遗传负荷
生物群体由于有害基因的存在而使其适合度降低的现象。通常用一个群体中每个个体所带有的有害基因的平均数量来衡量。参见“适合度”。
遗传负荷
生物群体中存在着遗传变异,其中也含有在纯合时有害的基因。由于群体中有这样的基因,就使群体的适应度降低。这降低部分是群体必须付出的代价,所以称为遗传负荷。遗传负荷的概念是Haldane(1937)首先提出来的,以后Muller(1950)把它联系到人类的未来而受到重视,最后,Crow(1958)对这概念进行深入研究而使之系统化。
遗传负荷由于产生的原因不同,可分如下几类:
(1) 突变负荷: 基因在杂合时并不有害,或仅有轻度危害,从而基因型AA和Aa的适应度几乎是一样的,只有基因型aa是不利的。在这种情况下,a基因频率很低,由反复突变来补充。由于反复发生有害突变,群体中隐性基因累积,形成了选择上不利的隐性纯合体,从而使群体承受了适应度降低的负担。因这负荷起因于反复突变,所以称为突变负荷。
(2) 分离负荷: 基因处于杂合时有最高的适应度,比任何一个纯合体都更适应。例如杂合体Aa的适应度比纯合体AA和aa都高。这样的基因位点就成为多态的,这位点上的任何一个等位基因都有不太低的频率。但最适的杂合体由于基因的分离可形成较为不适的纯合体,从而使群体担负了适应度降低的负荷。这种负荷即为分离负荷。又由于这负荷起源于超显性现象,所以也称超显性负荷。
(3) 替换负荷: 环境变化后,以前选择上不利的基因转为有利。起初,新的有利基因频率低,但由于选择上有利,逐渐取代环境改变前最适的等位基因。在替换过程中,由于群体中存在有环境改变后而适应度降低的等位基因,从而使群体承受了平均适应度下降的负荷,即替换负荷。因为这负荷起因于进化过程中由更适基因来替换较不适应的等位基因,所以也称进化负荷。
(4) 不相容性负荷:某些基因型间出现不相容现象。如Rh阳性婴儿(基因型Rhrh)出生于Rh阴性母亲(基因型rhrh)时,由于母亲的抗Rh抗体有可能通过胎盘进入胎儿血流中,所以婴儿有患新生儿溶血症的危险。这种基因型间的相互作用引起了群体适应度的减少,从而使群体承担了负荷。这种遗传负荷称为不相容性负荷。此外尚有由于突变体的移入而使群体有不利等位基因的输入负荷,以及由于近亲结婚而使群体中有害隐性纯合体的比例增加的近婚负荷等。
在上述各种负荷中,突变负荷与分离负荷较为重要,其他各种负荷对个别一些位点而言,可能占有重要位置,例如不相容负荷对Rh位点是重要的,但就总共位点来讲,很可能就不重要了。
要测定突变负荷与分离负荷的相对重要性,一个简单的方法是在实验动物中找到纯合时致死的基因,合成致死基因与其正常等位基因的杂合体,然后比较这些杂合体和正常纯合体的生存能力以及把它们的基因传给后代的相对有效程度,即适应度。如果大多数杂合体比较优越,那么遗传负荷主要部分是分离负荷。反之,如果大多数杂合体不及正常纯合体,那么分离负荷只占遗传负荷的小部分,从而可以说主要是突变负荷。木村资生(1975)认为由反复突变产生的突变负荷中,突变率大多很低,而源于选择对杂合体有利的超显性负荷中,选择系数往往较大,所以在一般情况下,突变负荷在遗传负荷中所占的地位远远不及超显性负荷或分离负荷。但也有持不同看法的。
遗传负荷可由遗传与环境的变化而增减。群体的混合,群体间基因流动,突变率的改变,以及对杂合体有利的环境因子的更动等都可能影响遗传负荷的增加或减少。遗传负荷的估计方法很多,其中一种方法是利用近婚系数的大小与适应度高低的关系来估计。例如有一个资料是我国某地区近亲结婚与非近亲结婚所生子女在9岁前的累积死亡率(见表)。
