细胞的超微结构
在光学显微镜下,细胞的内部结构被粗略地分为细胞膜、细胞质和细胞核三部分,对细胞内部的细节观察受到了限制。应用电子显微镜(电镜)才能研究细胞的超微结构,在电镜下发现细胞主要是一些膜性结构,如线粒体、高尔基体、内质网、核被膜及细胞膜等。细胞靠这些膜性结构,将各种酶分子隔离开来,避免相互混杂,以保证细胞代谢作用的正常进行。细胞的这种由膜性细胞器,将功能相近的酶系分隔于一定区域内的作用,便是细胞内膜系的区域化作用。
细胞膜 细胞的最外层有薄膜包围着,这层薄膜即细胞膜又称质膜。细胞膜具有一定的结构,并执行着各种各样的功能。
细胞膜主要由蛋白质和类脂组成。细胞膜内尚含有少量的糖类,后者与蛋白质或类脂结合,构成糖蛋白、糖脂。细胞膜中蛋白质与类脂的比例,各约占一半。由于细胞超微结构研究的进展,细胞“膜”的概念也被充实和扩大。广义的细胞“膜”除指细胞表面的细胞膜之外,还包括细胞内的各种膜,如内质网膜、线粒体膜、核被膜等在内,并把细胞表面和细胞内部的膜,统称为生物膜。
细胞学家根据红细胞、神经纤维和卵细胞等的实验资料,提出了膜的分子结构理论。单位膜模型(图1)认为各种生物膜在电镜下都具有三层结构,两条电子密度高的暗带和中间的一条电子密度低的亮带。两条暗带宽各约2nm,代表蛋白质,亮带宽约3.5nm为类脂。膜的厚度为7.5nm。这个学说还提出,原核细胞和真核细胞的膜皆具有共同的单位膜结构。单位膜模型长时间以来被人们所广泛采用,但是它还存在着不足之处,似乎把膜的结构看成静止的,不认为它会随着功能的不同而发生变化。另外也难于解释非脂溶性物质的膜通透性问题。
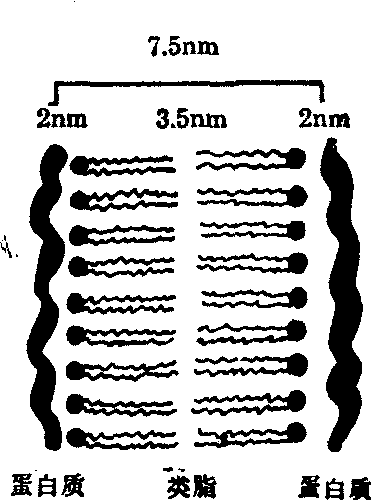
图1 单位膜模型图

图2 液态镶嵌模型图
目前较为公认的膜结构理论为液态镶嵌模型(图2),即在液态的类脂双分子层中,镶嵌着可以移动的球型蛋白质。蛋白质分为外在蛋白质和内在蛋白质两类。内在蛋白质镶嵌在类脂的双分子层中,其中有的埋在类脂的内层,有的埋在外层;外在蛋白质附着在类脂双层的内层表面,常与内在蛋白质的内侧表面以离子键相连。内在蛋白质的位置改变,可以影响外在蛋白质的功能,而外在蛋白质的收缩也可影响内在蛋白质的分布。在一般情况下,膜的类脂大多数处于液晶状态,这就使膜具有较大的流动性。实验证明,类脂和蛋白质在膜的流动结构中,能作一定程度的自由运动,如蛋白质分子在膜中可做侧向运动,内层的类脂分子可以移向外层等。生物膜的流动性使膜中的蛋白质能够适应功能的变化而改变其分布与结构,从而保证了细胞完成各种不同的生理功能。现已发现细胞的吞饮与吞噬作用,细胞分裂与细胞融合以及神经突触释放递质的作用等,皆与膜的流动性有关。
