糖脂glycolipid
含糖的类脂。由一个或几个半乳糖或其他糖通过糖苷键与二酰甘油或酰化的鞘氨醇连接而成的复合脂。根据醇的不同,分为两类:
❶甘油醇糖脂。由脂肪酸、甘油、半乳糖或其他糖组成,如单半乳糖甘油二酯、双半乳糖甘油二酯。
❷鞘氨醇糖脂。由脂肪酸、鞘氨醇、半乳糖或其他糖组成,如
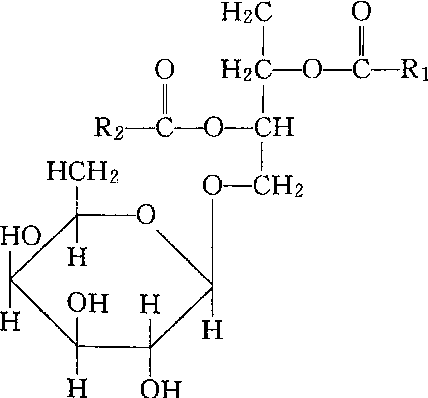
单半乳糖甘油二酯
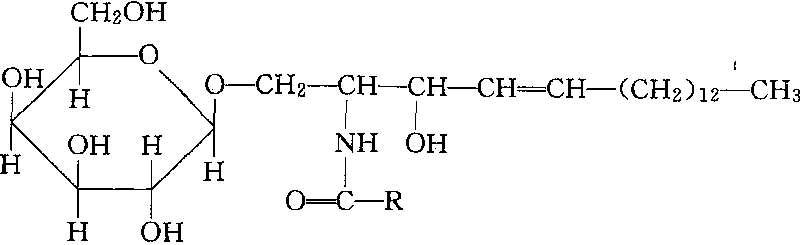
脑苷脂
脑苷脂、神经节苷脂。两类中的代表化合物的结构如上。分子中的脂肪酸和鞘氨醇链是疏水尾,糖基为亲水头,与磷脂结构类似,参与生物膜的组成。在植物的叶绿体及动物的脑组织和神经节细胞膜系统中含量较多。糖脂glycolipide
糖脂质总称为糖脂。含有半乳糖或葡萄糖、长链脂肪酸、神经氨基醇等,但不含磷酸。糖质是神经髓鞘的主要成分,亦是组成其他组织的膜结构和血型物质等。
糖脂
含有糖分子的脂类物质。可由甘油衍生,也可由神经鞘氨醇衍生出来,常归于甘油酯(如糖基二酰基甘油酯)和神经鞘氨醇糖脂类(如脑苷脂)。是细胞膜结构组分,也是一些生物大分子(如脂蛋白和脂多糖)的组成成分。
糖脂glycolipid
指含有糖的脂类。
糖脂
糖脂
含糖而不含磷的脂类统称糖脂类。糖脂类又可以按其分子组成再分为甘油糖脂和鞘糖脂两类。含硫的糖脂习惯上称为硫脂,如精脂及硫酸脑苷脂等。
甘油糖脂 又称糖基甘油脂,是一类由甘油脂和单糖乃至寡糖所组成的糖脂,结构比较简单,一般都是糖基通过苷键与1,2-二脂酰甘油相结合的产物。含硫的糖脂则是硫酸基以硫酯键的形式结合在糖基上。
甘油糖脂广布于植物以及微生物界,但近年在动物界也不断有所发现。如脑组织中的半乳糖甘油脂,唾液及胃液中的葡萄糖基甘油脂等。精脂也是一种甘油糖脂,不仅分布于哺乳动物的精囊和精子,并随精子的成熟含量递增。
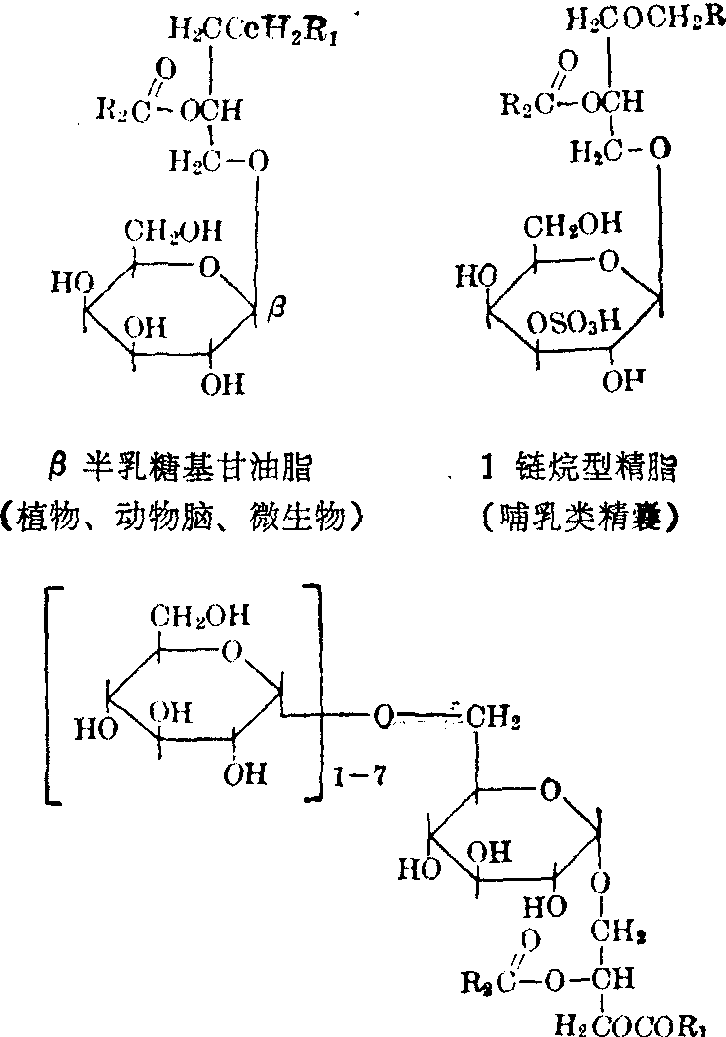
α聚葡萄糖基甘油脂(人唾液,人胃液)
鞘糖脂
鞘糖脂类的化学 鞘糖脂种类极多, 且结构复杂, 但其基本结构都是以N-酰鞘氨醇(又称神经酰胺)或具有与N-酰鞘氨醇结构相近似的长链碱基为基础,再通过其1位羟基与糖基所构成的β-糖苷衍生物,它们的化学通式可写为:
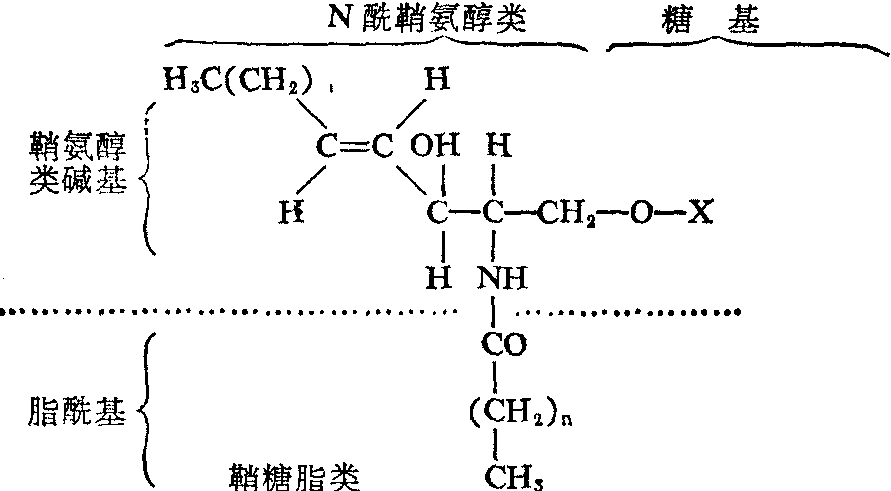
从上列通式不难看出,鞘氨醇类碱基及酯酰基具有疏水性,而糖基呈亲水性,因此在整个鞘糖脂分子内部形成疏水和亲水两个部分。这与鞘糖脂类的细胞分布和生物学作用有密切关系。
(1) 鞘氨醇类及N-酰鞘氨醇类:存在于自然界中的鞘氨醇类,其碳链长度固有不同,但是绝大多数由十八碳组成,分子中除含氨基、伯醇基各一个外,最少还必须有一个仲醇基。 由于长链上的第2、 第3位碳原子不对称,理应存在四种空间异构物,可是目前在动物组织中所发现的全属D—(+)—赤藓型。又在4、5位碳间常具有双键,而天然存在的一般总是反型的。分布于动物体的鞘氨醇类主要是鞘氨醇,植物的主要是二氢鞘氨醇、植物鞘氨醇以及8-脱氢-4-羟二氢鞘氨醇等。但是近年在动物肾脏也发现有少量植物鞘氨醇存在。几种常见鞘氨醇类的化学结构如下:
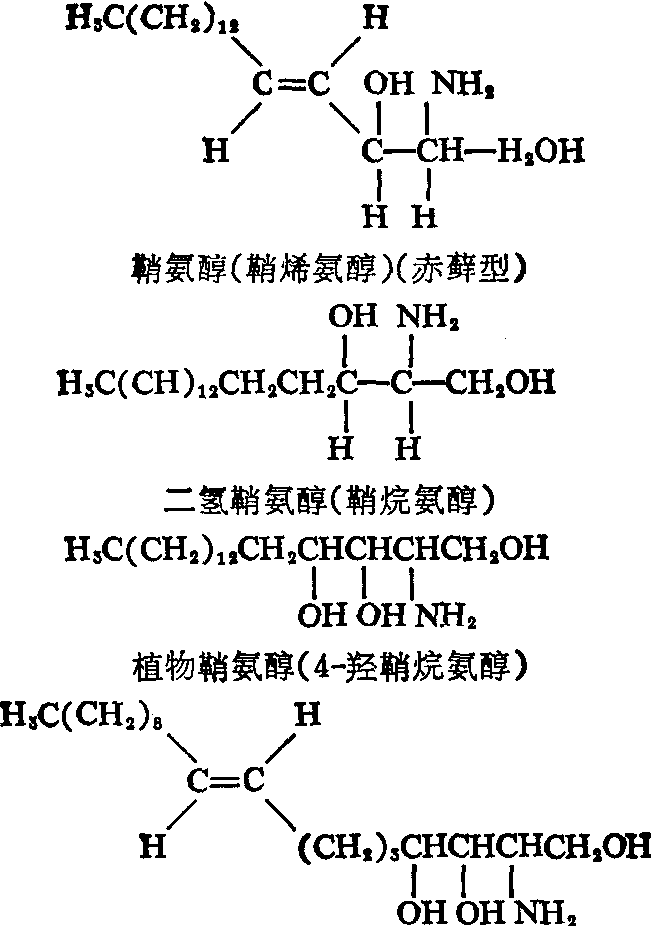
8-脱氢植物鞘氨醇(8-脱氢-4-羟鞘烷氨醇)
鞘氨醇类借其氨基与脂酰基相结合的酰胺化合物称N-酰鞘氨醇(神经酰胺)类。鞘糖脂类和鞘酯类化合物全可以看做是N-酰鞘氨醇类的衍生物。
存在于动物组织的N-酰鞘氨类分子中的脂酰基通常是由十六及十八碳组成,而最常见的又是硬脂酰基。但是人类脑中鞘糖脂常含有较多的廿四碳酰基,故曾以其所含廿四碳酰基来命名鞘糖脂(表1)。然而随着分子中糖基数的增加,糖脂的亲水性逐渐增大,在一些含寡糖基的糖脂中,即使酰基有一定差别,也不会影响整个分子的性质。
表1 脑中鞘糖脂的名称及其酰基组成
表2 部分糖基N酰鞘氨醇和硫脂的名称及结构
| 分 类 | 名 称 | 缩写记号 | 结 构 |
| 单糖基N酰鞘氨醇 | 葡萄糖基N酰鞘氨醇 (葡萄糖基脑苷脂) | GL-1a | Glc(β1→1′)Cer |
| 半乳糖基N酰鞘氨醇(脑苷脂)** 硫酸半乳糖基N酰鞘氨醇** | GL-1b GL-1bS | Gal(β1→1′)Cer HO3S3Gal(β1→1′)Cer | |
