肌肉收缩的能量来源
肌肉收缩的直接能量来源是ATP。但肌肉中ATP的浓度仅约8mmol/L,此贮存量仅够维持收缩半秒(即10次)左右。尽管如此,中等强度的肌肉收缩并不引致组织内ATP含量明显降低。这是由于磷酸肌酸可及时为ADP的磷酸化提供方便的高能磷酸基。当然,肌肉收缩的最终能量来源是物质代谢。糖类及脂类的分解代谢均可为肌肉活动提供ATP,不过不同种类或不同生理及病理状况下的肌肉产生ATP的代谢途径有所不同。
骨胳肌的红纤维含有丰富的线粒体、肌红蛋白及细胞色素,因而氧化脂肪酸、酮体及丙酮酸的能力很强; 主要通过氧化磷酸化作用为其收缩提供ATP。白纤维则相反,线粒体及肌红蛋白均少,而酵解酶体系活性很强,故消耗糖原的无氧酵解是为其收缩提供ATP的主要代谢途径。
静止的肌肉需要一定量ATP以维持其稳定的组成及连续的代谢。肌肉静止时靠氧化脂肪酸产生ATP,消耗葡萄糖很微少。肌肉收缩时能量的需要立即增加20~200倍。收缩初始时由于氧的供不应求主要由糖原的无氧酵解来满足瞬息增高的能量需要。在肌肉活动过程中肌细胞对氧及葡萄糖的摄取增多,同时肝细胞释放葡萄糖加速,如果活动强度不大,持续时间不久。糖的有氧氧化是其供能的重要来源。然而哺乳动物肌肉在最大限度活动时,每克组织每分钟耗用ATP约1mmol。在这种情况下氧的供应远不能满足需要。实验证明在激烈运动,如短时间疾跑时,乳酸大量生成,肌糖原甚至可以耗尽。足见,糖酵解作用是激烈运动时为其提供ATP的主要途径。此时所产生的大量乳酸需待静止之后由肌肉及其他组织氧化,或由肝脏转变为糖原 (糖异生作用,糖异生所需要的能量由部分乳酸的氧化提供)。以致在运动后的一定时期,机体仍然保持较快的呼吸及较高的氧消耗。例如一个赛跑者以最大速度疾跑10秒钟,当时约消耗1L氧(静息时在同等时间内仅消耗40ml氧),停跑后在恢复期还要额外消耗4L氧。即一次短跑所需要的氧有4/5是在运动停止后消耗的。这种在运动恢复期的额外耗氧量称为氧债。再者,由肌细胞生成的乳酸经血流运至肝脏通过糖异生作用合成的糖原又可分解为葡萄糖,再经血流回到肌肉。这个过程称为Cori循环。
肌肉组织通过一系列复杂而协调的代谢机构,以满足肌肉活动时大量的能量需要,例如肌肉收缩时通过4条途径加速糖的分解:
❶葡萄糖透过细胞膜的转运增强;
❷磷酸化酶活性增高,糖原分解加快;
❸已糖激酶活性增大,葡萄糖的磷酸化加快;
❹磷酸果糖激酶活性增大。此酶为糖酵解作用的限速酶。已知肌肉兴奋时肌质中Ca2+浓度增加。Ca2+不仅通过TN-C及其他调节蛋白质和肌动蛋白活化了肌球蛋白ATP酶,启动了肌肉收缩;同时还具有活化磷酸化酶激酶的作用。从而加速了糖原的分解,为肌肉收缩提供能量(图1)。亦有报道,Ca2+可活化己糖激酶及磷酸果糖激酶。此外,肌肉激烈活动可引起ATP/ADP比值降低,通过变构调节而增强磷酸果糖激酶活性,从而加速糖酵解。