细胞质 细胞质中除去特化的部分——各种各样细胞器(核糖体、内质网、高尔基体、线粒体和溶酶体等)之外,剩下的是胶态物质,即细胞质基质。基质中含有若干种酶(糖酵解酶系,与氨基酸合成和分解有关的酶系等),中间代谢产物,以及储存的脂质、糖原等。细胞质基质也在不断地进行新陈代谢活动。
核糖体 核糖体又称核蛋白体,在电镜下为15—25nm的致密小颗粒,是由核糖体核糖核酸(rRNA) 与蛋白质结合而成的,其中rRNA约占60%,蛋白质约占40%。现已知道核糖体是由大小两个亚单位构成的。哺乳动物细胞核糖体的大小两个亚单位的沉降系数分别为60S和40S。核糖体可以附着在内质网的表面上,也可游离于细胞的基质之中。不论是附着的或是游离的核糖体,皆可由信使核糖核酸(mRNA) 细丝串联成为多聚核糖体或称多聚体(图3、4)。多聚核糖体在电镜下呈花簇状或
螺旋状的集合体,一般由3—30个核糖体组成。核糖体是细胞内蛋白质合成的基地,也可譬喻为“装配”蛋白质的机器。附着于内质网的核糖体主要合成外输蛋白,而游离核糖体主要合成供细胞本身生长所需要的结构蛋白。

图3 多聚核糖体
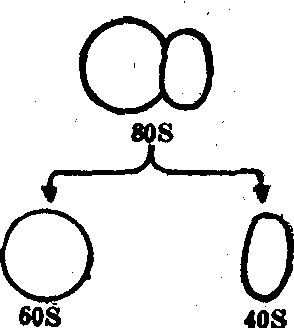
图4 核糖体组成成分示意图
内质网 内质网是细胞质中的膜性管道系统,普遍存在于一般细胞之中,但其数量各有不同。根据其表面有无核糖体的附着,可分为两种类型,粗面内质网 (rER)和滑面内质网(sER)。粗面内质网的主要特征是表面附着核糖体。粗面内质网常见于蛋白质合成旺盛的细胞之中,多为扁平囊状,彼此平行地密集在一起。粗面内质网的主要功能是合成外输蛋白(或称分泌蛋白),也合成少量细胞自身所需的结构蛋白 (结构蛋白主要由游离的核糖体合成)(图5)。核糖体是蛋白质合成的场所,膜性管道则是新合成蛋白的运输通路(图6)。

图5 粗面内质网(膜上小颗粒为核糖体)
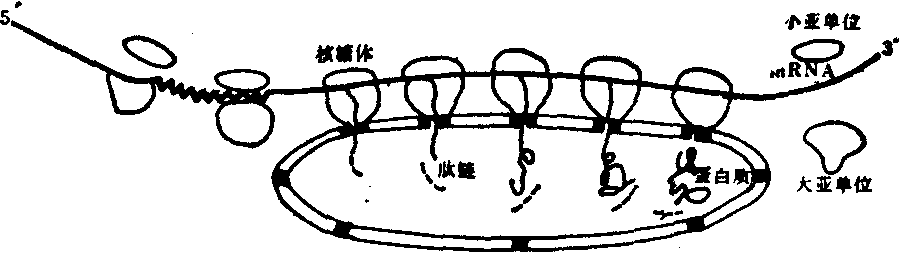
图6 粗面内质网的蛋白质合成示意图
滑面内质网一般都呈小管状,或小泡状,表面光滑,没有核糖体附着。不同细胞内的滑面内质网,具有不同的功能,如具有解毒,运输脂类,传导兴奋,分泌盐酸等作用。两型内质网在各种细胞的分布是不一样的。有的细胞粗面内质网很发达,如胰腺细胞、浆细胞等; 有的细胞富于滑面内质网,如平滑肌细胞、胃腺的壁细胞等; 有的细胞两者兼而有之,如肝细胞。内质网的出现,一方面把细胞质基质分隔成若干不同区域,使细胞内物质代谢能够在特定的环境下进行,另一方面也扩大了细胞内代谢活动的“表面”,而使各种反应得以高效率地进行。
