癌变学说
正常细胞受到各种致癌因素作用后转变为癌细胞的机制了解得还不多。过去曾经有慢性刺激说和胚芽残存说等。主张慢性刺激说者认为癌瘤都是慢性刺激引起的,例如胃癌与慢性胃溃疡,宫颈癌与慢性宫颈炎、宫颈糜烂,胆囊癌与胆石症等似乎都有密切的因果关系。主张胚芽残存说者认为肿瘤发生于胚胎发育早期遗留下来的胚胎细胞,例如腮弓残余,肾上腺或卵巢内的胚胎组织等均可形成肿瘤。这些学说均未接触到癌变的实质。现代关于癌变的理论基本上可分为两个学派,一派强调癌变是由于DNA (基因)本身的改变,另一派则强调癌变是DNA的表达也即基因的调控发生了改变。前者可称为基因学派,后者可称为基因表达失调学派。
体细胞突变说为基因学派的代表学说,最初由Boveri(1902)提出,认为恶性肿瘤细胞起源于一个染色体含量不平衡的细胞。以后Bauer(1928)系统地提出肿瘤的体细胞突变理论,设想正常细胞的基因发生突变而导致癌瘤。近代有些研究支持这种理论。有人证明动物和人的许多肿瘤是克隆性的,也即来源于单个细胞。对于人的肿瘤,大多数证据来自对黑人妇女长肿瘤时与X染色体连锁的葡萄糖-6-磷酸脱氢酶(G6PD)基因表达的研究。一个妇女的性染色体,一条来自母方,一条来自父方,在胚胎发育过程中有一条染色体随机灭活而形成X染色体,因此成年女性的体细胞中一半具备来自母方的有活性的性染色体,另一半具备来自父方的有活性的性染色体,从而是杂合子。如果发生的肿瘤起源于一个细胞,G6PD的电泳图应为纯合子型,否则为杂合子型。研究结果证明,绝大多数肿瘤起源于单个细胞,因而支持了体细胞突变说。其次,许多化学、物理和生物致癌因素均为诱变剂。过去曾有人强调化学致癌物无诱变作用,但自60年代以来,已证明大多数致癌物均可致突变,仅少数致突变物质不能致癌。过去认为一些致癌物无诱变作用,实际上是前致癌物,经过活化成终致癌物后均可表现出诱变作用。此外也发现化学致癌物虽与DNA、RNA及蛋白质均能结合,但与DNA结合较久,与DNA结合的能力与它的致癌力基本一致。X线、γ线、中子流等均有诱变作用,早已得到公认。至于DNA致瘤病毒和RNA致瘤病毒引起的肿瘤,都可以用致瘤病毒向宿主细胞引进新的遗传信息,从而引起突变来解释。DNA致瘤病毒进入细胞后病毒DNA即可整合到细胞核内的DNA中; RNA致瘤病毒可通过反转录酶,以病毒RNA为模板,形成互补DNA,再整合至细胞的DNA(前病毒学说)。原生病毒学说则认为细胞DNA上有一个原生病毒区,以这段DNA作模板合成的RNA可携带反转录酶到另一个细胞,合成新的DNA,再整合到第二个细胞的原生病毒区或另一个区,反复进行即可形成RNA致瘤病毒。所以原生病毒说认为RNA致瘤病毒是由原生病毒进化来的,而原生病毒的基因是从细胞DNA中的一部分转录而成。
晚近Knudson根据儿童期一些肿瘤的流行病学研究,提出了二次突变说,认为一个正常细胞经过二次以上的突变就会恶变成癌瘤细胞,以后这个恶性细胞在一定条件下形成增殖优势,就将成为癌。家族性视网膜母细胞瘤多为双侧,早发,属显性遗传,是因为第一次突变发生于生殖细胞,出生后体细胞中再发生第二次突变。非家族性视网膜母细胞瘤多为单侧,晚发,较少见,二次突变均发生于体细胞,而体细胞连续二次突变的概率就小得多。
分化异常说是基因表达失调学派的代表学说。这一学说认为细胞的分化过程实质上是基因的表达、阻遏和调节的结果,癌细胞有分化异常的表现,不仅有形态学的表现。还可出现胚胎抗原和同功酶,某些肿瘤可产生激素而引起异位激素综合征。这些现象都是一些基因去阻遏后获得表达而产生的,因此癌细胞很可能不是基因突变的结果,而是基因的表达、阻遏和调节的异常。下列实验有利于说明这一观点: 有人用疱疹病毒在三倍体豹蛙诱发肾腺癌(Lucke腺癌),所产生的癌细胞核具一套三倍体染色体,然后分离出癌细胞核,移植于已被激活但摘除胞核的正常蛙卵内,结果可得外观正常和功能完备的三倍体蝌蚪。这一事实说明,不管是正常的还是癌瘤的细胞核,表现的行为主要由该细胞核所在的胞质环境所决定,尽管移植的癌细胞核带有DNA致瘤病毒的信息,但处于阻遏状态,不表达出来。哺乳动物的肿瘤也有类似的例子。Mintz等(1975)将黑小鼠的畸胎癌细胞注入棕小鼠产生的胚泡中,再将胚泡移植于假孕母鼠子宫内,结果分娩出有黑色条纹的正常小鼠。这也说明恶性状态及其逆转并非突变。Mintz认为畸胎癌起源于组织结构打乱的胚胎,由此推测其他一些干细胞也可通过组织紊乱、基因表达发生改变而产生恶性状态。
