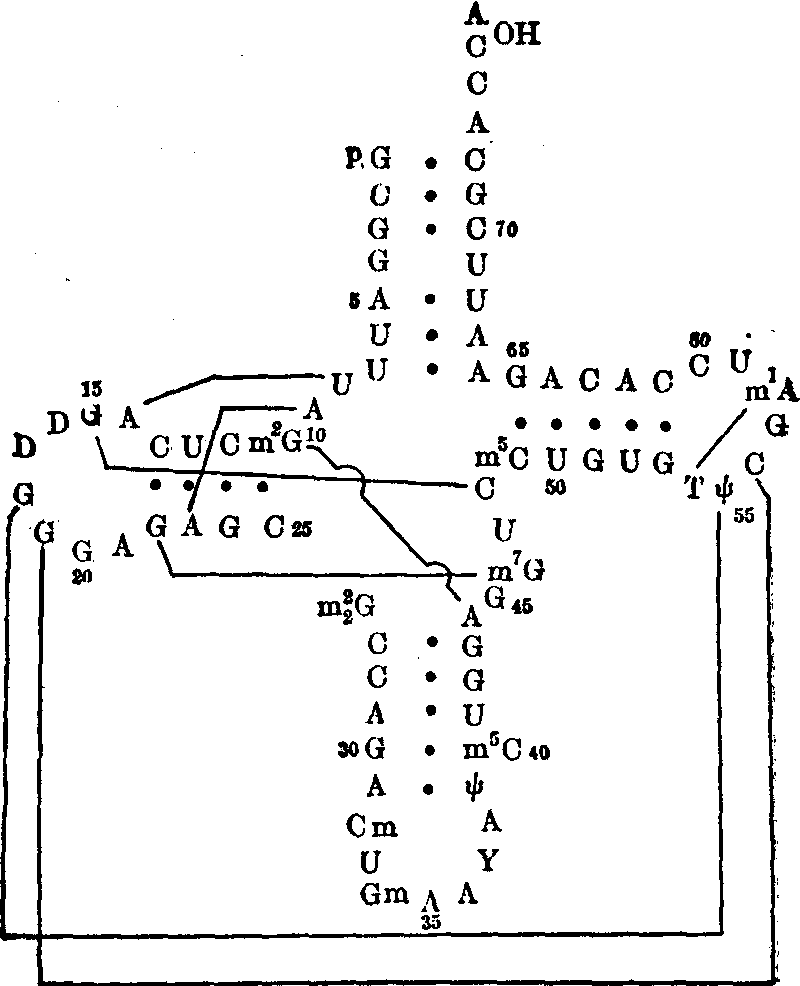
图5 以三叶草形二级结构排列的酵母苯丙氨酸tRNA的核苷酸序列
双螺旋基中的碱基对氢键用大圆点表示,三级结构中新碱基对氢键用细长线表示,以便与图比较
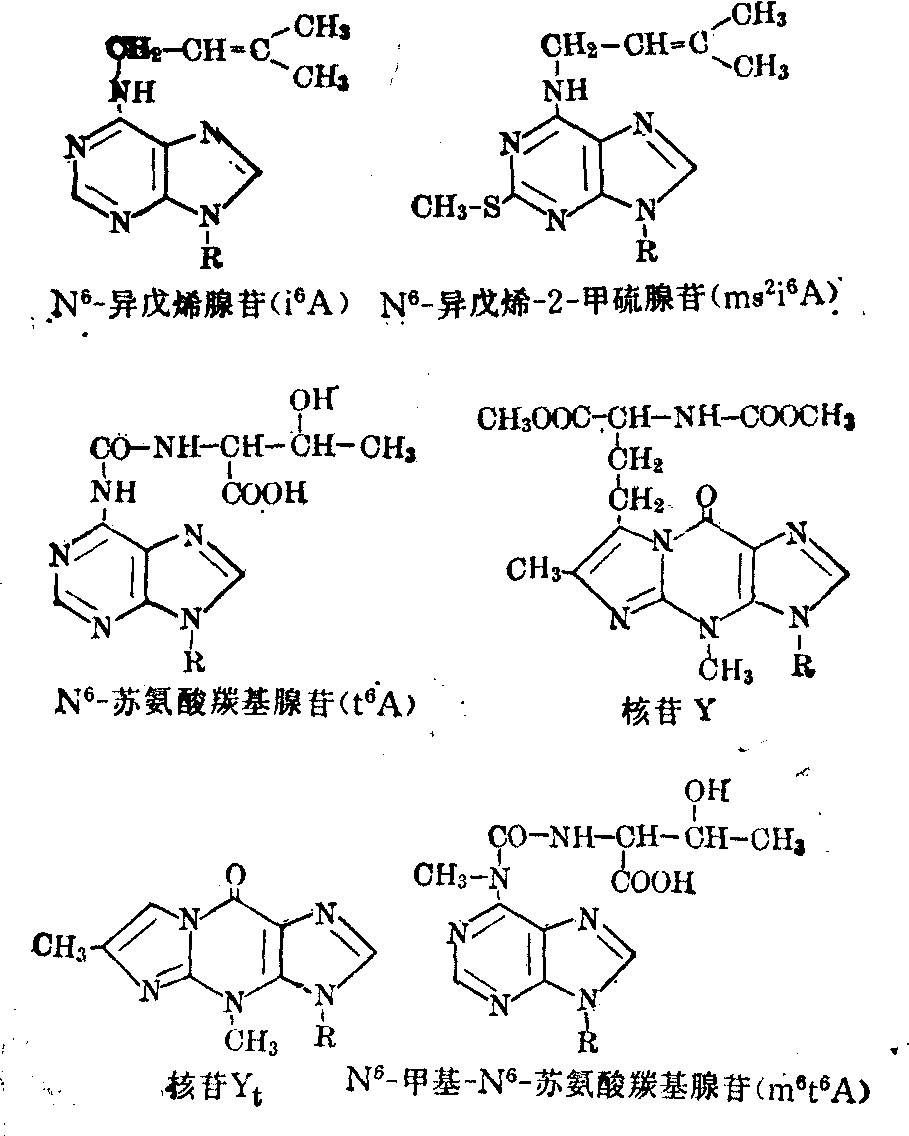
图6 “反密码3′-边”的某些稀有核苷
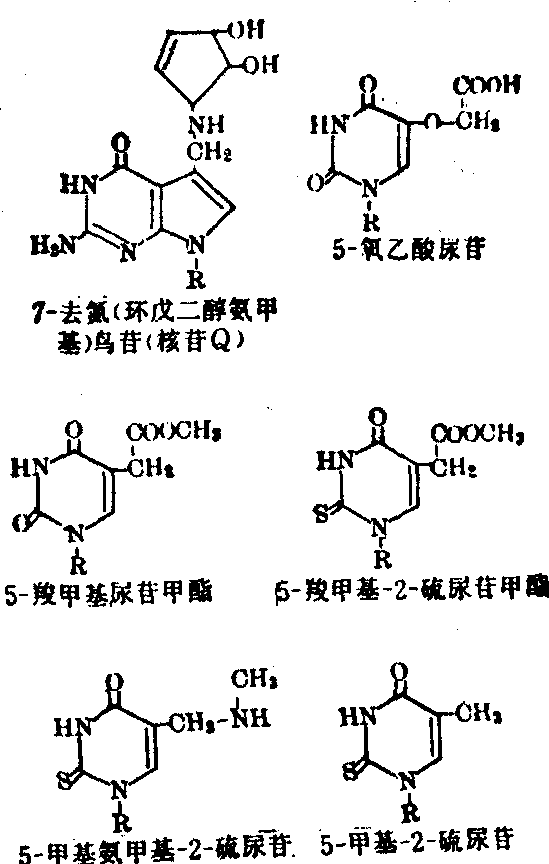
图7反密码中第一位(5′端)的某些稀有核苷
- 做错了事,怕人揭疤; 见人低语,以为说他是什么意思
- 做长工是什么意思
- 做长活的是什么意思
- 做长远的安排是什么意思
- 做门户是什么意思
- 做门面花是什么意思
- 做门面货是什么意思
- 做闹是什么意思
- 做闹热是什么意思
- 做阔是什么意思
- 做队是什么意思
- 做阴是什么意思
- 做阴寿是什么意思
- 做阵是什么意思
- 做阵农是什么意思
- 做陴是什么意思
- 做难是什么意思
- 做雅粿是什么意思
- 做雅馃是什么意思
- 做集是什么意思
- 做雨是什么意思
- 做雨天是什么意思
- 做雪是什么意思
- 做零工是什么意思
- 做零工的人是什么意思
- 做零活儿的报酬是什么意思
- 做零活,打短工是什么意思
- 做霉是什么意思
- 做霜是什么意思
- 做霜构是什么意思
- 做青是什么意思
- 做非凡之事是什么意思
- 做面是什么意思
- 做面做目是什么意思
- 做面子是什么意思
- 做面孔是什么意思
- 做面皮是什么意思
- 做面目是什么意思
- 做面线是什么意思
- 做鞋是什么意思
- 做鞋佬是什么意思
- 做鞋则是什么意思
- 做项是什么意思
- 做顺水人情是什么意思
- 做顺莫是什么意思
- 做领袖要学会隐藏恐惧是什么意思
- 做额子是什么意思
- 做风是什么意思
- 做风做涌是什么意思
- 做风台是什么意思
- 做风水是什么意思
- 做风灾是什么意思
- 做食是什么意思
- 做食品的猪羊大肠是什么意思
- 做饭是什么意思
- 做饭个是什么意思
- 做饭做菜是什么意思
- 做饭厦是什么意思
- 做饭哩是什么意思
- 做饭处是什么意思