核糖体ribosome
细胞内参与合成蛋白质的颗粒状结构。亦称核糖核蛋白体。其功能是根据信使核糖核酸(mRNA)的指令将氨基酸按照一定的顺序连接成多肽。除极少数高度分化的细胞外,所有细胞都含有核糖体。
结构与分布 直径为15~27nm,由大小两个亚基组成(见表)。大小亚基聚合或解离,取决于胞质中的离子,尤其是Mg2+的浓度。核糖体大亚基形似单人沙发,小亚基两端较大,分别称“头”和“体”,由较窄小的“颈”部连接。核糖体上有许多结合和反应部位,其中小亚基上有mRNA的结合部位,大亚基上有形成肽链的部位,即转肽酶的活动中心。此外,还有由大小亚基共同构成的A部位和P部位,A部位是氨酰基tRNA的结合部位,携带氨基酸的tRNA首先进入这个部位,P部位是肽基tRNA的结合部位。
各类生物中核糖体的沉降系数
| 核糖体来源 | 核糖体 | 大亚基 | 小亚基 |
| 细菌、叶绿体 植物细胞质 动物细胞质 线粒体(哺乳类)* | 70S 80S 80S 55S | 50S 60S 60S 35S | 30S 40S 40S 25S |
* 因种类而异。
细胞内核糖体的数量和分布由蛋白质合成的旺盛程度决定。合成旺盛的细胞,核糖体多,否则较少;同一细胞内蛋白质合成旺盛区域的核糖体也较多。真核细胞的线粒体和叶绿体也有各自的核糖体,可合成部分蛋白质(见线粒体、叶绿体)。
化学组成 蛋白质约占60%,rRNA约占40%。原核细胞核糖体小亚基含有16S rRNA和21种蛋白质;大亚基含有23S rRNA、5SrRNA和34种蛋白质。真核细胞核糖体小亚基含有18S rRNA和34种蛋白质(物种间有较大差异); 大亚基含有28S rRNA(植物为25S rRNA)、5S rRNA (有些真核细胞的大亚基尚有5.5S、5.8 S或7S rRNA)以及39种蛋白质。核糖体还含有少量多胺,如尸胺、四甲烯二胺、亚精胺及Mg2+、Ca2+、Cr3+、Mn3+、Ni3+、Fe3+等离子。目前除已知亚精胺、Mg2+离子有促进核糖体亚基的聚合作用外,其他物质的作用尚不十分清楚。
存在状态 在细胞中与膜结合时称附着核糖体,游离存在时称游离核糖体。真核细胞的附着核糖体主要结合在粗面内质网上,合成的蛋白质多数是分泌蛋白和膜结合蛋白。游离核糖体一般 为被“锚锭”在微梁网络上,主要合成细胞本身生长、繁殖所需要的蛋白质,如结构蛋白、基质蛋白和酶蛋白等,故在未分化细胞和肿瘤细胞中数量较多。但也有例外情况,如大鼠肝细胞的输出蛋白中85%由附着核糖体合成,15%由游离核糖体合成。原核细胞缺内膜系统,其附着核糖体结合于质膜内表面。
核糖体与蛋白质合成 在蛋白质合成过程中,小亚基在起始因子的参与下和mRNA结合,形成小亚基与mRNA的复合体;接着蛋氨酰tRNA 的反密码子与mRNA的起始密码结合,大亚基再结合上去,这时蛋氨酰tRNA处于核糖体的P部位,而A部位处于空位状态;第二个氨酰基tRNA在延长因子的参与下进入核糖体A部位,在转肽酶的催化下,两个 tRNA所带的氨基酸之间通过肽键相连,第二个tRNA成为连接有两个氨基酸残基的肽基tRNA,而第一个tRNA脱离核糖体。当上述反应完成后,在移位酶的作用下,核糖体从mRNA的3L末端作相对位移移动一个三联密码,肽基tRNA从核糖的A部位移到P部位使A部位成为空位;接着第三个氨酰基tRNA进入A部位,如此反复进行,使肽链不断伸长,直到核糖体的A部位到达mRNA的终止密码为止。在终止因子的参与下,多肽链从最后一个tRNA上脱落,mRNA、tRNA亦脱离核糖体,核糖体本身解离成大小两个亚基。
实际上蛋白质合成过程中,往往由几个乃至几十个核糖体与一个mRNA分子结合,高效率地进行蛋白质的合成。多个核糖体串连在一个mRNA分子上时称多聚核糖体。mRNA分子越长则串连的核糖体数目越多。
核糖体ribosome