高尔基体 高尔基体又称高尔基器,位于细胞核附近,其形态呈网状,所以也叫内网器。在电镜下高尔基体包括扁平囊泡、小泡和大泡三种成分。扁平囊泡又叫高尔基囊泡,由3—8个扁平囊泡平行排列在一起,形成高尔基体的主体。扁平囊泡一般略弯曲而呈弓形,其凸面为生成面,凹面称成熟面或称分泌面。在有极性的细胞中,形成面朝向细胞基底部,成熟面则面向细胞表面。成熟面的膜上附有酶,能够促使分泌物的浓缩和成熟。小泡常见于形成面和扁平囊泡周围,数目较多。一般认为小泡是由粗面内质网出芽生成的,其中充满蛋白质分泌物,并不断地向扁平囊泡并入。大泡一般为数个,常位于扁平囊泡的成熟面,是扁平囊泡末端或成熟面的膨大部分。大泡带着由扁平囊泡生成的分泌物,形成分泌囊泡离开高尔基体(图7)。扁平囊泡由于小泡的不断并入和大泡的经常脱离,而处于动态的变化中。高尔基体参与细胞分泌物的储存和加工活动。具有分泌作用的细胞中,高尔基体皆较发达。粗面内质网合成的蛋白质(或称分泌蛋白)在高尔基体内储存,经过脱水缩合,加入糖类物质之后,再被转运到细胞之外。
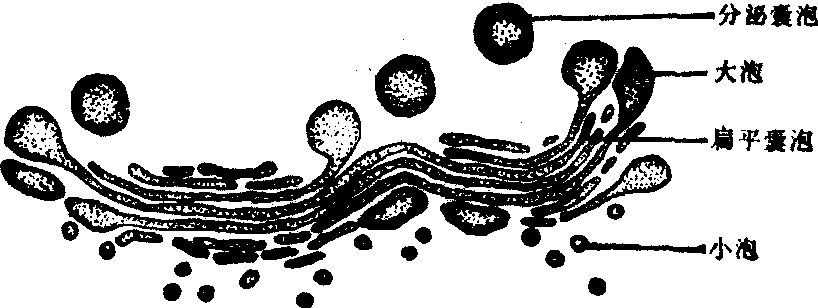
图7 高尔基体
线粒体 在电镜下,线粒体是由内外两层膜(即两层单位膜) 包围而成的封闭结构。内膜向线粒体内部突伸形成嵴,用以扩大内膜的表面积。嵴一般为板层状,常与线粒体的长轴垂直。围于线粒体内膜的腔,称为内腔,其中充满基质。内膜朝向基质的面上尚有许多内膜亚单位又称基粒,由头部、柄部和基片组成(图8)。基质中除含有蛋白质(包括酶类)及某些类脂成分外,还含有DNA,RNA及核糖体。线粒体可以合成少量构成自身的结构蛋白。在基质中尚可看到一种明显的致密结构,称为基质颗粒,为Mg2+、Ca2+等离子磷酸盐,它们与调节线粒体内部的离子环境有关。线粒体是细胞供能的中心,细胞代谢所必需的总能量中,大约95%来自线粒体。三羧酸循环、电子传递、磷酸化、脂肪酸氧化和氨基酸氧化等过程均在此处进行。参加上述化学反应的各种酶系,在位置和空间上具有严格的排列顺序。线粒体的外膜上分布有单胺氧化酶以及与磷脂代谢有关的一些酶。内膜上结合有电子传递链及氧化磷酸化的酶系。基质中含有与三羧酸循环和脂肪酸氧化有关的酶系。
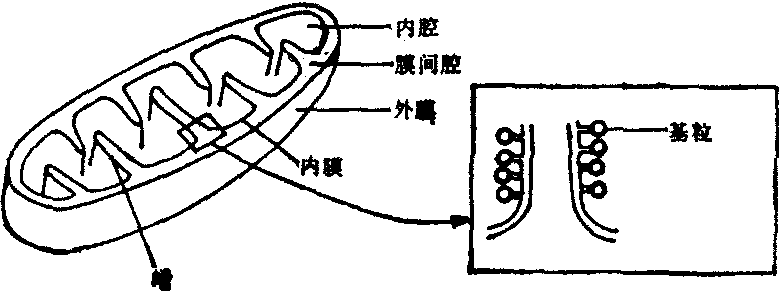
图8 线粒体构造
