cDNA库
与基因库一样,cDNA库也是指一群重组DNA的细菌或噬菌体克隆。差异的是在体外重组DNA时,选用的供体DNA不是来源于生物体的基因组,而是由细胞的mRNA所提供。由于反转录酶的作用,mRNA可以在体外被反向转录合成DNA拷贝。拷贝DNA的核苷酸序列完全互补于模板mRNA,因此这种以mRNA为模板、在体外由反转录酶酶促合成的DNA链,称之互补DNA (cDNA)。反转录形成的DNA链是单链。若再以此单链DNA为模板,用大肠杆菌DNA聚合酶Ⅰ可以合成第二链,得到双链DNA。当起始的mRNA制剂包含了某细胞存在着的各种mRNA分子,被合成的cDNA产物将是各种mRNA拷贝的群体。把它们和载体DNA重组,并转化到宿主细菌里或者包装成噬菌体颗粒,得到一系列克隆群体。每个单一克隆只含一种mRNA的信息,足够数目克隆的总和就包含着细胞的全部mRNA的信息。因此这样的克隆群体叫做cDNA库。
在基因库中,克隆所含的外源性DNA片段来自基因组,因此插入的片段包括有可被转录和翻译的序列,也可能含有不被转录的序列。对于真核细胞的基因来说,大部分结构基因中含有插入序列,它们可以被转录于初级的转录产物(mRNA的前体)中,但在由mRNA前体变为成熟的mRNA的加工过程中,这些不能翻译的序列将被除去。而cDNA库中每一插入片段,由于都是来自mRNA的反转录产物,因此所含序列既可被转录同时又可被翻译。从cDNA库中分离得到的外源性DNA片段,均可直接用于基因的表达。所以在基因工程中真核细胞的cDNA库往往比基因库更为有用。
但是在所含基因信息的代表性来说,cDNA库就有一定的局限性。特别是高等生物如哺乳动物,不同种细胞有着不同的细胞结构和组份,也承担着不同的生理功能,因此其mRNA的成分和量随细胞种类不同而不一致,就是同种细胞,它在分泌、成熟不同阶段中mRNA的活力和成分也不尽相同。因此,要想从cDNA库中筛选出有兴趣的、可直接用于表达研究的基因,首先得知道其相应的mRNA存在于哪种组织中,从这些细胞的cDNA库中才有可能被分离。胰岛素的mRNA仅出现于胰脏的胰岛细胞内,要制备胰岛素的cDNA只能取胰岛细胞来抽提mRNA以及从胰岛细胞的cDNA库去筛选。肝细胞cDNA库中就不能分离到胰岛素mRNA的拷贝。基因库则不然,同一生物体的不同器官或组织抽提出来的基因组所构建的基因库具有相同代表性。
近年来cDNA的制备技术和克隆技术的越趋成熟和更为有效,加上筛选克隆方法的不断改良,尤其是cDNA可被克隆到表达型载体直接表达基因产物,并可用抗体作探针进行免疫筛选,这样从真核细胞cDNA库来筛选某一特异真核基因的路径被广为采纳。cDNA库的构建过程简述如下。
1. mRNA的分离: 在真核细胞中一般每105个细胞才含有1μg的RNA,而其中80~85%是核糖体RNA,10~15%为小分子RNA,mRNA仅占总RNA的1~5%。mRNA分子是不均一的,包含几千到上万种不同的特异序列,密集着各种细胞结构的成分、蛋白质和酶等等。而且不同序列的mRNA拷贝数不同,一般预计有30% mRNA拷贝数低于14挎贝/细胞,有的甚至只有一个拷贝。因此制备mRNA要防止RNase酶活力。所用玻璃器皿的干烤消毒,或用二乙基焦碳酸酯溶液浸泡除去RNase活力。其余化学试剂也要经过处理,在有mRNA参加的各种反应过程中加入能抑制RNase活力的试剂,例如常用的RNasin (一种由鼠肝或人胎盘中提取出的具有抑制RNase活力的蛋白质)。
mRNA的分离过程包括:
❶制备总RNA;
❷采用Oligo-dT纤维素的亲和层析柱分离polyA+RNA,因为大多数的mRNA在其分子的3′末端具有polyA的序列。制备总RNA的方法常用的是在异硫氰酸胍存在下裂解细胞,裂解液用CsCl梯度离心收集RNA沉淀。但也可以采用酚法抽提RNA,或用尿素-氯化锂分离RNA。
mRNA的活性可以采用兔网织红细胞裂解液或麦胚的体外翻译体系来检测。
2. cDNA第一链的合成: 由于反转录酶的发现和应用,使mRNA能在体外反转录成DNA。人们最早从一种RNA病毒(禽成髓细胞白血病病毒,AMV)中分离出反转录酶,它是以RNA为模板的DNA聚合酶,其作用是由5′→3′方向延伸DNA链。要合成与模板mRNA相同链长的cDNA,关键在于反转录酶的质量。酶制剂不能杂有RNase活力。另外在反应时常加入RNase抑制剂(如RNasin)以保护模板mRNA不被降解。酶和模板(mRNA)的合适比例也是得到全链长cDNA的重要因素。此外与酶反应有关的离子种类、离子强度、pH等等也不容忽视。
3. cDNA第二链的合成: 所合成的第一链在3′端形成一个发夹形结构的原因虽不清楚,但它可成为合成第二链的引物。一般在合成第二链时必须首先除去与cDNA第一链氢键相连的mRNA,最简单的方法是碱水解。其次加入大肠杆菌DNA聚合酶Ⅰ(或Klenow酶,或反转录酶),及四种脱氧核苷三磷酸(dNTP)。合成得到的双链cDNA其一端有一环状单链,是发夹式结构,这可用核酸酶S1酶解作用除去。
近来有另一种合成双链cDNA的方法,即在第一条cDNA链合成之后,不必先用碱处理除去mRNA,而是在反应体系中随四种脱氧核苷三磷酸dNTP加入三种酶:RNaseH、大肠杆菌DNA聚合酶I和大肠杆菌DNA连接酶,直接合成第二链。
RNase H是一种水解RNA链的核酸酶,但只有当RNA和DNA以氢键形成杂合链时,RNase H才能发挥作用,将RNA水解 (图1)。所以实际上RNase H是DNA研究中的工具酶。
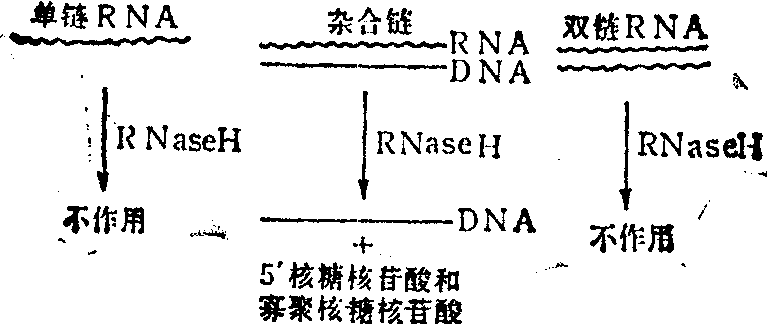
图1 RNase H作用特性
在cDNA链的合成中,第一条链cDNA和mRNA的杂合链正是RNase H的最适底物。加入RNase H后,酶作用mRNA分子形成缺口,提供类似Okazaki样的RNA引物。在此引物下大肠杆菌DNA聚合酶I合成DNA链,并以DNA链替代RNA链,得到一组DNA片段,然后大肠杆菌DNA连接酶把各单链DNA片段连接在一起(图2)。其中连接DNA片段一定要用大肠杆菌DNA连接酶,以防止双链DNA的平端连接。
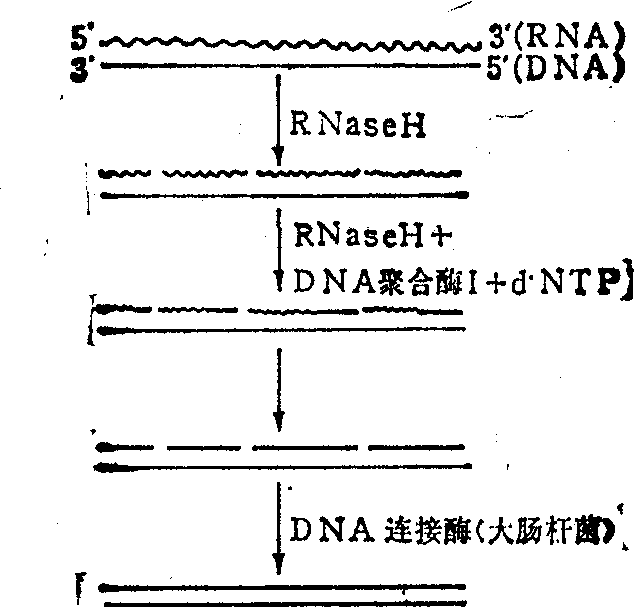
图2 RNase H在cDNA合成中的应用
4. 双链cDNA的修饰以及与载体的连接:双链cDNA合成后的修饰取决于应用何种载体和如何与载体相连接。常用方法为:
(1)接均聚体尾巴:不少双链cDNA与质粒pBR322载体连接,而且较多选用PstI切口,这样就在cDNA3′-OH端藉末端转移酶作用加入OligodC均聚体尾巴,而在pBR322 PstI切口的3′-OH接上OligodG尾巴。插入片段与pBR322可借dG和dC间的互补氢键而粘连在一起。由此种接尾得到的重组体DNA往往不必用连接酶连接,可以直接转入宿主细胞,在细胞内完成双链补齐的连接,并再行DNA复制。