又称核蛋白体或核糖核蛋白体(ribonucleoprotein)。核糖体核糖核酸(rRNA)和蛋白质结合成的无包膜胞质颗粒。普遍存在于各种细胞中。直径约15~25nm,由大小不同的两个亚单位(亚基)结合而成。依其与内质网的关系分为两类,一类散于细胞质中,称游离核糖体,另一类附着在内质网膜外表面,称附着(膜旁)核糖体。单个存在时称单核糖体,由mRNA链将多个核糖体串联在一起时称多核糖体,是细胞内蛋白质合成的重要场所。
核糖体hetangti
广泛存在于各类细胞的一种颗粒状细胞器,是细胞内合成蛋白质的场所。1953年罗伯逊(Robinson)和布朗(Brown)用电镜观察到植物细胞内的核糖体颗粒,1955年帕拉登(Palade)观察到动物细胞中核糖体颗粒。1958年罗伯特(Roberts)建议命名为核糖核蛋白体。核糖体的主要化学成分是蛋白质和rRNA。核糖体颗粒的直径一般为15~20毫微米,由大小两个亚单位组成。真核细胞中核糖体类型为80S型,原核细胞为70S型,叶绿体中核糖体也为70S型,但线粒体中核糖体因种类而不同,有55 S到80S多种。80S型核糖体的大小亚单位分别为60S和40S,70S型核糖体的两个亚单位分别为50S和30S。电镜下观察肝细胞的单核糖体,大亚单位略呈半圆形,直径约为23毫微米,有一侧伸出三个突起,中央为一凹陷;小亚单位呈葫芦状,其大小为23毫微米。大小两个亚单位结合在一起时,凹陷部位彼此对应形成一个隧道,在翻译过程中,mRNA穿行于隧道中。在核糖体上有功能活性部位,在蛋白质合成过程中,核糖体上的功能活性部位配合作用,把tRNA转运的氨基酸,按照mRNA的信息密码顺序连接起来,形成具有特定氨基酸序列的多肽链。
核糖体
一种直径为10—20纳米的核糖核蛋白颗粒。由两种不等的亚基通过镁离子联结构成。每一亚基由大致等量的RNA与蛋白质组成。
核糖体
核糖体(核蛋白体)是由核糖核酸和蛋白质结合成的一种胞质颗粒,存在于各种细胞,是蛋白质合成的重要场所,首先由Palade发现,故曾称Palade颗粒。此外,在线粒体内腔也有此体,称线粒体核糖体(参见“线粒体”条)。核糖体在电镜下呈近似球形的致密颗粒,大小约为15×25nm,由大小不同的两个亚单位(亚基)结合而成。大亚单位的直径约23nm,略呈圆锥形,两侧稍隆起,底面较平,中有一凹沟,中心有细管自底面通到顶端。小亚单位的大小约11.5×14×23nm,近似扁椭圆形,侧面似弧形,凸面向外,凹面朝内,有一横沟。小亚单位凹面与大亚单位底面结合,其间有一孔道(见图)。
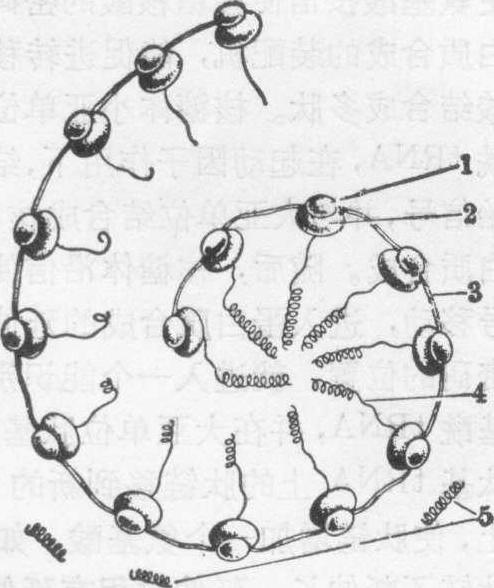
游离的多核糖体
1.核糖体小亚单位 2.核糖体大亚单位 3.mRNA 4.形成中的蛋白质 5.已形成的蛋白质
多核糖体也名多核蛋白体,由3~30个核糖体连接于一个mRNA长分子组成。核糖体由核糖核酸(核糖体核糖核酸,rRNA)和约70种蛋白质所组成。大亚单位为60s,含3种核糖核酸(28s,58s,5s)和约40种蛋白质。小亚单位为40s,含1分子核糖核酸(18s)和约30种蛋白质。在核糖体亚单位上有几个功能中心,在蛋白质合成中起特殊的作用。如小亚单位上有与作为蛋白质合成起始者的蛋氨酰tRNA相结合的部位,并有肽基转移酶活性部位。
单核糖体和多核糖体 单个的核糖体称单核糖体(或单核蛋白体),此时它不具有蛋白质合成的活性。有人认为核糖体未进行多肽合成时或多肽合成结束后,解离成亚单位,分布于细胞质基质的亚单位库中。多个核糖体通过其小亚单位结合于一个信使核糖核酸分子上,形成一条串珠状的结构,称多核糖体,在电镜下,多核糖体呈螺旋状或花簇状,核糖体间的距离约30~35nm。在磷钨酸负染色的标本上,见一条直径1~1.5nm的细丝,即信使核糖核酸分子穿过各个核糖体的大、小亚单位之间。组成多核糖体的核糖体数目与信使核糖核酸的长度(即所合成蛋白质的氨基酸残基数)成比例,合成血红蛋白的多核糖体有5~6个核糖体,合成肌球蛋白的多核糖体有60~100个核糖体。每个核糖体的部位都有一个多肽链正在形成(见图)。