溶酶体 溶酶体是一些由厚约6nm的单位膜包围的小颗粒,内含40多种水解酶,对蛋白质、糖类、脂肪、核酸和磷酸等物质都能分解,所以被视为细胞内的消化器。溶酶体可分为两类,即初级溶酶体和次级溶酶体。初级溶酶体是刚形成的,仅含水解酶而没有作用底物的溶酶体。初级溶酶体与吞噬泡或自噬泡合并,便形成次级溶酶体。次级溶酶体不仅含有水解酶,而且还有作用底物。溶酶体作用的底物可以是细胞内退变或崩解的某些细胞器,也可以是由细胞的吞噬或胞饮作用而摄入的外源物质。溶酶体对细胞自身受损或衰退的细胞器,进行的消化分解,称自溶作用,对被吞噬或胞饮的异物进行的消化分解,称异溶作用(图9)。电镜下常见的次级溶酶体有多泡体、脂褐质和髓鞘样结构等。多泡体可能是由移行于细胞内的胞饮小泡群聚而成,其作用底物可以是内源性的,也可以是外源性的。在一些内分泌细胞中,多泡体可以消化分泌颗粒,并借以调节分泌过程。脂褐质为成群的不规则小体,电子密度较高,常含中等密度的透明脂滴,其作用底物有脂类、糖原和细胞器等。髓鞘样结构的内容物为膜性成分,常呈指纹排列,可能是溶酶体内一些未能分解的脂类物质水化后而形成的。髓鞘样结构可见于正常细胞中,但在病理情况下会增多。
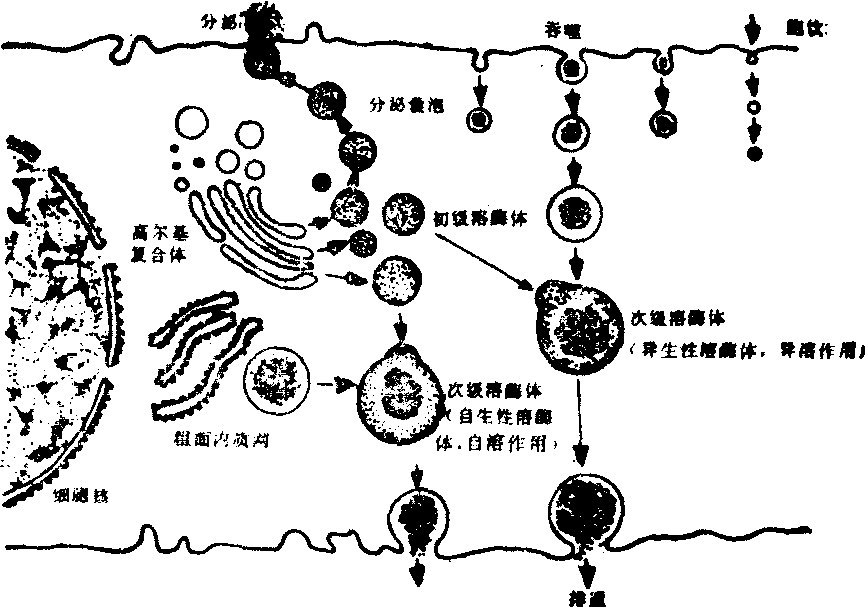
图9 细胞内溶酶体的形成与转化
过氧化物酶体 过氧化物酶体或称过氧化氢体,是由单位膜围成的一种特殊细胞器,其体积比溶酶体小,直径约为0.3—0.5 μm,呈圆形或卵圆形。过氧化物酶体里既含有催化生成H2O2的氧化酶,又含有分解H2O2的过氧化氢酶。H2O2对细胞有毒害作用,过氧化物酶体中的过氧化氢酶可分解过多的H2O2,以控制H2O2的含量。这种细胞器常见于动物的肝细胞、小肠细胞和肾脏近曲小管细胞内。有人认为它是溶酶体的一种,但其结构及成分与溶酶体确有不同之处,如鼠类肝细胞的过氧化物酶体内存在着晶体状或板状结构。
细胞骨架 细胞骨架为细胞内的复杂网架系统,由微丝、微管和中等纤维组成,与细胞的形状和运动有密切关系。
(1) 微丝: 微丝又称细胞质丝,为直径5—15nm的细丝广泛地存在于各种细胞之中。微丝是一种实心性结构,主要由一些伸展性的蛋白质分子集合而成。细胞内的细丝可根据其功能分为以下几种。细胞支架细丝具有支撑作用,存在于微绒毛的轴和软骨细胞中。肌细丝主要存在于肌细胞中,具有收缩功能。存在于神经细胞的树突和轴突中的神经细丝除具有支架作用外,可能还与神经细胞内代谢物质的运输有关。
(2) 微管: 微管普遍存在于真核细胞中,为不分支的中空小管,长短不定,直径为18—25nm,管壁厚度为4—5nm。微管的管壁由十三根细丝围成,细丝是由微管蛋白组成。在不同的细胞内微管所起的作用各不相同。神经细胞突起内的微管呈平行状排列,可能具有支持以及运输神经递质的功能。纤毛、鞭毛和中心粒的微管融合成双体和三合体,主要起着运动的作用。除纤毛和鞭毛的微管之外,其他微管结构都是不稳定的。它们可以随着细胞功能的变化,不断解聚为微管蛋白,同时另一些微管蛋白又可合成新的微管。