肌肉中广泛存在着腺苷酸激酶又名肌激酶。肌肉活动时ADP浓度增高,该酶催化二分子ADP之间的磷酸基转移, 产生ATP及AMP。 2ADP⇌ATP+AMP(K=1)。AMP是磷酸果糖激酶的激活变构剂;并可直接活化磷酸化酶b,因而AMP的生成亦可加速糖酵解。然而,过多的AMP对肌肉具有毒害作用,有人认为它可使肌肉发生僵直; 幸而肌肉中含有腺苷酸脱氨酶,可催化AMP脱去氨基产生次黄嘌呤核苷酸IMP(肌苷酸)及氨,氨可缓冲肌肉中的乳酸。必要时IMP可通过与门冬氨酸及GTP作用再合成AMP。AMP不能直接磷酸化产生ADP,但可通过与ATP交换高能磷酸基而生成ADP。实验证明,当进行中等强度的持久肌肉活动时,呼吸商逐渐降低。这表明随着活动时间的延长,脂类物质氧化供能所占的比重越来越大。肌肉所利用的脂类包括肌细胞内贮存的甘油三酯以及从血中摄取的游离脂肪酸、酮体及甘油酯化脂肪酸。持久运动时血脂肪酸浓度升高。再者,肌肉中脂肪酸及酮体的氧化可以抑制糖的酵解。反之,当葡萄糖氧化增多时(如在胰岛素作用下),磷酸甘油的生成增多,从而使脂肪酸较多进行酯化而较少进行氧化,结果细胞内甘油三酯增多。脂肪酸与葡萄糖代谢之间的联系见图2。
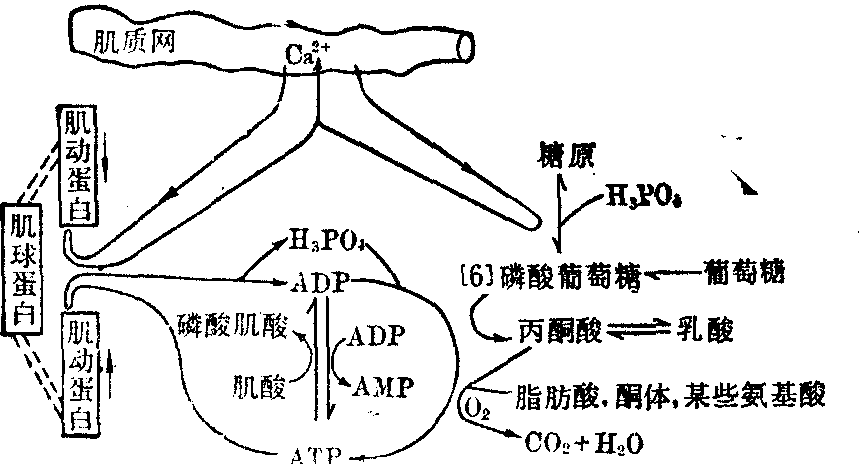
图1 肌肉收缩时的化学变化
AMP+H2O—→IMP+NH3IMP+门冬氨酸+GTP—→腺苷酰琥珀酸+GDP+Pi腺苷酰琥珀酸—→AMP+延胡索酸
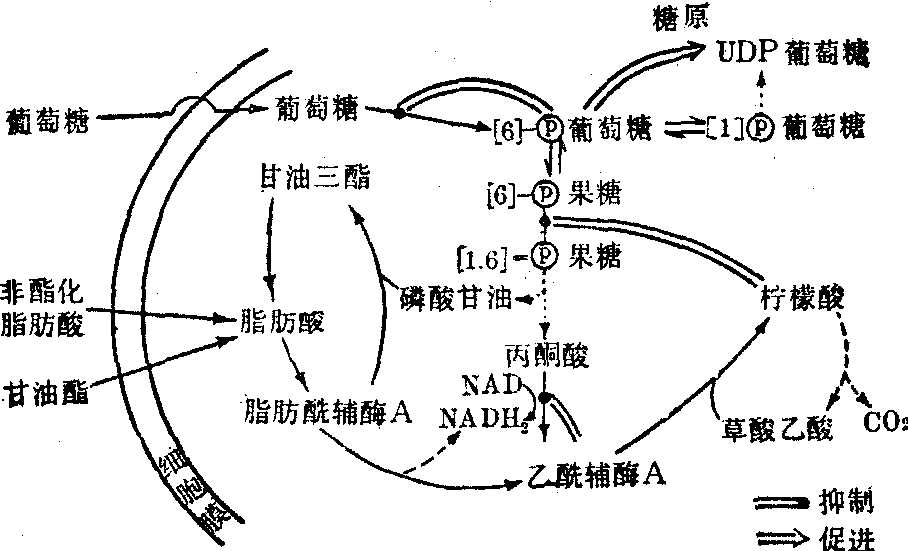
图2 肌肉组织中脂肪酸葡萄糖代谢之间的联系
经劳动或体育锻炼的肌肉,能代谢有显著改善。有训练之人的肌肉组织不但在活动时从血中摄取葡萄糖、脂肪酸等燃料的能力显著增强,而且肌细胞中糖原及甘油三酯的贮量亦明显增加。同时肌红蛋白的含量及线粒体的数目亦见增多; 参加糖及脂类有氧代谢及氧化磷酸化作用的酶活性升高(表)。综上可以认为,训练有素的肌肉组织在更大的程度上靠有氧代谢提供能量。比较有锻炼的人与无锻炼的人从事同等强度的肌肉活动时的代谢状况,以氧的消耗量代表有氧代谢进行的强度,以乳酸生成量代表无氧酵解进行的强度,结果发现有锻炼的人耗氧量较大,乳酸生成较迟亦较少。这充分说明锻炼确实使肌肉组织有氧代谢能力增强,因而使无氧酵解处于更为保留的地位,这是一种既有利于整个机体又有利于肌肉工作的适应; 亦即有锻炼的肌肉比较耐劳的原因。对于疲劳感目前还未能从化学上给以满意的解释。就目前所知,与疲劳感有关的化学因素包括:肌肉中供能物质(主要是肌糖原及磷酸肌酸)的耗竭; 终末产物 (主要是乳酸)的堆积; 组织中pH值的降低以及高能磷酸化合物的再合成能力不足或丧失等。所有这些因素可能是互相关连而不是彼此孤立的。如组织中的pH值降低不仅影响ATP的生成,而且可以活化ATP酶促进ATP的分解。甚者可使ATP空竭而引起肌肉僵直。
锻炼对骨胳肌生化性质的影响
| 单位 | 未经锻 炼的肌 肉 | 经过 锻炼的 肌肉 | |