核糖体与蛋白质合成 在分子生物学中,由氨基酸合成蛋白质的过程称翻译,分起动阶段、延伸阶段和终止阶段,需多种成分参与,有信使核糖核酸、转移核糖核酸、核糖体、一些蛋白质因子(起动因子、延伸因子、释放因子)及ATP、GPT和镁离子等。信使核糖核酸(mRNA)是蛋白质合成的模板,它有特定排列的特异的三位体密码子(每个密码子由三个相邻的核苷酸的碱基组成),规定了所合成蛋白质的各种氨基酸的顺序。并在靠近其5′端有起动信号(起动密码AUG),靠近其3′端有终止信号(终止密码UAA、UAG或UGA),确定蛋白质合成的起动与终止。转移核糖核酸(tRNA)是氨基酸的搬运工具,有许多种,每种能与一种特定的氨基酸结合,形成特定的氨基酰tRNA。每种转移核糖核酸上有特异的三位体反密码子,能辨认和翻译信使核糖核酸上相应的特定密码,使氨基酸按信使核糖核酸的密码顺序排列。核糖体是蛋白质合成的装配机,能促进转移核糖核酸所携带的氨基酸结合成多肽。核糖体小亚单位及具有起动作用的蛋氨酰tRNA,在起动因子作用下,结合于信使核糖核酸的起始信号,并与大亚单位结合成核糖体起动复合体,开始蛋白质合成。随后,核糖体沿信使核糖核酸逐渐向终止信号移动,进入蛋白质合成的延伸阶段。每向前移动一个密码的位置,就进入一个能识别与翻译该密码的新的氨基酰 tRNA,并在大亚单位肽基转移酶催化下,使原有的肽基tRNA上的肽链移到新的tRNA所携带的氨基酸上,使肽链增加一个氨基酸。如此,随着核糖体的移动,肽链不断伸长。延伸过程有延伸因子参与。当核糖体移到终止信号,翻译过程即告结束,在终止因子参与下,将所合成的多肽链释放,核糖体也脱离信使核糖核酸重返亚单位库。原有的核糖体不断于终止信号处脱离,新的核糖体不断从起始信号处加入。
游离核糖体和附着核糖体 游离在细胞质中的核糖体,称游离核糖体(或游离核蛋白体),它主要参与合成细胞的结构蛋白质,供细胞生长、分裂和普通功能之需。未成熟的细胞或分化低的细胞,如干细胞、幼稚细胞、胚胎细胞及培养的细胞,尤其分裂活跃的细胞,一般均含丰富的游离核糖体。分化低的癌细胞也含丰富的游离核糖体。附着于内质网表面的核糖体,称附着核糖体(或附着核蛋白体),它主要参与合成分泌蛋白质(参见“粗面内质网”条)。
核糖体
核糖体是蛋白质合成的细胞器,由蛋白质和核糖核酸构成。氨基酰tRNA和mRNA都结合在核糖体上,并由核糖体催化形成肽键等反应。早在40年代就观察到蛋白质合成和细胞中核糖核酸颗粒息息相关,此等颗粒随之称为核糖核蛋白体或核糖体。各种核糖体可按其沉降系数而分为两大类: 80s核糖体和70s核糖体。真核细胞胞质核糖体属前者,原核细胞核糖体属后者。真核细胞细胞器的核糖体如线粒体核糖体也属于70s类。两类核糖体对某些抗生素的敏感性也有区别。80s者对亚胺环己酮敏感,对氯霉素不敏感;70s者则对氯霉素敏感,对亚胺环己酮不敏感。核糖体由两个亚单位组成,它们可逆地缔合和解缔;降低Mg2+可致解缔,恢复Mg2+的浓度又可重新缔合。70s核糖体解缔为大亚单位(50s) 和小亚单位(30s);80s核糖体解缔为60s和40s大小亚单位。
原核细胞核糖体以大肠杆菌者所知最详尽,有关核糖体的实验资料及各种学说大多得自这种细菌。大肠杆菌核糖体质量为2.7×106,其中RNA约占65%,余下为蛋白质。用双向聚丙烯酰胺凝胶电泳分离,可鉴定出30s亚单位有21个蛋白质点,分别名为S1、S2……S21;50s亚单位34个蛋白质点,分别名为L1、L2……L34。小亚单位的21个点是21种蛋白质;大亚单位的L8是L7/L12和L10的复合体,实际只有33种蛋白质。核糖体蛋白质均已经纯化至均一,而且除个别几种外,它们的一级结构亦已测出。它们在组成上都是各不相成的蛋白质,只有二
已测得一级结构的大肠杆菌核糖体蛋白质
| 氨基酸数 | 分子量 | 氨基酸数 | 分子量 | ||
