生物膜与物质运送
生物膜与物质运送
生物膜是细胞与环境之间物质通透的屏障,它具有高度的选择性。生物膜对物质的运送方式多种多样,主要有以下几种。
中介运送 脂质双分子是生物膜结构的基本骨架,按理,不带电荷的脂溶性物质容易通透,而带有电荷或极性基团的亲水物质则难于自由出入。但实际上一些水溶性小分子(如氨基酸、葡萄糖等)或离子能以很高速率穿越生物膜,而另一些却不能。换言之,通过生物膜进行运送具有高度选择性。这是由于有专一的运送载体、运送体或运送酶系存在。这样的运送过程称之为中介运送或促进运送。除专一性外,它的特点在于:
❶与酶催化反应速率随底物浓度增加而增加相类似,在小于饱和浓度情况下,运送物质的速率也随着其浓度增加而增加;
❷阻碍运送过程的抑制剂具有选择性。
被动运送 根据运送过程自由能变化的情况,基本上可分为两大类:被动运送和主动运送。
物质从浓度较大一侧通过膜运送到浓度较小一侧,犹如溶质通过透析袋那样,它的运送速率依赖于膜两侧运送物质的浓度差及其分子的大小,电荷性质等等。根据热力学第二定律,被动运送过程的自由能减少,熵是增加的。换言之,这是一个不需供给能量的自发过程。红细胞膜第三带蛋白运送阴离子(Cl-、SO-4等)的过程就属于这一类。
主动运送 凡物质逆浓度梯度进行运送的过程称为主动运送。在此过程中自由能是增加的,需要供给能量才能进行运送。主动运送过程的自由能变化可以下式表示:
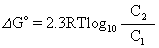
如果运送物质带有电荷,则主动运送时需要逆两个梯度,一是浓度梯度,二是电荷梯度,这两者的总和称为电化学梯度。在这里,运送过程的自由能变化应为:
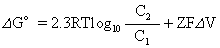
在生物体内,为了进行主动运送,至少必须具有两个体系,一是参与运送的传递体,二是酶或酶系组成的能量供应系统。这二者偶联才能进行主动运送。
载体假说与通道假说 对物质运送通过生物膜的机理一般可分为载体假说和通道假说。
载体假说认为,被运送物质与膜上载体相结合后,通过扩散、旋转或构象变化使被运送的物质从一侧运至另一侧。
通道假说认为,在运送过程中膜上存在着一定的通道。它可能是一种受控的孔道,在运送过程中,被运送物质并不一定要求与通道结构组份进行结合。这种通道也不一定是一种永久性结构。
钠、钾的运送 可兴奋细胞膜(如神经元、心肌和各种肌细胞)存在着受膜电位控制的、分别对钠离子和钾离子可通过的钠通道和钾通道。静息时它们处于‘关’状态,通道关闭; 兴奋时,膜电位去极化,它们转为‘开’状态,通道开放,允许离子通过。除受电压控制的离子通道外,在可兴奋膜上还有一种化学可兴奋的离子通道,其中研究得比较清楚的是神经肌肉接头后膜上的乙酰胆碱受体通道,这种离子通道在乙酰胆碱作用下开放。
细胞运送Na、K的体系除Na、K通道外还有细胞质膜线粒体内膜等的Na+-Ca2+交换体,细胞质膜Na+、K+泵(或称Na+、K+-ATP酶)。
钠通道 对乌贼轴突、青蛙神经、海鞘卵细胞膜的钠通道已经进行了不少研究。近年来,对离子通道的生化研究也取得了不小的进展。从大鼠肌纤维膜可分离、纯化一种钠通道蛋白质,如果将它嵌入脂体就能增加22Na+的流入。从哺乳动物脑分离的钠通道蛋白质含有分子量分别为270,000和37,000的两种多肽。对乙酰胆碱受体的分离、纯化、组成、结构及其运送离子的功能的研究所获得的成果尤为明显(参见“生物膜与信息传递”)。