| 二糖基N酰鞘氨醇 | 乳糖基N酰鞘氨醇(胞糖苷H) 双半乳糖基N酰鞘氨醇** 硫酸乳糖基N酰鞘氨醇 | GL-2a GL-2b GL-2aS | Gal(β1→4)Glc(β1→1′)Cer Gal(α1→4)Gal(β1→1′)Cer HO3S3Gal(β1→4)Glc(β1→1′)Cer |
| 三糖基N酰鞘氨醇 | Fabry己三糖基N酰鞘氨醇 (pK抗元) | GL-3a | Gal(α1→4)Gal(β1→4)Glc(β1→1′)Cer |
| 氨基己三糖基N酰鞘氨醇 | GL-31c | GlcNAc(β1→3)Gal(β1→4)Glc(β1→1′)Cer | |
| Tay-Sach红细胞糖苷脂 | GL3g (即无唾液酸GM2) | GalNAc(β1→4)Gal(β1→4)Glc(β1→1′)Cer | |
| 四糖基N酰鞘氨醇 | 红细胞糖苷脂Ⅰ (P抗原、胞糖苷脂K) | GL-4a | GalNAc(β1→3)Gal(α→4)Gal(β1→4)Glc(β1→1′)Cer |
| 胞糖苷脂R | GL-4a(iso) | GalNAc(β1→3)Gal(α1→3)Gal(β1→4)Glc(β1→1′)Cer | |
| 己四糖基神经节N酰鞘氨醇 | GL-4g (即无唾液酸GM1) | Gal(β1→3)GalNAc(β1→4)Gal(β1→4)Glc(β1→1′)Cer | |
| 己四糖基N酰鞘氨醇 | GL-41c | Gal(β1→3)GalNAc(β→3)Gal(β1→4)Glc(β1→1′)Cer |
* Cer: N酰鞘氨醇基; Glc:葡萄糖基;Gal:半乳糖基; GlcNAc: N酰氨基葡萄糖基; GalNAc:N-酰氨基半乳糖基;
α或β糖苷的空间配位; 1′,1,2,3,4: 结合位置。例如HO3S3Gal(β1→4)Glc (β1→1′)Cer表示
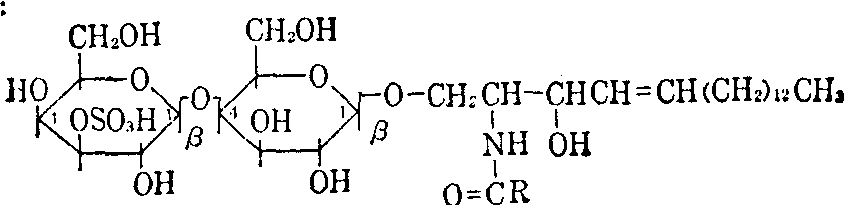 **均具有半乳糖与N酰鞘氨醇直接相连结的结构特点,故有人把它们统称为半乳糖脂