| S3 S4 S5 S6 S7 S8 S9 S10 S11 S12 S13 S14 S15 S16 S17 S18 S19 S20 S21 L1 L3 L5 L6 L7 | 232 203 166 135 177 129 128 105 128 123 117 98 87 82 83 74 91 86 70 233 209 178 176 120 | 25,852 23,137 17,515 15,704 19,732 13,996 14,569 12,290 13,728 13,606 12,968 11,162 10,001 9,191 9,573 8,896 10,299 9,553 8,369 24,599 22,258 20,171 18,832 12,206 | L10 L11 L12 L13 L14 L15 L16 L18 L19 L20 L21 L22 L23 L24 L25 L26 L27 L28 L29 L30 L31 L32 L33 L34 | 165 141 120 142 120 144 136 117 114 117 103 109 99 103 94 86 84 77 63 58 62 56 54 46 | 17,736 14,874 12,164 16,019 13,227 14,961 15,296 12,770 13,002 13,366 11,565 112,111 11,013 11,183 10,694 9,553 8,993 8,875 7,273 6,111 6,971 6,315 6,255 5,381 |
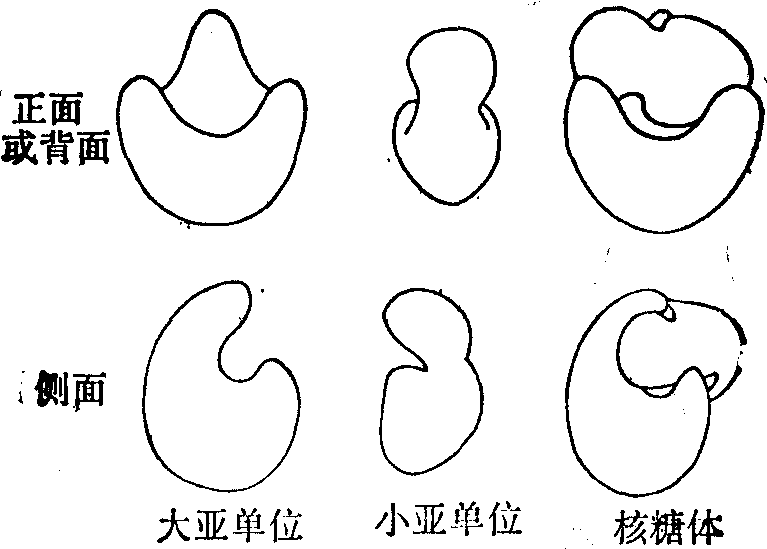
核糖体模型示意图
为了了解蛋白质合成的分子机制,详尽阐明核糖体三维结构实属必要。如同研究酶的结构,理想的是获得2~3A分辨率的X线衍射图,但迄今尚未获得适于这种技术的核糖体结晶。只好采用他法: 免疫电子显微镜法(IEM)、蛋白质交联法、荧光能量传递和中子衍射等。I-EM的根据是抗核糖体各蛋白质的抗体可以在完整亚单位上和相关蛋白质起抗原抗体反应。同时恰当的染色又使核糖体或其亚单位可在电子显微镜下呈现出其形状,并且其甚不对称的形态具有提供很好标志的特异结构,方便了对抗体结合位置的辨认。如用对某一核糖体蛋白质特异的抗体和亚单位作抗原抗体反应,则由于IgG是两价的而形成了亚单位二聚体。抗体的Fab臂结合到亚单位,而结合位置则可视为该特定蛋白质暴露在核糖体该处表面上的抗原决定簇。按此即可勾划出亚单位的形状以及各个蛋白质的抗原决定簇在其上面的位置。此法随对资料的解释的不同而得出迥异的核糖体模型,希望进一步改善方法而求出一致的模型。图示从IEM资料构成出的一种模型。大亚单位形似摇床,小亚单位像胎儿; 核糖体如胎儿安睡在摇床上。用双功能试剂交联蛋白质可以决定蛋白质的相邻关系。从已经交联了的亚单位可得到交联的蛋白质,再用免疫化学或电泳等方法鉴定出相交联的蛋白质。在30s亚单位可以鉴定出三十多对交联的蛋白质,按此可提出在亚单位中各个蛋白质的相互关系。核糖体亚单位中两个蛋白质之间的距离也可应用荧光标记蛋白质的能量传递来测量。两种核糖体蛋白质各共价结合荧光剂,把这衍生蛋白质重组到亚单位中,根据荧光剂之间的能量传递效率就可计算出该两分子的距离。中子衍射可测量两个氘标记蛋白质重组在亚单位里的距离。各种方法各有长短,可相互补充,但核糖体的三维结构尚待解决。
线粒体的核糖体属70s类,解缔为30s和40s亚单位;在组成上和大肠杆菌者相差颇远。30s亚单位含一条RNA和约40种蛋白质;40s亚单位也只含一条RNA,但含约50种蛋白质。线粒体核糖体蛋白质含量高,占65~75%,且不含5s RNA。