(3) 中等纤维: 中等纤维是一类结构相似,而化学成分不同,直径约为10μm左右,介于微丝和微管之间的细胞质内纤维,故称中等纤维。有人认为,它们是由四个直径为2—3nm的亚单位所组成。中等纤维具有种属与组织的特异性,根据这一特点可用来进行肿瘤细胞来源的鉴别诊断。目前至少已发现五种不同生化组成的中等纤维。如角质蛋白纤维、结蛋白纤维、波形蛋白纤维、胶质蛋白纤维和神经蛋白纤维等。
中心粒 一般情况下,每个细胞具有一对中心粒,位于细胞核附近。中心粒具有自身复制能力,在细胞有丝分裂时期,中心粒则变为两对。在肿瘤细胞和巨核细胞中,中心粒的数目可成倍地增多。电镜下,中心粒为短小的筒状结构,直径0.1—0.5μm,长0.3—0.7μm。其筒壁由九组微管构成,每组微管又由三个微管组合而成,这三个微管也称为三合体,每个微管均由十三根细丝组成。成对的中心粒通常互相垂直排列,在横切面上,可见九组微管,一组紧挨一组地斜向排列成环状,略似风车的旋翼(图10)。目前,对中心粒的生理功能尚不十分了解,但认为它对细胞有丝分裂时期中染色体的移动方向和纺锤丝的排列方向有密切关系。

图10 中心粒立体结构示意图
包含物 通常包含物是指积聚在细胞内并具有一定形态的代谢产物。过去大多数人认为包含物只是细胞内暂时储存的物质,对细胞不一定是必需的。但是,现在却认为包含物广泛参与细胞的代谢及功能活动。近年来应用电镜对细胞进行的超微结构研究,发现包含物与细胞器之间似乎没有截然的界限。如神经细胞的脂褐
 ,实际上是细胞内的次级溶酶体。细胞内常见的包含物,有糖原、脂类及蛋白质等。
,实际上是细胞内的次级溶酶体。细胞内常见的包含物,有糖原、脂类及蛋白质等。(1) 糖原: 糖原为动物细胞内的碳水化合物存在形式。主要存在于细胞质内,有时也可在细胞核、溶酶体或线粒体中观察到。糖原颗粒可分为两种类型,α颗粒和β颗粒。α颗粒的直径为80—100nm,聚集成花簇状,如肝细胞内的糖原。β颗粒的直径则为15—30nm,分散存在于细胞质内。这两种颗粒的生物意义尚不清楚。细胞内的糖原数量可由于不同生理及病理情况而发生变化。如肝细胞内的糖原,饱食时增加,禁食时减少。
(2) 脂类:脂类以脂滴的形式存在于细胞质内,也可出现于细胞核、内质网、高尔基体或线粒体中。电镜下呈大小不等的泡状结构,内含物为均质性,但致密度深浅不一。脂类在脂肪细胞和肾上腺皮质细胞中特别多,在一般细胞中较少。脂类可作为细胞的能量来源,构成膜或形成富于脂类结构的成分。