**均具有半乳糖与N酰鞘氨醇直接相连结的结构特点,故有人把它们统称为半乳糖脂(2) 糖: 鞘糖脂分子中的糖基可以是单糖、双糖、叁糖,直至寡糖,而且组成这些糖基的单位主要是己糖和N-乙酰氨基己糖。海鸟类的木糖脂虽含有木糖,但在其他动物尚无发现。酸性糖脂的神经节苷脂分子中还含有酸性较强的N-乙酰神经氨酸(又称唾液酸)或N-羟乙酰神经氨酸,前者存在于一切动物,后者仅发现于人和鸡以外的动物。单糖的结构见“糖类”条。
(3) 鞘糖脂类的糖链结构与分类: 随着新的鞘糖脂的不断发现,给鞘糖脂类的命名和分类带来很多困难。目前比较普遍应用的一种分类是先按分子组成,将鞘糖脂分为三组:
❶只含己糖基及N-乙酰氨基己糖基的鞘糖脂称为中性鞘糖脂;
❷含N-乙酰神经氨酸基或N-羟乙酰神经氨酸基的鞘糖脂称酸性鞘糖脂或神经节苷脂;
❸带硫酸基的称硫脂。中性鞘糖脂还可根据糖基中单位糖基数的多少,再分成单糖基N-酰鞘氨醇(GL-1),二糖基N-酰鞘氨醇(GL-2),三糖基鞘氨醇(GL-3),四糖基N-酰鞘氨醇(GL-4)等。神经节苷脂按糖基中N-乙酰神经氨酸或N-羟乙酰神经氨酸基的数目又可以分为GM、GD、GT、GQ等, 并且每种还可以根据其薄层层析的Rf值的不同再分为几个亚属、如GM1、GM2、GM3、GM4…,(GM1的Rf值最小; Rf值的大小与糖基的长度成反比关系)。现将部分鞘糖类按上述分类原则列表如下(表2,3,4):
鞘糖脂类的合成代谢 参与鞘糖脂合成的酶类主要是膜结合性的,不易分离纯化,因此仍有许多不够清楚的地方。一般认为合成鞘糖脂的直接原料是N-酰鞘氨醇类(作为糖基受体)和糖基单核苷酸(作为糖基供体)。N-酰鞘氨醇类的合成酶系统主要存在于微粒体,而糖基的转移反应则主要依靠高尔基复合体中的多种糖基转移酶类的催化。
表3 部分血型糖脂*的结构
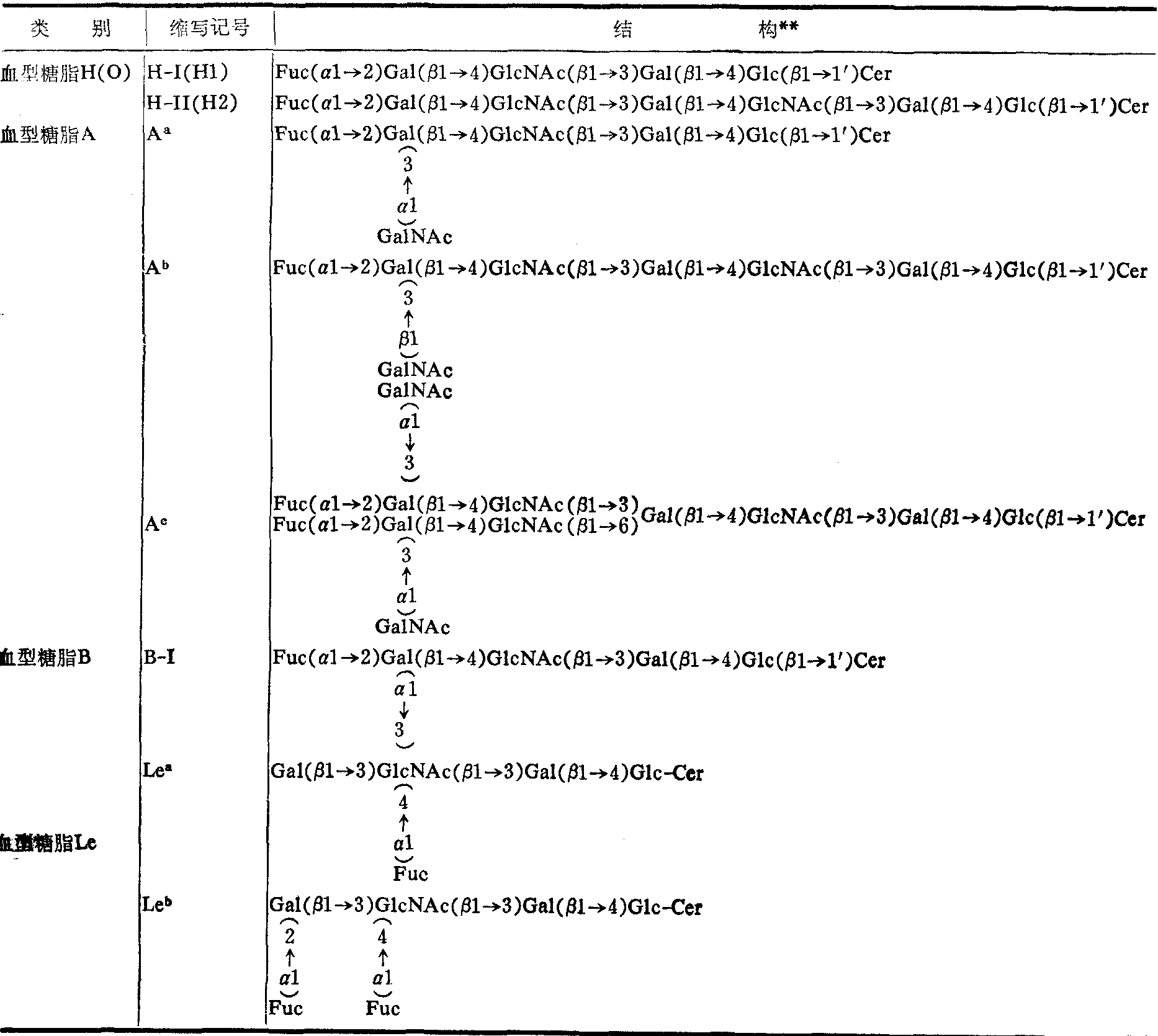
*组成上均含GlcNAc,且具特殊生物学活性,故另列表
**Fuc:岩藻糖基,其它与上表相同
表4 部分神经节苷脂的结构
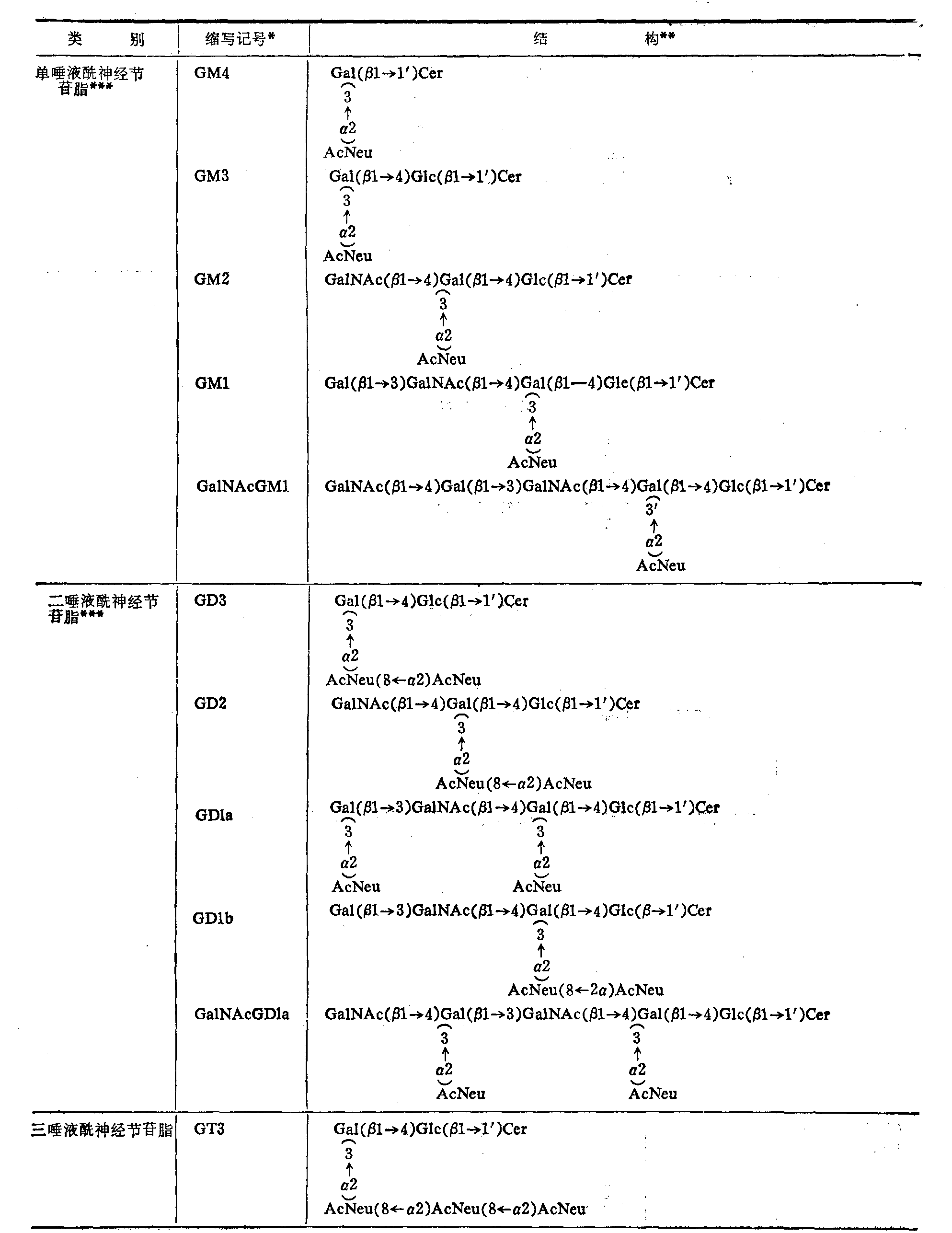
(续表)
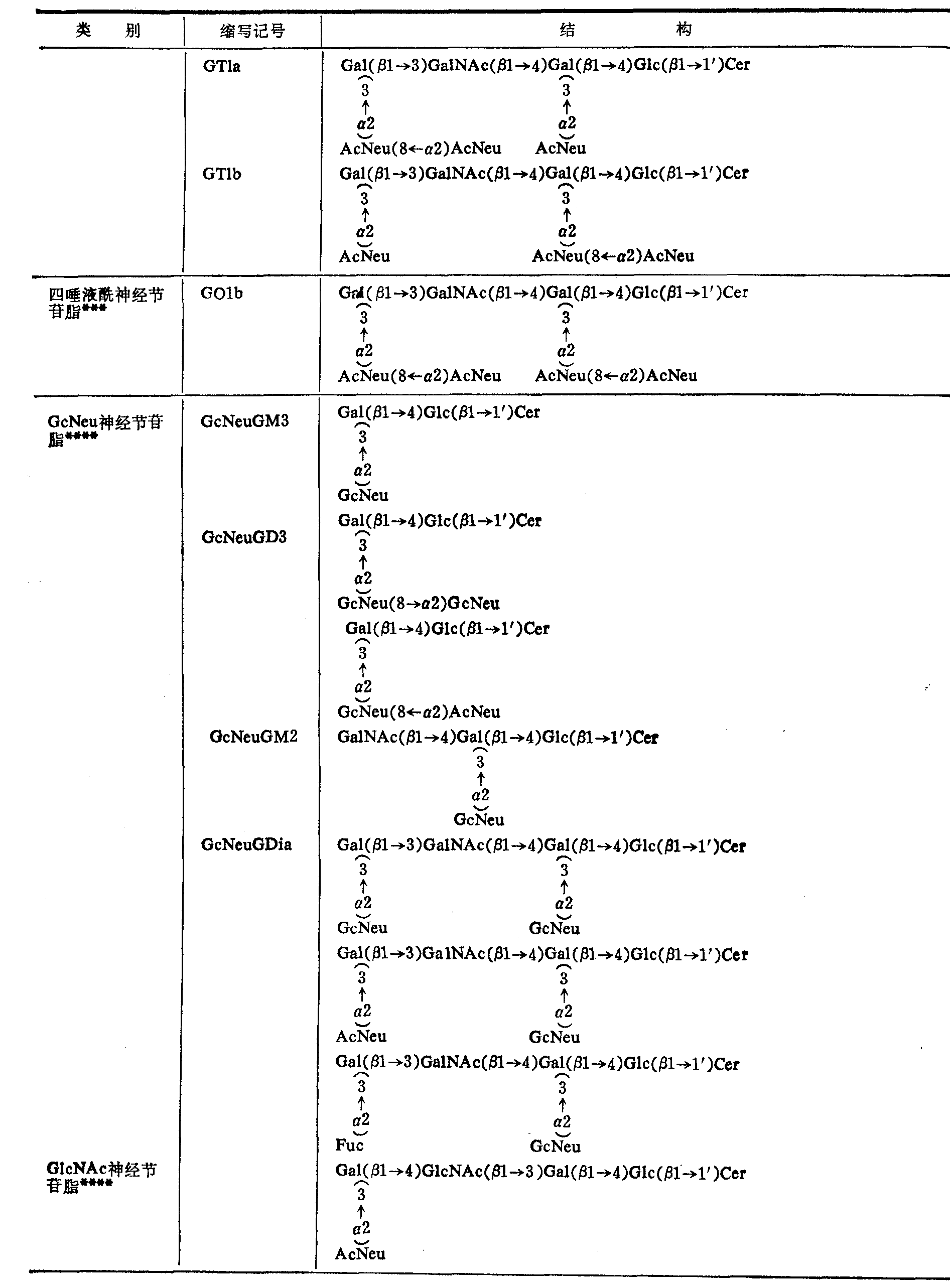
(续表)

*按Svennexhalm
**AcNeu: N乙酰神经氨酸基(即唾液酰基)GcNeu: N羟乙酰神经氨酸基;其他与上表相同
***这类神经节苷脂主要存在于动物脑组织。
****这类神经节苷脂主要分布于神经组织以外的各种组织,但在人体尚未发现GcNeu神经节脂苷类。
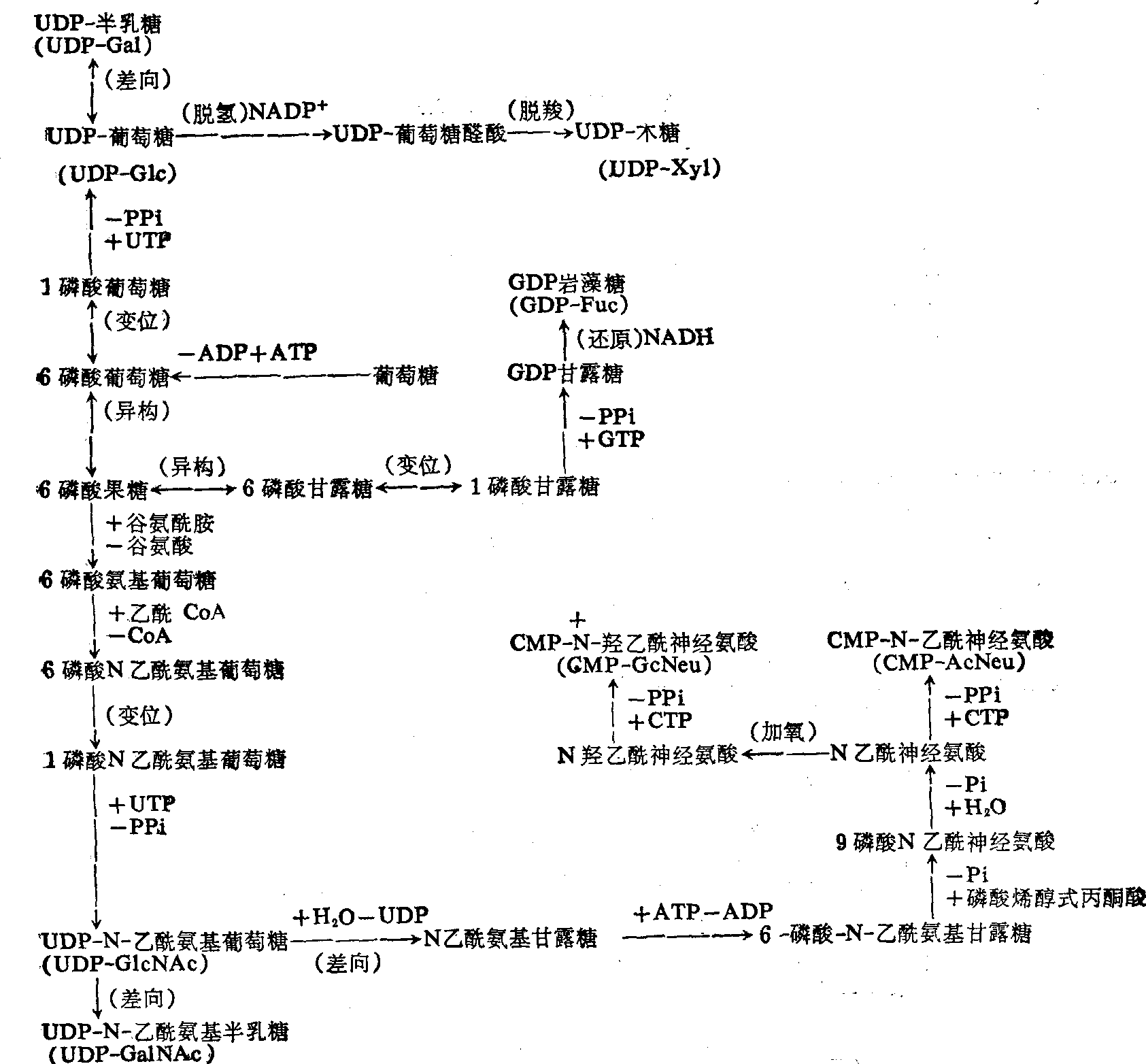