真核细胞胞质核糖体质量约为4.6×106,沉降系数约80s。小亚单位质量1.5×106,含1分子18s RNA和约30种蛋白质。大亚单位质量随物种而异,自2.4×106(植物)至3.0×106(哺乳动物)不等,含3种RNA(28s、5.8s和5s)以及约50种蛋白质。80s比70s核糖体大,主要是蛋白质多了些。蛋白质在80s核糖体约占质量50%,而70s只占33%。这两类核糖体参与的蛋白质合成的各反应步骤甚相似,而蛋白质数目相差颇远;这或者是与蛋白质合成的调节机制在真核细胞远为复杂 (尤其在起始阶段)有关,或者是与核糖体要通过核膜有关。
18s RNA(0.7×106)在各物种间都很相近,而28sRNA(~1.7×106)则变化颇多。这两种RNA都经过大量的修饰作用,如Hela细胞的18s和28s RNA分别含46个和71个甲基,还有假尿苷和其他修饰过的碱基及糖。从多种物种的18sRNA中发现其3′端有一共同的八核苷酸片段——GAUCAUUA。人类的18s RNA和爪蟾(Xenopus)者约有60%的共同序列。人、小鼠,大鼠和兔的5s RNA是相同的。5.8s RNA含150~160个核苷酸,其功能相当于原核的5s RNA。而真核细胞的5s RNA都是外加上的。
80s核糖体的约70个蛋白质大多已经分离出,并纯化至均一。大鼠肝细胞核糖体蛋白质,小亚单位者的分子量自11,200~39,000不等,平均约21,400;大亚单位的自11,500~41,800,平均21,200。已经知道了50多种核糖体蛋白质的氨基酸组成。多数是富含精氨酸和赖氨酸的碱性蛋白质。也有少数酸性蛋白质。蛋白质种类的切实数目还未弄清。因方法不同,各实验室报告的数字互有出入。但同一动物的不同器官的核糖体蛋白质在双向聚丙烯酰胺凝胶电泳上是相同的。不同物种核糖体蛋白质也近似,但免疫方法检查大鼠和鸡的核糖体蛋白质,却很不一样。真核细胞核糖体蛋白质和原核细胞者在免疫化学上未见交叉反应。但是动物的L40/L41和酵母的一种酸性蛋白质在功能上却都可和大肠杆菌的L7/L12互换,提示这些蛋白质的高度保守性。真核细胞核糖体结构资料很少,电镜初步所见除大小不同外,形态大致相似。
核糖体
核糖体是细胞的重要结构之一,是合成蛋白质的细胞器,其唯一的功能是将氨基酸装配成肽链。核糖体几乎存在于一切细胞内,不论是原核细胞或真核细胞中均含有大量核糖体,即使是最简单的细胞支原体也至少含有数以百计的核糖体,目前仅发现在无核红细胞等极个别高度分化类型的细胞内没有核糖体,因此可以认为核糖体是细胞最基本的,不可缺少的结构。
核糖体是一种颗粒状结构,直径为15—25nm,主要成分为蛋白质与RNA。蛋白质含量约占40%,RNA约占60%。蛋白质分子主要排列在核糖体的表面。附着核糖体与内质网形成的复合结构,称为粗面内质网(或叫颗粒内质网)。有很多核糖体不附着在生物膜上,呈游离状态,分布在细胞质的基质内,称为游离核糖体。附着在内质网膜上的核糖体与游离核糖体所合成的蛋白质不同,前者主要合成向细胞外输送和分泌性的蛋白质,如抗体、酶原与蛋白质类的激素等。游离的核糖体主要是合成细胞的结构蛋白、基质蛋白与细胞本身所需要的酶蛋白,即合成细胞生长与繁殖所需要的蛋白分子。此外,还参与合成一些特殊蛋白质,如血红蛋白分子等。因此,在一些具有分泌功能的细胞内,粗面内质网比较发达,而在一些未分化的细胞与肿瘤细胞内,游离的核糖体比较多。在原核细胞内,因为没有内质网,虽然有些核糖体附着于细胞膜的内表面,但绝大多数核糖体分布在细胞的基质内,呈游离状态。核糖体常常分布在蛋白质合成旺盛的区域,其数量与蛋白质合成的强度有关系。在细菌培养的指数生长期,每个细胞内大约有数以千万计的核糖体,核糖体含量占细胞干重可达40%。而在饥饿培养状态的细胞内仅有几百个核糖体。在蓝藻细胞内,核糖体常分布在细胞质的中部,与DNA纤维紧密结合,因为在原核细胞内RNA的转录与蛋白分子的翻译是同时进行的。