钠、钾泵 动物细胞Na+的浓度一般都比周围环境低,K+浓度比周围环境高,很多细菌和植物细胞也有类似情况。这种离子梯度的形成是主动运送的结果,这个运送系统称为钠、钾泵。细胞内、外保持Na+、K+ 的浓度梯度具有很重要的生理意义,它是神经、肌肉膜可兴奋的基础,也是某些细胞中氨基酸、葡萄糖运送得以进行的动力。
1. 钠、钾泵与Na+、K+-ATP酶: 一系列的研究结果表明,Na+、K+-ATP酶是钠、钾泵的主要组成部分。主要证据有:
❶凡具有主动运送Na+、K+功能的细胞膜都表现Na+、K+-ATP酶活性,二者的活性是平行的。
❷钠、钾泵与Na+、K+-ATP酶在膜上有相同的取向,用红细胞空泡作实验材料,控制其中所含的溶质浓度进行研究表明,Na+必须在膜内侧,K+必须在膜外侧才能激活Na+、K+-ATP酶活性和钠、钾泵功能,而且只有细胞内的ATP才能被钠、钾泵和Na+、K+-ATP酶所利用,抑制剂乌本苷只有在细胞外才能对两者起抑制作用(图1)。
❸钠、钾泵和Na+、K+-ATP酶活性受乌本苷抑制时,半抑制所需浓度也相同。
❹将提纯的Na+、K+-A-TP酶重组到脂质体上,成功地显示钠、钾泵的功能。
2. Na+、K+-ATP酶的性质和作用机理: Na+、K+-ATP酶是跨膜分布的内在蛋白,提纯的酶含30%左右的脂,去脂后酶即丧失活性,加回脂后酶活性可重新呈现,其中以磷脂酰丝氨酸和二磷脂酰甘油最为有效。
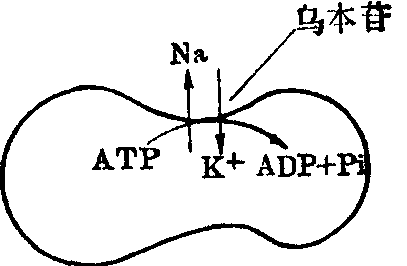
图1 红细胞Na+、k+的主动运送示意图
Na+、K+-ATP酶分子量为250,000~300,000,由二个亚基组成,大的亚基分子量约100,000,拥有催化活性部位,并含乌本苷的结合位点。小亚基分子量约50,000,在膜外侧表面连接有糖的残基。Na+、K+-ATP酶在膜上可能以二聚体存在。
利用γ-32P标记的ATP进行研究表明,Na+、K+-ATP酶在反应过程中至少可分为磷酸化和去磷酸化两步反应:
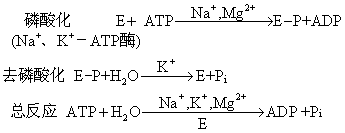
红细胞膜的Na+、K+-ATP酶每水解一分子ATP能运入2K+和运出3Na+。
钙的运送 Ca2+不仅参与细胞的兴奋过程,而且还调节细胞的很多功能,例如,内吞和外排,激素、神经递质的分泌释放,离子通道的开、闭等等。此外,Ca2+对细胞内一些酶反应的调节以及肌纤维的收缩,微管、微丝的聚合、解聚,纤毛、鞭毛的运动都起着重要的作用。一般讲,细胞质的Ca2+浓度很低(10-8~10-7mol/L),而细胞外则高达10-3mol/L。维持这样的浓度梯度可以通过Ca2+与可溶性蛋白质(如拟清蛋白)相结合,但主要还在于细胞质膜和细胞内膜系统运送Ca2+的体系参与作用。真核细胞的质膜、线粒体膜和内质网、肌浆网膜都含Ca2+的运
送系统。它们是:兴奋性细胞质膜上的受电压控制的Ca2+
通道,质膜的Ca2+-Na+交换体和Ca2+泵,内质网膜或肌浆网膜上的Ca2+泵和Ca2+释放系统,以及线粒体内膜的Ca2+-Na+ 交换体和受H+电化学梯度驱动的电泳单向运送体。
钙调蛋白(CaM) 对调节细胞内Ca2+ 浓度具有重要作用。它由149个氨基酸组成,分子量为16,700,每个分子能结合4个Ca2+。在胞内Ca2+极低浓度下,CaM主要以不与Ca2+结合的非活性状态存在,不能激活Ca2+-ATP酶,酶对Ca2+的亲和力也很低。如果胞内Ca2+浓度达到10-6~10-5mol/L,CaM与Ca2+形成复合物,它可与Ca2+-ATP酶结合,并提高酶对Ca2+的亲和力,同时酶活性增加6~7倍,使Ca2+的主动运送大大加强,从而使胞内Ca2+浓度又达到原有稳态水平。