(3) 蛋白质: 蛋白质存在于分泌颗粒和初级溶酶体中,也可呈结晶状存在于细胞质中。有时也可在线粒体和内质网中发现结晶状蛋白质。
细胞核 细胞核是细胞内的一个重要构造。细胞核包含着遗传物质,蕴藏着遗传信息,决定细胞内蛋白质的生物合成,并在很大程度上控制细胞的生命活动过程。失掉核的细胞,由于失去其固有的生活功能,便很快趋于死亡。哺乳动物的红细胞成熟后由于细胞核消失,不能合成蛋白质,细胞不再分裂,因而寿命是有限的。
多数细胞只有一个细胞核,有的细胞具有两个核,也有的多达数百个,如骨胳肌细胞。有的则没有核,如哺乳动物的红细胞。一般情况下,细胞核与细胞质体积之间有一定的比例关系。细胞体积增大时,细胞核也变大。幼稚细胞的细胞核相对较大,而衰老细胞的则较小。细胞核的形态与细胞的形态相关。在球形、立方形和多角形的细胞中,细胞核一般为球形; 在柱形细胞中常为椭圆形;在梭形细胞中为杆状等。但也有不少例外,如中性粒细胞核分成2—3叶,而呈不规则状。处于分裂间期的细胞核,可分为核被膜、核质、核仁和染色质等部分(图11)。
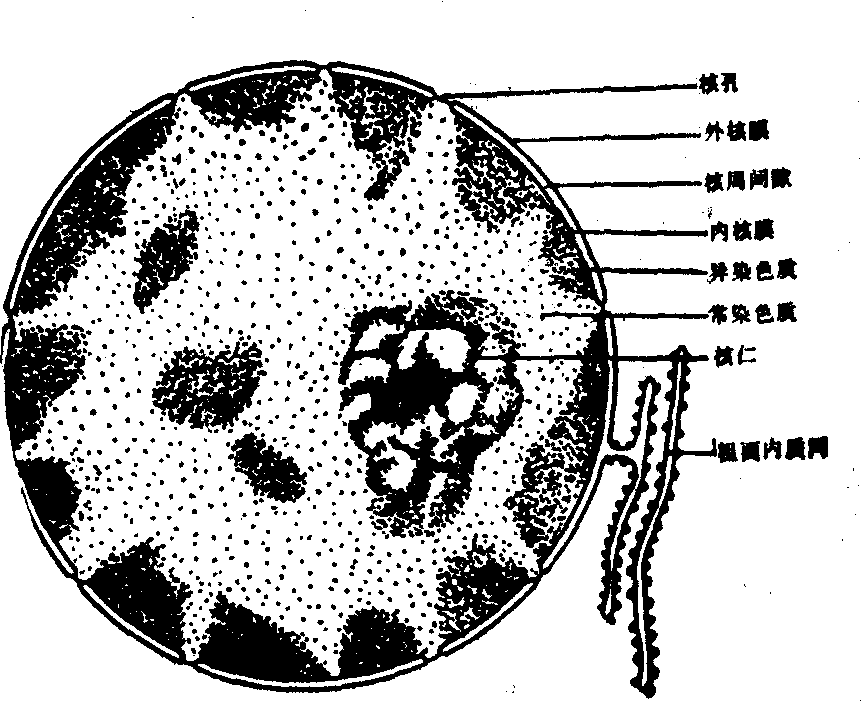
图11 细胞核结构示意图
核被膜 光镜下核被膜为包围细胞核的一层薄膜。但应用电镜观察,发现核被膜构造比较复杂,由内、外核膜、核周间隙和核孔等构成。内、外核膜大致是平行的,每层膜的结构类同一般细胞膜,厚约7—8nm。常可见到外核膜与内质网膜相连,核周间隙与内质网腔相通的情况。外核膜上附有核糖体及多聚核糖体,一般认为在核被膜也能合成蛋白质。核周间隙为两层核膜之间的腔隙,宽约15—30nm。核被膜上散在有许多小孔,称为核孔,是由内、外两层核被膜融合而成的。孔径变化较大,一般为70nm左右。核孔约占核表面积的5—15%。正常情况下,每一平方微米核被膜上约有35—65个核孔。核孔的多少与细胞的功能状态有关,代谢旺盛的细胞,其核孔较多。核内大部分物质可通过核孔移到核外。核被膜对调节细胞核与细胞质间的物质出入、维持核内环境的恒定起着重要作用。
核质 核质是被核被膜所包围的无定形基质。其中含有水、各种无机盐和一些可能在分布上具有区域性、功能上带有专一性的酶类等。