核糖体的结构与成分 目前已发现在所有生物机体细胞内有两种基本类型的核糖体:一种是70S的核糖体,其分子量为2.6×10 ,所有原核细胞内的核糖体均为70S。真核细胞的核糖体(除线粒体与叶绿体核糖体)分子量为4.1×106,均为80S。不论是70S或80S的核糖体均由两个不同大小的亚单位构成。在高分辨率电镜下用负染法可以观察到核糖体大小亚单位之间有一条沟,大亚单位近似椭圆形,小亚单位颇似扁帽形或杆状,大亚单位在与小亚单位邻近的一面呈扁平,而后者则稍微凹陷,因此在大小亚单位之间形成一条明显的间隙,大小亚单位结合在一起形成球状的核糖体。应该指出,由于核糖体的来源不一样,与样品制备技术不一样,并不是经常能观察到上述的有规则的典型结构。
,所有原核细胞内的核糖体均为70S。真核细胞的核糖体(除线粒体与叶绿体核糖体)分子量为4.1×106,均为80S。不论是70S或80S的核糖体均由两个不同大小的亚单位构成。在高分辨率电镜下用负染法可以观察到核糖体大小亚单位之间有一条沟,大亚单位近似椭圆形,小亚单位颇似扁帽形或杆状,大亚单位在与小亚单位邻近的一面呈扁平,而后者则稍微凹陷,因此在大小亚单位之间形成一条明显的间隙,大小亚单位结合在一起形成球状的核糖体。应该指出,由于核糖体的来源不一样,与样品制备技术不一样,并不是经常能观察到上述的有规则的典型结构。
70S的核糖体广泛存在于各类原核细胞中,从细菌细胞中提取70S的核糖体较为方便。近年证明在线粒体与叶绿体中虽有与原核细胞类似的70S的核糖体,但数量甚少,提取的技术难度较大,对分离提取的核糖体进行分析时,除了最基本的70S核糖体,经常还夹杂30S、50S与100S的三种不同沉降系数的颗粒,许多试验说明,70S的核糖体在溶液的镁离子浓度小于0.001 mol/L,易降解为50S与30S的大小亚单位。当溶液的镁离子浓度大于0.01 mol/L时,2个核糖体常常形成100S双聚体,以上所述的物理化学过程均是可逆的(图1)。
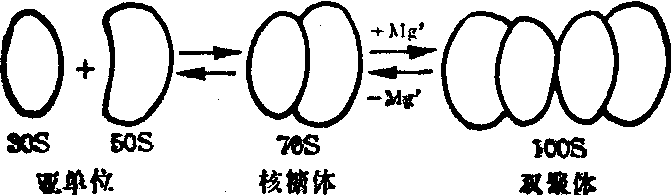
图1 70S核糖体在不同镁离子浓度下的物理状态
对70S核糖体(以细菌的核糖体为例)的结构与成分进一步分析见图2。核糖体先降解为50S与30S的大小亚单位,其分子量分别为1.8×106与0.8×106,将大小亚单位分别用酚提取,并进一步分析核酸与蛋白质的含量;小亚单位中含有一个16S的rRNA分子,分子量为5.5×106约含1650个核苷酸。大亚单位中主要含有一个23S的rRNA分子,其分子量为1.1×106,约含3 300个核苷酸,在大亚单位中经常还有一个5S的RNA分子,分子量为4×106,仅含115—116个核苷酸。从30S的亚单位中已发现含有20种不同的蛋白分子,而在50S的大亚单位中约含34种蛋白分子。
80S的核糖体普遍存在于真核细胞内,对分离的核糖体进行理化特性测定,发现与原核细胞核糖体具有类似的现象。随着溶液镁离子浓度的降低,80S的核糖体可降解为60S与40S的大小亚单位。当镁离子浓度增高时,80S的核糖体可以形成120S的双聚体(图3)。
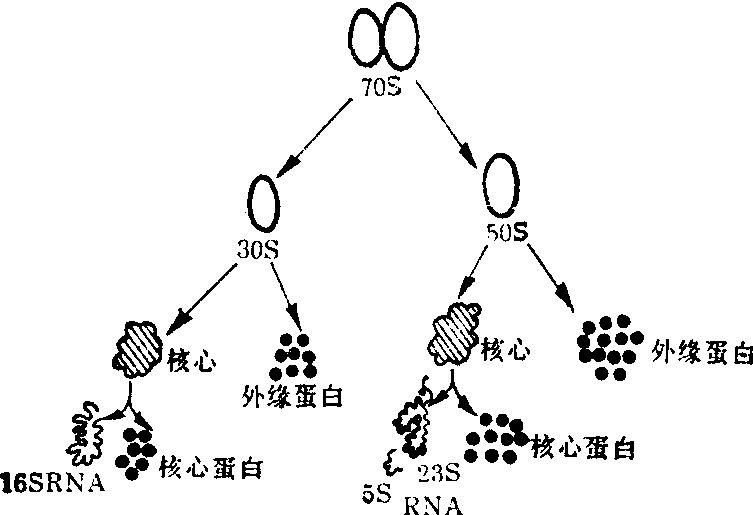
图2 原核细胞核糖体(70S)结构与成分解析
对80S核糖体的结构与成分进一步解剖见图4。核糖体首先降解为60S与40S的亚单位,分子量分别为2.7×106与1.4× 106。小亚单位中含有1个18S的RNA分子,分子量为6×105。大亚单位中含有1个28SRNA,分子量为1.3×106,还含有1个5SRNA分子,有人在大亚单位中分离出5.8SRNA。真核细胞内核糖体也有差异。动物细胞核糖体大亚单位内有28S rRNA,而植物细胞、真菌细胞与原生动物细胞内大亚单位中却不是28SRNA,而是25—26SRNA。在低等生物真核细胞的核糖体内所含rRNA的类型比较复杂,可能不仅限于以上几种rRNA,有人曾发现有25S与16S的rRNA等。小亚单位中约含有23种蛋白质分子,大亚单位中含有30—34种蛋白质分子(图4)。
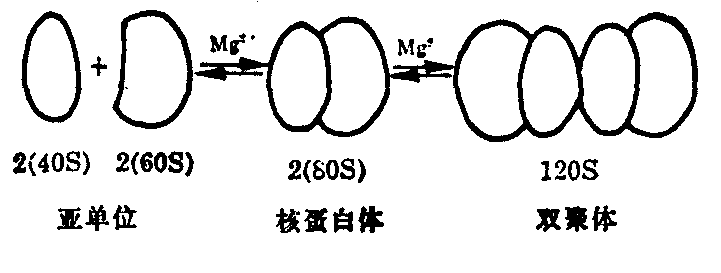
图3 80S核糖体在不同镁离子浓度下的物理状态