近亲结婚与非近亲结婚所生子女在9岁前的累积死亡率
| 组别 | 婚配对数 | 子女数 | 9岁前死亡率 | |
| 数目 | % | |||
| 表亲结婚组 非近亲结婚组 | 203 199 | 1158 1001 | 284 165 | 24.53 16.48 |
从上表可见,9岁前累积死亡率,包括流产和早产在内,表亲结婚组为24.53%,一般结婚组为16.48%,两者之差为8.05%。因为一般结婚所生子女的近婚系数F是0,而表亲结婚所生子女的近婚系数F是1/16,这就意味着,有1/16的基因本来应该是杂合的,现在因为他们的父母是表亲而纯合了。所以可以把差数0.0805看作是由于0与1/16之差造成的,因为这个缘故,用1/16来除0.0805,
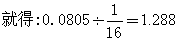 这个数值代表每个配子带有的致死基因数。因为每一个体有两个配子,所以1.288×2=2.576,即每个人大约带有2个以上的致死基因或致死相当量 (所以称为致死相当量,是因为纯合时100%致死效应的基因一个,50%二个,或25%四个不能区别的缘故)。这就是由于近亲结婚而使群体承担的遗传负荷,即近婚负荷。
这个数值代表每个配子带有的致死基因数。因为每一个体有两个配子,所以1.288×2=2.576,即每个人大约带有2个以上的致死基因或致死相当量 (所以称为致死相当量,是因为纯合时100%致死效应的基因一个,50%二个,或25%四个不能区别的缘故)。这就是由于近亲结婚而使群体承担的遗传负荷,即近婚负荷。根据遗传负荷理论,遗传多样性(如由于突变和杂合体优势)要付出一定的代价,因为它们会产生在某一环境条件下选择上不利的基因型个体。但环境条件会随时间经过和地理改变而更动,原来在选择上不利的基因型个体有可能转为有利,所以遗传负荷的估计有时会失之过高。
- 检搜是什么意思
- 检摄是什么意思
- 检操是什么意思
- 检擿是什么意思
- 检收是什么意思
- 检料是什么意思
- 检斤是什么意思
- 检断是什么意思
- 检方是什么意思
- 检晒是什么意思
- 检束是什么意思
- 检束自身是什么意思
- 检束,约束是什么意思
- 检来的孩子是什么意思
- 检柅是什么意思
- 检柙是什么意思
- 检查是什么意思
- 检查、反省自身言行是什么意思
- 检查丈量是什么意思
- 检查上报是什么意思
- 检查举发是什么意思
- 检查书是什么意思
- 检查井是什么意思
- 检查人是什么意思
- 检查修理是什么意思
- 检查值班情况是什么意思
- 检查军马的牧养事务是什么意思
- 检查制度是什么意思
- 检查制止是什么意思
- 检查单是什么意思
- 检查反省自己的错误言行是什么意思
- 检查后修理法是什么意思
- 检查后收下是什么意思
- 检查后答复是什么意思
- 检查员是什么意思
- 检查员(副裁判)是什么意思
- 检查哨是什么意思
- 检查哨兵执行任务是否尽职是什么意思
- 检查哨所是什么意思
- 检查器械是什么意思
- 检查团是什么意思
- 检查坑是什么意思
- 检查城防事务是什么意思
- 检查复习的技能是什么意思
- 检查委员会是什么意思
- 检查完毕是什么意思
- 检查官是什么意思
- 检查审定是什么意思
- 检查察看是什么意思
- 检查察看事物的情况是什么意思
- 检查射是什么意思
- 检查局是什么意思
- 检查并纠正错误是什么意思
- 检查征收是什么意思
- 检查性税务文书是什么意思
- 检查性阅读是什么意思
- 检查房屋是否渗漏是什么意思
- 检查所是什么意思
- 检查手册是什么意思
- 检查扣留是什么意思