图1 各种糖基单核苷酸的生成途径
(1) 糖基单核苷酸类的合成: 参与糖脂合成的各种糖,首先需要激活成为单核苷酸衍生物,其转变途径大致如图1。
(2) N-酰鞘氨醇类的合成,长链脂酰辅酶A和丝氨酸缩合及脱羧成了酮二氢鞘氨醇,还原为二氢鞘氨醇,再脱氢生成鞘氨醇类,再进行酰化即可形成N-酰鞘氨醇类。离体实验认为,机体内进行的酰化反应有两种,而在生理情况下以微粒体酶系途径为主。
(3) 鞘糖脂的合成. 机体内单糖基鞘糖脂的合成可有两种方式,一是N-酰鞘氨醇的糖化,由糖基转移酶类催化进行,糖基直接来自糖基单核苷酸,即鞘氨醇先N-酰化后糖基化; 二是糖基鞘氨醇的酰化,即鞘氨醇先被糖基单核苷酸糖化,然后被脂肪酰CoA酰化。动物组织内可能以前者为主。
单糖基糖脂又可以在多种糖基转移酶(半乳糖基转移酶、葡萄糖基转移酶、岩藻糖基转移酶、N乙酰氨基葡萄糖基转移酶、N乙酰氨基半乳糖转移酶以及N-乙酰神经氨基转移酶等) 的作用下进一步糖化,逐渐加长糖链,合成不同的糖脂。硫脂类是在硫酸转移酶作用下进行硫酸酯化的,3’磷酸腺苷-5’-磷酰硫酸是硫酸基供体。
鞘糖脂的分解 组织中的鞘糖脂在多种水解酶的共同作用下可以逐步分解。这些酶主要有: 芳基硫酸酯酶(有A、B、C三种)、β-葡萄糖苷酶 (有膜结合性和可溶性两种)、β-半乳糖苷酶(有Ⅰ、Ⅱ两种)、α-半乳糖苷酶(有A、B两种)、α-岩藻糖苷酶、β-N-乙酰氨基己糖苷酶(有A、B、C三种)、N-乙酰神经氨酸酶(唾液酸酶)以及N-酰基鞘氨醇酶等。这些酶主要分布于溶酶体中,但血清等体液中也含少量。高等动物体内的各种糖苷酶通常全属外切糖苷酶,只能作用于糖脂中糖链的非还原末端苷键,对组成糖苷键的糖的种类和α,β构型具有特异性。
糖脂的生物学功能 糖脂只是由糖与脂两类物质组成,故具有与磷脂相似的兼溶性特性,是构成某些生物膜的原料。此外,糖脂所含的糖不只一种,链长不一,其中单糖的排列顺序亦有别,而且其间的连接键的位置也不尽相同,足见糖链的种类各式各样,因此其生物学性质和功能就各具特点。
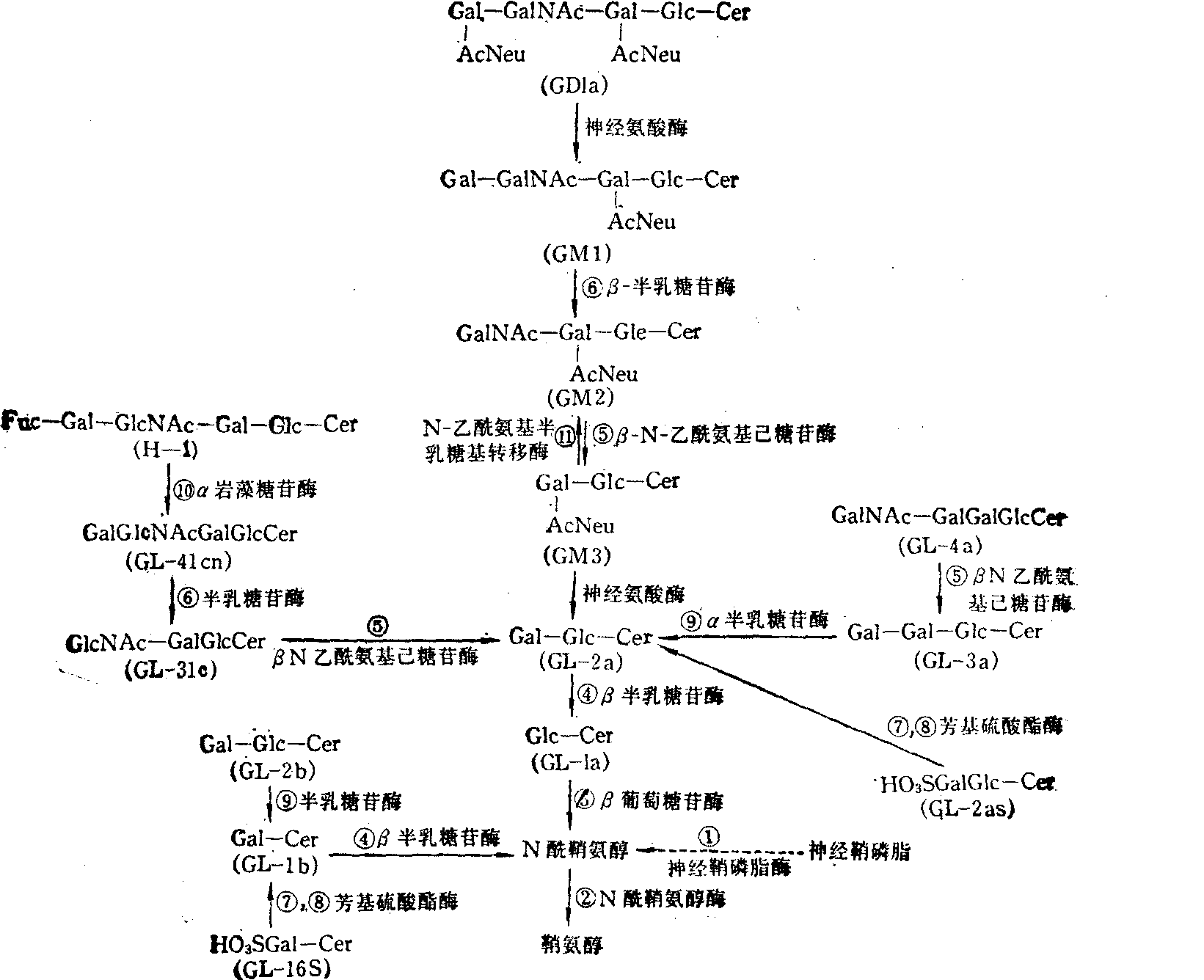