核仁 核仁一般为圆形,常靠近细胞核的一侧。一般细胞有1—2个核仁,也有的细胞没有核仁,如中性粒细胞。在代谢旺盛、生长迅速的细胞中核仁较大。核仁通常由原纤维成分、颗粒成分、核仁相随染色质和核仁基质等部分组成。原纤维成分为紧密排列的原纤维丝,粗约5—10nm,构成核仁的海绵状网架。颗粒成分为圆形致密颗粒,直径约15—20nm,散布于网架之间,或分布于原纤维周围。电镜下观察到的核仁丝是由上述两种成分构成的。核仁相随染色质可分两部分: 一部分围绕着核仁,也叫核仁周围染色质,主要为异染色质; 另一部分则深入核仁内,叫做核仁内染色质,主要为常染色质。核仁基质与核质相同,核仁的有形成分埋藏于核仁基质之中。上述各种成分的比例关系不是固定不变的,而是随着细胞功能状态而不断地发生变化。
核仁丝与颗粒成分含有RNA,核仁染色质则含有DNA。应用电镜,可以看到核仁染色质中的DNA分子转录rRNA的情景。新形成rRNA与蛋白质结合,形成核糖体的前体之后,经核孔进入细胞质内,再形成核糖体(在细胞质内多个核糖体可由mRNA加以串联形成多聚核糖体,多聚核糖体便是细胞内蛋白质合成的场所)。核仁由于能够合成核糖体的前体而与蛋白质合成密切相关。所以,在一些蛋白质合成旺盛的细胞,如消化腺上皮细胞和一些快速生长和分裂的细胞,如肿瘤细胞、再生细胞等,核仁比较大。
染色质 染色质主要由DNA、组蛋白与非组蛋白构成,为嗜碱性物质,易被碱性染料染色,故而得名。染色质为丝状结构,也叫染色质丝,在间期核内呈分散状态,不易见到。但当细胞分裂时,染色质便高度螺旋化,形成明显可见的染色体。由此可见,染色质和染色体实际上是同一物质,只是在不同条件下,显示出不同的形态结构。
在电镜下,可观察到染色质表现为两种形态结构,一为异染色质,另一为常染色质。异染色质电子密度较高,通常分布于核被膜的内核膜内侧,厚薄不一,同时也以大小不等的斑块状分散于核质中,另外有些异染色质围绕核仁形成一层外壳。异染色质为螺旋化程度高,功能不活泼的染色质。一般认为当异染色质中的DNA与组蛋白结合时DNA的活性被抑制,而不再转录RNA。常染色质位于异染色质之间的浅亮区域,也有一部分分散于核仁染色质中。异染色质与常染色质相间排列时,可使细胞核呈现车轮状,如浆细胞的细胞核,常染色质为螺旋松解、功能活泼的染色质,一些人认为常染色质中的非组蛋白如果与组蛋白发生竞争性结合,或者与DNA直接结合皆可使DNA显示活性,而进行RNA的转录。一般状态下,分化程度高的细胞,如精子、低等脊椎动物的红细胞等,其异染色质所占比例较大;而迅速分裂的细胞,如胚胎细胞、肿瘤细胞等则常染色质的含量较多。