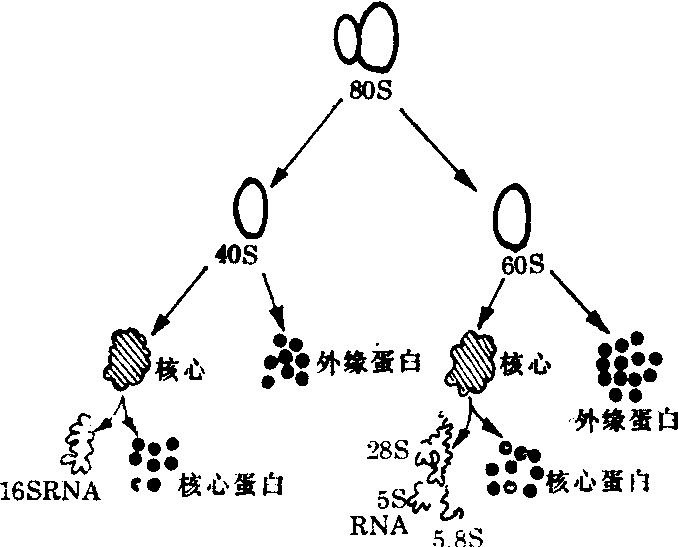
图4 真核细胞核糖体(80S)结构与成分解析
真核细胞核糖体的形成 核糖体的发生与形成是一个较为复杂的过程。在真核细胞内核糖体的主要成分(rRNA与蛋白质)是在核仁内合成。在核仁的丝状区,以DNA(核仁内染色质)为模板,先转录45S的prRNA,prRNA是rRNA的前身物, 由它再转化为28S、5.8S与18S的rRNA; 5S rRNA的转录是在染色质内。rRNA与蛋白质如何装配成核蛋白体的一些具体环节与机制还不完全清楚,但大亚单位的装配(28S rRNA,5.8S rRNA,5S rRNA+蛋白质)肯定是在核仁内进行,小亚单位的装配(18S rRNA+蛋白质)可能主要是在核的基液内进行,因为一系列的试验说明:从核仁可以分离出60S的亚单位,核仁颗粒可能就是大亚单位的前身物,而从核仁中从未分离到40S的亚单位;相反,却可从核液中分离到大量的40S的小亚单位。现在一般认为大、小亚单位分别通过核膜孔进入细胞质,在执行肽链合成功能时,装配成核糖体。
多聚核糖体 核糖体是合成蛋白质的机器,但核糖体在细胞内并不是单个独立地执行功能,而是由很多个或甚至几十个核糖体串连在一起,高效率地进行肽链的合成,具有这种功能与形态结构的核糖体聚合体称为多聚核糖体,它的基本结构是由一条mRNA分子链将核糖体串连在一起,犹如念珠一样。在细胞的超薄切片上,不论是附着在内质网上的核糖体或分布在胞质的游离核糖体,可以经常看到它们排列成簇状、环状、串珠状,甚至雪花状等,这均是多聚核糖体的形态特点。用非常柔和的方法从细胞匀浆提纯的核糖体亦可以看到以上类似的形态特征。每种多聚核糖体所包含的核糖体数量是由mRNA分子的长度与它所合成的肽链分子的大小来决定的,也就是说合成肽链的分子愈长,多聚核糖体就愈长,所含的核糖体数就愈多,串连它们的mRNA分子就愈长。现在将这方面最出色的工作者 (Waner与Rish等)为代表进行叙述,他们发现网织红细胞内负责血红蛋白分子多肽链合成的多聚核糖体基本是五聚体,即由5个核糖体组成。网织红细胞内合成的唯一蛋白质是血红蛋白,它由4个多肽链组成,其分子量为16 000,其中一个多肽链包含150个氨基酸。按照蛋白质翻译的三联密码规律:即三个碱基决定一个氨基酸排列,翻译一条血红蛋白多肽链需要含450个核苷酸的一个mRNA分子作为模板。每个核苷酸长0.34nm,则mRNA的长度应是450×0.34nm=153nm,那么从理论上说,负责血红蛋白肽链合成的多聚核糖体长度应该是153nm。他们用梯度离心技术与电镜负染技术相结合,试验结果令人信服地证实了这一推论的正确性,也观察到网织红细胞内的多聚核糖体是由一条粗约1—1.5nm的mRNA串着5个(有时6个或4个)核糖体,每个核糖体的直径为23nm,从一个核糖体的中心到另一个核糖体的中心为30—35nm,因此多聚核糖体的总长约150nm,这与以上的推论是完全符合的。
实验更进一步证明核糖体之间的连结是非常脆弱的,用一般匀浆梯度离心法均可使多聚核糖体降解为单个蛋白体,只有用非常柔和的方法提取才能保证多聚核糖体的完整性。用核酸酶作用时,多聚核糖体很快就降解为单个核糖体,这就证明串连核糖体的可能是一根单股RNA,而且推测mRNA很可能是通过核糖体大小亚单位之间的沟串连核糖体的。对保证多聚核糖体的完整性来说起主要作用的是mRNA。此后又证实HeLa细胞内可以分离出两种不同的多聚核糖体,一种含5—6个核糖体,这种数量较多,另一种含30—40个核糖体,这种数量较少,这说明在一种细胞内可以同时合成多种蛋白质。脊髓灰质炎病毒在细胞内诱导产生的多聚核糖体含有50—70个核糖体。近年在原核细胞(细菌) 内分离的多聚核糖体都是与DNA结合在一起。一串串的多聚核糖体以其mRNA链直接垂直连结在DNA链上,因为在原核细胞内由DNA转录mRNA与mRNA翻译蛋白质可以同时在一个部位进行。