图2 鞘糖脂类的分解途径及有关酶类(数字表示表5所列代谢障碍病)
糖脂,尤其是糖鞘脂,虽然也是生物膜的一种组成成分,但与磷脂在分布上不同,只存在于双分子层的外层。迄今所有研究都表明,哺乳动物细胞的质膜都含有糖鞘脂,而且全部都在膜的外侧,其糖链均暴露于细胞表面。糖是高度亲水的,除非消耗很多能量,要将糖脂中糖的部分置于双分子层疏水烃链核心中是不可能的。这样看来,糖在糖脂分子中起着定向的作用,使糖脂不能从膜的外侧挪到内侧,以维持生物膜两面的不对称性。
含糖脂最多的膜可能要算髓鞘质,超过其干重的20%(见“磷脂”条图5)。髓鞘质实际只是神经鞘细胞的质膜。神经鞘细胞确实将其胞质及核挤到一边,以其质膜多层裹绕末梢神经轴索及树突,感觉神经节细胞,以及中枢神经白质等,使其绝缘,以利神经传导。如此看来,糖脂在构成神经元的绝缘物质上占有一定地位。
多种组织的细胞表面及某些体液中均存在有抗原性物质。红细胞表面的血型物质就是一些抗原性物质。在遗传上,人的血型分类已经分辨出的有14种,共包括上百种的血型抗原。每一分类均含有一组结构上相关的抗原。只有ABO及刘易斯两种分类的血型物质在化学结构上已经详细弄清。其抗原分别称为A,B,H及Lea,Leb,都是一些糖蛋白及糖脂,但其决定子则存在于糖链中。虽然糖链的一级结构有些差异(表3),但其无还原性末端的差异才是起决定作用的,而且影响输血的抗原只是A、B及H而已,这在糖脂及糖蛋白都是如此。此外,刘易斯抗原,Lea及Leb,不是在红细胞成长期间即在其膜中合成,而是成熟红细胞从血浆脂蛋白吸收而来的糖鞘脂。
糖在细胞表面还可能起着彼此互相识别的作用。不同的细胞能彼此识别并互相作用而结合成为一种组织,同时亦能查出不应有的外来细胞,便于清除。免疫过程首先依靠的就是细胞表面的识别。这种识别能力是由糖脂及糖蛋白的糖链结构所赋予。上面已提到糖链的结构有多种多样,这给予细胞互相识别的能力可能是较强的。
糖脂贮积病 在正常情况下,糖脂,主要是糖鞘脂,与其它结构物质一样,在体内是不断合成,同时也不断分解,以维持其恒定的状态。如果在合成或分解代谢中,由于遗传上的原因,造成一种酶的欠缺,以致一个反应不当或根本不能进行,就会发生先天性疾患。糖鞘脂代谢中的先天性疾患有多种,多发生于分解代谢中,且多为溶酶体中水解糖链的某一特殊酶的缺乏或部分缺乏,因而分解代谢到相应的步骤即不能进行或进行甚慢,以致相应的分解产物在溶酶体中出现堆积,故常称这异常现象为糖脂贮积病或鞘脂沉积病。