染色质的DNA是主要的遗传物质基础,DNA中蕴藏着遗传信息(即DNA多核苷酸链上各种碱基的特定排列顺序)。DNA通过碱基配对原则决定mRNA的多核苷酸排列的方式,而将其遗传信息传递给mRNA,称为转录。由DNA转录的mRNA经核孔进入细胞质,在核糖体上把RNA搬运的氨基酸,按严格顺序排列起来,称为翻译,组成蛋白质。蛋白质则是遗传性状的具体表达者。虽然从细胞的性状传递,蛋白质的合成来看,细胞核起着重要作用,但细胞核并不能脱离开细胞质而长期存在。从实际情况看,细胞核控制细胞质的合成活性,细胞质又能影响细胞核的功能。细胞的各种生命活动是在细胞核和细胞质相互协调、共同作用下完成的。
细胞的超微结构主要是由膜、微管、细丝和微粒等构成的,它们的化学成分为蛋白质、核酸、脂类和糖类等生物大分子。细胞内的各种超微结构不是固定不变的,而是随着细胞的生命活动过程不断的发生调整和改变。例如,构成细胞内各种结构的膜,在一定的条件下,可以相互移行和补充,细胞膜可以参与内质网的形成,内质网又可以构成高尔基体,从高尔基体成熟面脱下来的分泌颗粒,由细胞排出后,其界限膜又与细胞融合而构成细胞膜。
- 匀称是什么意思
- 匀称型是什么意思
- 匀称型发育迟缓是什么意思
- 匀称整齐是什么意思
- 匀称的体形是什么意思
- 匀空儿是什么意思
- 匀红点翠是什么意思
- 匀细是什么意思
- 匀胶铬版是什么意思
- 匀脂抹粉是什么意思
- 匀脸是什么意思
- 匀苗是什么意思
- 匀调是什么意思
- 匀贴是什么意思
- 匀身是什么意思
- 匀辫是什么意思
- 匀适是什么意思
- 匀透是什么意思
- 匀速圆周运动是什么意思
- 匀速直线运动是什么意思
- 匀速跑是什么意思
- 匀速运动是什么意思
- 匀静儿是什么意思
- 匀顶是什么意思
- 匀𬘿是什么意思
- 匁是什么意思
- 匂是什么意思
- 匃是什么意思
- 匄是什么意思
- 匄注山房是什么意思
- 匄者贱儒是什么意思
- 包是什么意思
- 包
 彭是什么意思
彭是什么意思 - 包、保、核经济责任制是什么意思
- 包㞘儿是什么意思
- 包一民是什么意思
- 包一虚是什么意思
- 包万荆贺是什么意思
- 包三鏸是什么意思
- 包上是什么意思
- 包不了符是什么意思
- 包不住是什么意思
- 包不得是什么意思
- 包世臣是什么意思
- 包世臣与小倦游阁是什么意思
- 包世臣农业经济思想是什么意思
- 包世臣楷书录语轴是什么意思
- 包世臣的消费观是什么意思
- 包世臣的货币思想是什么意思
- 包世臣草书临书谱轴是什么意思
- 包世臣草书对联是什么意思
- 包世臣行书节临秋深帖是什么意思
- 包世臣行草摘录书谱屏是什么意思
- 包世荣是什么意思
- 包丞相出身全传宝卷一卷是什么意思
- 包严儿是什么意思
- 包中庆是什么意思
- 包中青是什么意思
- 包举是什么意思
- 包举四海是什么意思