核糖体与蛋白质合成 核糖体在蛋白质合成中所起作用的具体细节目前尚不能认为已清楚。根据实验与逻辑的推理,一般认为核糖体的具体作用可能有两方面,一是与mRNA的连接,二是为多肽链的形成提供表面位置。大小亚单位所起的作用是不一样的; 小亚单位主要是对mRNA起连接作用,小亚单位上可能有与mRNA连接的专门位置。而大亚单位主要为多肽链的形成提供位置,大亚单位可能具有两个位区,其中一个是多肽链形成点,叫做D点(doner site或peptidyl site),另一点叫做A点,即氨基酸受点(acceptor site或aminoacyl si-te)。
氨基酸以一定顺序装配成肽链,是以mRNA携带的信息为模板、以核糖体为基地进行的。具体说是要以多聚核糖体这样特定结构来执行的(图5)。但还必须要有tRNA的参加,以及一系列必要条件(如足够的氨基酸,必要的聚合酶,能量即ATP的供应与Mg2+、K+等)具备的情况下,多聚核糖体才能完成肽链的合成。肽链的合成过程必须要有以下一些环节:
❶激活的氨基酸在氨酰—tRNA酶的催化下与tRNA一端结合形成氨基酰tRNA。tRNA与氨基酸结合具有严格的专一性。
❷氨酰—tRNA转移到核糖体的氨基酸受点,由tRNA—端的核苷酸反三联密码与mRNA的正三联密码以互补的形式结合,将氨基酸安排在肽链特定序列的位置上。
❸核糖体可能是从mRNA链的5′向3′滑动,在滑动的过程中每一个核糖体按mRNA密码的序列完成多肽链的装配。当一条肽链在一个核糖体上合成完毕,核糖体将多肽链释放下来,它自己也从mRNA链右端解脱下来,约在同时,在mRNA的左端又挂上一个核糖体,再开始一条新的多肽链的合成。在一个mRNA链上同时有很多核糖体依次滑动,同时合成多条多肽链,这样就大大地提高了蛋白质合成的效率(图6)。
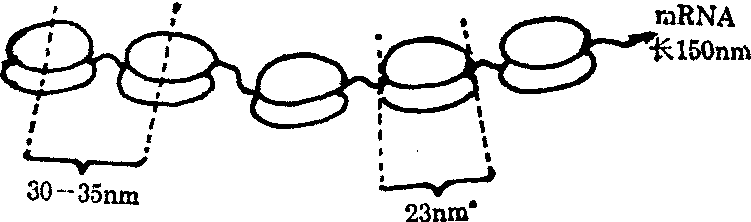
图5 多聚核糖体示意图
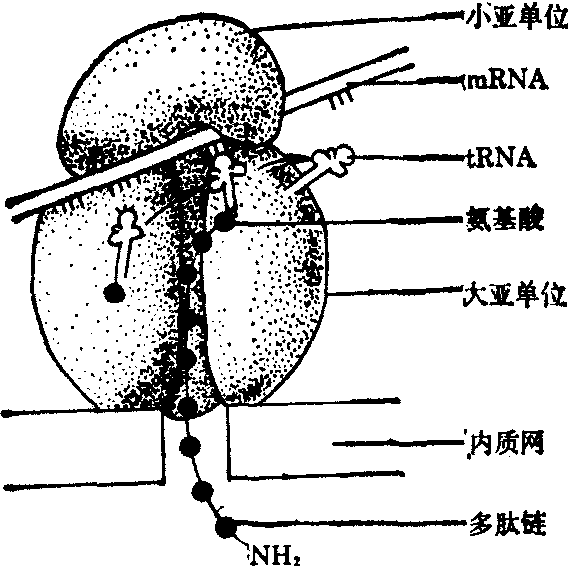
图6 在多肽链合成时核蛋白体与mRNA、tRNA的关系
- 玉花鸟佩是什么意思
- 玉芳是什么意思
- 玉芽是什么意思
- 玉芽开手爪,酥颗点肌肤。是什么意思
- 玉苍是什么意思
- 玉苏子是什么意思
- 玉苏普阿勒克塔格山是什么意思
- 玉苏甫琼买里斯是什么意思
- 玉苏音古勒其是什么意思
- 玉苕是什么意思
- 玉苞是什么意思
- 玉英是什么意思
- 玉英凋落尽,更何人识,野棠如织。是什么意思
- 玉英宝卷是什么意思
- 玉英时共饭,芝草为余拾。是什么意思
- 玉茎是什么意思
- 玉茎下疳方是什么意思
- 玉茎不痿方是什么意思
- 玉茎作肿方是什么意思
- 玉茎强中方是什么意思
- 玉茎湿痒方是什么意思
- 玉茎生疮方是什么意思
- 玉茎疮溃方是什么意思
- 玉茗是什么意思
- 玉茗堂是什么意思
- 玉茗堂主人是什么意思
- 玉茗堂全集是什么意思
- 玉茗堂四梦是什么意思
- 玉茗堂四梦全谱是什么意思
- 玉茗堂四种传奇是什么意思
- 玉茗堂文是什么意思
- 玉茗堂文集是什么意思
- 玉茗堂派是什么意思
- 玉茗堂选集是什么意思
- 玉茗堂遗址是什么意思
- 玉茗堂集是什么意思
- 玉茗花是什么意思
- 玉茧是什么意思
- 玉茭是什么意思
- 玉茭则是什么意思
- 玉茭圪低是什么意思
- 玉茭圪党是什么意思
- 玉茭圪揽是什么意思
- 玉茭地里种豆角——捎办是什么意思
- 玉茭子是什么意思
- 玉茭子圪榄是什么意思
- 玉茭子毛毛是什么意思
- 玉茭子皮是什么意思
- 玉茭子窝窝是什么意思
- 玉茭子糊糊是什么意思
- 玉茭子花儿是什么意思
- 玉茭子茬是什么意思
- 玉茭子面是什么意思
- 玉茭子面糊糊是什么意思
- 玉茭捞捞饭各是各是什么意思
- 玉茭棒子是什么意思
- 玉茭的是什么意思
- 玉茭花是什么意思
- 玉茭茭是什么意思
- 玉茭草是什么意思