糖鞘脂在神经组织中含量最多,因此当其分解代谢出现障碍时,往往在神经中发生异常症状。Gaucher病是最早被研究的一种糖脂贮积病,是遗传上有缺陷的结果,影响许多种组织,证明为溶酶体中缺乏β葡萄糖苷酶,造成葡糖苷脂酰鞘氨醇的堆积,如果发生在成年人中,肝、脾肿大,而后者可为正常的4或5倍; 如果在少年时期,影响脑较为明显,故智力发育不全。Fabry病是缺乏α半乳糖苷酶的一种先天性糖脂贮积病,表现为三糖苷脂酰鞘氨醇堆积,主要在肾,其次在肠、淋巴等中堆积,以致逐渐损害肾及其它器官的功能。Tay-Sachs病及Sandhoff病均为缺乏β、N乙酰氨基己糖苷酶的糖脂贮积病,其不同之处是前者缺乏酶的A型,而后者A及B型都缺乏,故前者在脑及脾中只堆积大量GM2(Gal
 及无唾液酸GMa, 而后者除此之外,还沉积四糖苷脂酰鞘氨醇等。其它的先天性糖脂贮积病不在此一一叙述,一并列于表5中,且在图2中标示。
及无唾液酸GMa, 而后者除此之外,还沉积四糖苷脂酰鞘氨醇等。其它的先天性糖脂贮积病不在此一一叙述,一并列于表5中,且在图2中标示。表5 鞘脂类沉积病的临床生化特征
| 图2 编号 | 病 名 | 缺陷的酶 | 组织主要沉积物 | 主要受害组织 | 临床检验标本 | ||||
| 血细 胞 0 0 0 | 体细 胞* 0 0 0 | 血 清 | 尿 | 脏 器 0 0 | |||||
| ❶ ❷ ❸ | Nieman-Pick病 Farber病 Gaucher病 | 神经鞘磷脂酶 N-酰鞘氨醇酶 β葡萄糖苷酶 | 神经鞘磷脂 N-酰鞘氨醇 GL—la | 肝、脾、骨髓,网状内皮 肝、肾、皮下结节 肝、脾、骨髓、网状内皮 | |||||
❹ ❺ | Krabbe病 GM2神经节苷脂病 | β半乳糖苷酶 | GL—2a GM2,无唾液酸GM2 | 脑 | 0 | 0 | 0 | 0 | |