(2)接人工的寡核苷酸接头:目前构建cDNA库时常选用表达型载体λgt11。把cNNA片段藉EcoRI位点插入半乳糖苷酶基因的3′末端,插入基因藉乳糖操纵子的启动子随半乳糖苷酶的基因表达而表达,形成融合蛋白。然后用插入片段相应的抗体作探针,将此克隆从cDNA中选出。在构建cDNA库时为了使cDNA末端带有EcoRI粘性末端,双链cDNA的修饰就包括:先以EcoRI甲基化酶作用修饰cDNA链中EcoRI酶识别序列,使之不被EcoRI相切,再在cDNA平末端接上EcoRI连接子(经常采用三种不同长度8、10、12核苷酸的寡聚体),并用EcoRI酶切产生EcoRI末端。修饰后cDNA可直接与质粒或噬菌体载体相接。当用噬菌体载体DNA相连之后需要经体外包装成噬菌体颗粒。重组的噬菌体颗粒可直接供特异基因的筛选,也可扩增之后在4℃长期保存,供筛选基因用。
当与质粒载体重组,重组质粒需转化到合适宿主细胞中。
一个完整的cDNA库必须包含有所用细胞内全部种类的mRNA,一个典型的哺乳动物细胞含有1万至3万不同的mRNA序列。而且不同种mRNA在细胞内拷贝数也不同,有部分种类的mRNA是多拷贝的,而大部分种类mRNA在细胞中拷贝数不多,属于低含量的mRNA。有一个公式可以计算一个完整的cDNA库所需克隆数目:
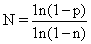
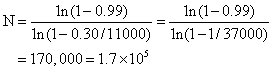
 克隆才能找到一个单拷贝的mRNA表达的克隆。
克隆才能找到一个单拷贝的mRNA表达的克隆。有时不一定要构建全cDNA库,可以先把相应mRNA富集之后,再来构建cDNA克隆,筛选的概率就更高了。
- 空手归航是什么意思
- 空手成家是什么意思
- 空手打空拳是什么意思
- 空手打空拳。是什么意思
- 空手打虎是什么意思
- 空手抓白鱼——难逮是什么意思
- 空手拉脚是什么意思
- 空手挖萝卜——一个个提拔是什么意思
- 空手捋虎须是什么意思
- 空手捞白鱼是什么意思
- 空手揉球法是什么意思
- 空手搏斗是什么意思
- 空手白嚼是什么意思
- 空手白舌是什么意思
- 空手白荡是什么意思
- 空手而去是什么意思
- 空手而回是什么意思
- 空手而归是什么意思
- 空手而归;满载而归是什么意思
- 空手而返是什么意思
- 空手荡脚是什么意思
- 空手荡臂是什么意思
- 空手袖、剑锋懊恼。是什么意思
- 空手裸面是什么意思
- 空手赶不上挑担的,挑担的撵不上要饭的是什么意思
- 空手跑进中药店是什么意思
- 空手进衙门——非输不可是什么意思
- 空手道是什么意思
- 空扑一场是什么意思
- 空扒拉是什么意思
- 空打涝是什么意思
- 空投是什么意思
- 空投任务是什么意思
- 空投伞是什么意思
- 空投信号是什么意思
- 空投信袋是什么意思
- 空投包装是什么意思
- 空投地雷是什么意思
- 空投场是什么意思
- 空投场净空区是什么意思
- 空投场指挥是什么意思
- 空投场标志是什么意思
- 空投平台是什么意思
- 空投广告是什么意思
- 空投水雷是什么意思
- 空投油料补给装备是什么意思
- 空投系统是什么意思
- 空投航线是什么意思
- 空投计划是什么意思
- 空投训练是什么意思
- 空投进入方向是什么意思
- 空投高度是什么意思
- 空报头是什么意思
- 空拃白手是什么意思
- 空担那份咸心是什么意思
- 空担那份咸(闲)心是什么意思
- 空拉弓弦而不放箭是什么意思
- 空拉拉是什么意思
- 空拢拢是什么意思
- 空拳是什么意思