| 糖 原 肌红蛋白 线粒体蛋白质 丙酮酸转换率 顺乌头酸酶 NAD异檬酸脱氢酶 琥珀酸脱氢酶 肉毒碱软脂酰转移酶 软脂酰-辅酶A脱氢酶 NADH脱氢酶 细胞色素a 细胞色素b 细胞色素c 细胞色素氧化酶(红肌) 氧吸收 | mg/g组织 mg/g组织 mg/g组织 μmol/(min·mg蛋白质) μmol/(min·g) μmol/(min·g) μmol/(min·g) μmol/(min·g) μmol/(min·g) μmol/(min·g) μmol/g组织 μmol/g组织 μmol/g组织 μlO2/(min·g) μ原子/(min·mg蛋白质) | 36.5 3.8 2.97 0.137 5.8 2.77 4.2 0.27 1.23 5.6 5.3 5.2 4.3 427 0.112 | 54.7 6.7 4.67 0.188 11.7 5.4 8.4 0.51 2.91 11.8 8.2 8.56 8.0 691 0 170 |
| 己糖磷酸激酶 | |||
| 红 肌 白 肌 肌球蛋白ATP酶 | U/mg蛋白质 U/mg蛋白质 μg无机磷/5min25℃ | 5.1 4.6 31 | 9.2 8.9 44.6 |
- 他人名号是什么意思
- 他人名字是什么意思
- 他人后代是什么意思
- 他人商店是什么意思
- 他人堂兄弟是什么意思
- 他人声音或文字是什么意思
- 他人夫妇是什么意思
- 他人女儿是什么意思
- 他人女婿是什么意思
- 他人如何欢,我意又何苦。是什么意思
- 他人妹妹是什么意思
- 他人妻兄是什么意思
- 他人妻子是什么意思
- 他人妻父是什么意思
- 他人姐姐是什么意思
- 他人姓名与官衔是什么意思
- 他人嫁衣是什么意思
- 他人子嗣是什么意思
- 他人子孙是什么意思
- 他人子孙的美称是什么意思
- 他人宅第是什么意思
- 他人宗族是什么意思
- 他人家属是什么意思
- 他人家族是什么意思
- 他人年龄是什么意思
- 他人幼子是什么意思
- 他人弓莫挽,他人马休骑是什么意思
- 他人情意是什么意思
- 他人所剩余的一点利益是什么意思
- 他人所教的学生是什么意思
- 他人指教或来信是什么意思
- 他人文字是什么意思
- 他人是地狱是什么意思
- 他人暗示是什么意思
- 他人有心,予忖度之是什么意思
- 他人有心,予忖度之.是什么意思
- 他人母亲是什么意思
- 他人演奏是什么意思
- 他人父亲是什么意思
- 他人父母是什么意思
- 他人牙慧是什么意思
- 他人生辰干支是什么意思
- 他人疾病是什么意思
- 他人的事迹是什么意思
- 他人的只言片语是什么意思
- 他人的妻妾是什么意思
- 他人给予的恩惠是什么意思
- 他人翰墨是什么意思
- 他人莫相笑,未易会吾曹。是什么意思
- 他人著作是什么意思
- 他人藏书是什么意思
- 他人言语是什么意思
- 他人言辞是什么意思
- 他人言辞·诗文是什么意思
- 他人言辞诗文是什么意思
- 他人评价法是什么意思
- 他人诗文是什么意思
- 他人诗文、信札是什么意思
- 他人诗文、赠礼是什么意思
- 他人诗文信札是什么意思