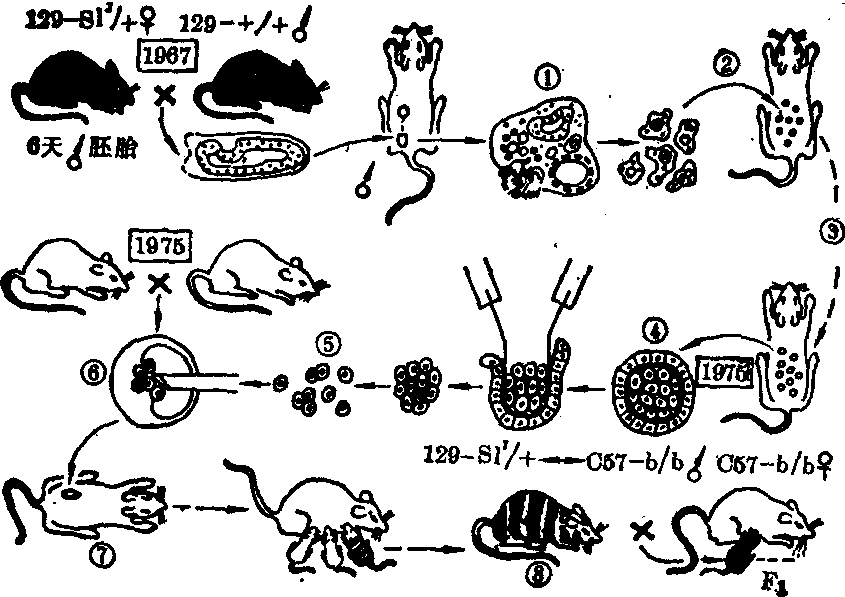
畸胎癌细胞注入胚泡后移植于寄养母鼠可娩出嵌合体
小鼠(引自Mintz,1975)
另外有些学说主张正常细胞内都有肿瘤基因或肿瘤DNA,但受阻遏而不表达,只是在内外刺激因子作用下,使这种基因表达出来而产生肿瘤。例如瘤基因学说(Huebner和Todaro,1969),根据大多数或全部脊椎动物的细胞内均含有C型RNA致瘤病毒的基因组,认为动物在亿万年的进化过程中早就被感染,通过垂直传播,C型RNA致瘤病毒的基因组由亲代传至子代。病毒基因组包括有能引起癌瘤转化的一部分瘤基因,正常时瘤基因处于抑制状态,但可由化学致癌物、电离辐射或其他肿瘤病毒(如DNA致瘤病毒或RNA致瘤病毒)去阻遏,通过转录、翻译形成一种转化蛋白质而使细胞转化为癌瘤细胞。但有人用核酸分子杂交法证明,人的白血病细胞含有与鼠白血病RNA致瘤病毒相应的遗传信息,而正常细胞中并不含有白血病细胞的致瘤病毒信息,以同法检查同卵双生子中的白血病患者的白细胞为阳性,而其健康同胞的白细胞为阴性。这些结果均不支持瘤基因学说。
癌变的基因学说和基因表达失调学说都有一定的事实根据,由于对真核细胞基因组的构成及其调控仍然很不了解,因此这些学说也只指出了癌变机制的某一方面,细节都不清楚。众所周知,胞质和胞核处于复杂的对立统一的相互关系中,细胞的遗传特性不仅取决于胞核,胞质也有相当大影响,基因的表达受到许多胞质因子的调控。Rous肉瘤病毒温度敏感突变种感染鸡成纤维细胞时,这些细胞在36℃(容许温度)下可转化为肿瘤细胞,但同样受感染的细胞在41℃(不容许温度)时表现正常。这一事实启示肿瘤细胞既有遗传信息的改变,又说明与恶性有关的基因的表达或阻遏取决于一定条件,而且是可以接受人工控制的。
近来有人综合基因改变和基因表达失调在癌变中的作用,提出了癌变是DNA损伤,去阻遏和去分化以及选择的结果(DDS学说),即认为各种致癌因素均可引起DNA损伤,但异常的基因受到细胞内各种因子的阻遏,不一定表达出来,助癌因子可发挥去阻遏作用,去阻遏的同时,胚胎基因也可同时开放,这样的癌细胞再经历体内生物学选择作用,形成增殖能力强、有侵犯性、能逃避免疫监视的克隆,才能长成癌瘤。
- 云表是什么意思
- 云谲波诡是什么意思
- 云谷是什么意思
- 云豆是什么意思
- 云豹是什么意思
- 云贵高原是什么意思
- 云起是什么意思
- 云轩是什么意思
- 云逸是什么意思
- 云遮雾障是什么意思
- 云量是什么意思
- 云锣是什么意思
- 云锦是什么意思
- 云门是什么意思
- 云阳是什么意思
- 云际是什么意思
- 云雀是什么意思
- 云集是什么意思
- 云雨是什么意思
- 云雨巫山是什么意思
- 云雷是什么意思
- 云雾是什么意思
- 云霄是什么意思
- 云霓是什么意思
- 云霞是什么意思
- 云霭是什么意思
- 云顶是什么意思
- 云风是什么意思
- 云飞是什么意思
- 云髻是什么意思
- 云鬓是什么意思
- 云鹏是什么意思
- 云鹤是什么意思
- 云龙是什么意思
- 互是什么意思
- 互为是什么意思
- 互为因果是什么意思
- 互为表里是什么意思
- 互利是什么意思
- 互动是什么意思
- 互助是什么意思
- 互助会是什么意思
- 互助组是什么意思
- 互勉是什么意思
- 互市是什么意思
- 互惠是什么意思
- 互感是什么意思
- 互换是什么意思
- 互换性是什么意思
- 互教互学是什么意思
- 互文是什么意思
- 互斥是什么意思
- 互派是什么意思
- 互济是什么意思
- 互溶是什么意思
- 互爱是什么意思
- 互生是什么意思
- 互用是什么意思
- 互相是什么意思
- 互相呼应是什么意思