| Tay-Sachs病(B变型) Sandhoff病(O变型) AB变型 | βN乙酰氨基已糖苷酶A βN乙酰氨基已糖苷酶A、B βN乙酰氨基已糖苷酶活性因子 | GM2,GL—31c,GL4a GM2 GM1,GL—4a | 脑、脾 脑、肝、肾 脑 | 0 0 | 0 0 | 0 0 | 0 0 | 0 0 0 | |
❻ | GM1神经节苷脂病 | β半乳糖苷酶 | GL—1bS,GL,—2aS | 脾、肝、肾 | 0 | 0 | 0 | 0 | |
❼ ❽ ❾ ❿ (11) | 异染性白质变性病 多发性硫酸酯酶缺陷病 Fabry病 岩藻糖病 GM3神经节苷脂病 | 芳基硫酸酯酶A 芳基硫酸酯酶A,B、C及胆固醇 硫酸酯酶等 α半乳糖苷酶 α岩藻糖苷酶 βN乙酰氨基半乳糖基转移酶 | GL—1bS,GL—2aS 胆固醇硫酸酯等 GL—3a,GL—2b H—I GM3 | 脑、肾、肝 脑、肾、肝 肾、血管 肝、脾、脑 脑 | 0 0 0 0 | 0 0 0 0 | 0 0 0 | 0 0 0 0 | 0 0 0 0 0 |
☚ 磷脂 血脂 ☛
- 水仙丹是什么意思
- 水仙亭是什么意思
- 水仙图是什么意思
- 水仙图是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子是什么意思
- 水仙子·夜雨是什么意思
- 水仙子·讥时〔双调〕是什么意思
- 水仙子(二首)是什么意思
- 水仙散是什么意思
- 水仙散是什么意思
- 水仙春梅图是什么意思
- 水仙根是什么意思
- 水仙根是什么意思
- 水仙根是什么意思
- 水仙根是什么意思
- 水仙桃是什么意思
- 水仙桃是什么意思
- 水仙桃是什么意思
- 水仙欲上鲤鱼去,一夜芙蓉红泪多是什么意思
- 水仙膏是什么意思
- 水仙膏是什么意思
- 水仙膏是什么意思
- 水仙花是什么意思
- 水仙花是什么意思
- 水仙花是什么意思
- 水仙花节是什么意思
- 水仙阿溪是什么意思
- 水代法制油是什么意思
- 水份是什么意思
- 水会是什么意思
- 水伤药是什么意思
- 水伤药是什么意思
- 水伤药是什么意思
- 水位是什么意思
- 水位是什么意思
- 水位下降是什么意思
- 水位表是什么意思
- 水体放射性强度是什么意思
- 水体污染是什么意思
- 水体污染是什么意思
- 水体污染是什么意思
- 水体污染是什么意思