钙泵(Ca2+-ATP酶) 在上述几种Ca2+的运送体系中,对内质网膜或肌浆网膜的Ca2+泵有较多的了解。肌浆网膜是研究Ca2+泵结构与功能的常用实验材料。其中特别是骨胳肌含有丰富的肌浆网膜,Ca2+泵约占膜蛋白的70%。在神经传导引起肌肉收缩的过程中,神经传导先导致肌浆网膜的兴奋,引起Ca2+大量释放,从而肌肉收缩。当肌肉松弛,又将肌纤维之间的大量Ca2 +运送入肌浆网,膜内外Ca2+浓度比可高达10 000倍,这是一种主动运送,由肌浆网膜的Ca2+泵来完成。Ca2+泵具有Ca2+激活ATP酶 (Ca2+-ATP酶) 活性,Ca2+泵主动运送Ca2+是由ATP水解提供的能量来驱动的。Ca2+-ATP酶对Ca2+的亲和力很高,Km约为10-7mol/L。Ca2+泵具有效率高、容量大的运送功能在≈20~100毫秒内每克肌肉能将0.2μg 分子Ca2+进行运送。每水解一分子ATP可运送二分子Ca2+。
Ca2+泵在运送Ca2+的过程中也经历磷酸化和去磷酸化的循环反应:
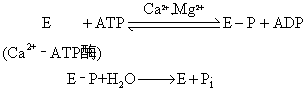
从肌浆网膜分离提纯的Ca2+-ATP酶分子量约为110,000,在总共1015个氨基酸残基中,80%以上的一级结构已经阐明。Ca2+-ATP酶用胰蛋白酶处理,先分解成两个大小基本相同的肽段(分子量分别为57,000,55,000)。分子量为57,000的肽段可继续分解成分子量为24,000和33,000的两个小肽段,前者具有运送Ca2+的功能,后者呈现水解ATP的活性。Ca+2-ATP酶是一个跨膜分布的膜结合酶,它在膜上可能以四聚体形式存在。
提纯的Ca2+-ATP酶已在脂质体上重组成功,当加入ATP时表现出Ca2+泵的功能,将Ca2+主动运入脂质体内。
阴离子运送 通过膜也需一定的载体,例如红细胞膜的第三带蛋白就是一种运送阴离子(包括一价和二价、有机和无机阴离子)的载体。第三带蛋白占红细胞膜蛋白的25%以上。它是一种糖蛋白,其亚基分子量为93,000~95,000。氨基酸组成中37.5%是疏水氨基酸,N端是乙酰甲硫氨酸,C端是赖氨酸。糖通过门冬酰胺与蛋白质连接,含有29个残基,它们主要是半乳糖、N-乙酰葡萄糖胺,也有少量甘露糖、岩藻糖与唾液酸。
第三带蛋白在膜内以二聚体形式存在,然而它的作用单位都是单体。
第三带蛋白按其分布情况可分成三个部分,即A肽(细胞质一侧)、B肽(膜内部分)及C肽 (细胞外侧)。C肽除与B肽连接外也与膜紧密结合,它对阴离子运送有比较重要的作用。
协同运送 有些细胞(如小肠上皮细胞)通过主动运送摄入葡萄糖或氨基酸时,周围介质中必须有很高浓度的Na+存在,在进行上述主动运送过程中都伴随有Na+进入细胞,这与细胞膜上钠、钾泵的作用方向是相反的。协同运送假说认为,以葡萄糖主动运送为例,细胞膜上的钠、钾泵利用ATP为能源使细胞内外产生Na+的浓度梯度,这是将葡萄糖输入的动力。由于膜外侧的Na+浓度较高,因而它流向细胞内是一个不需能的自发过程。葡萄糖就利用Na+浓度梯度,通过专一运送载体伴随Na+一起运送进入细胞。当细胞内Na+浓度因回流而增加时,钠、钾泵又再次运转将Na+泵出。这样,葡萄糖并不直接利用ATP而利用钠、钾泵产生的Na+浓度梯度进行协同运送,不断进入细胞。
基团运送 一般运送物质在通过膜时并不进行化学修饰,但有些糖在通过细菌膜时需进行磷酸化反应,因而加入一个磷酸基团,这称为基团运送。以葡萄糖为例,它在运送进入细菌过程中转变成葡糖-6-磷酸。由于细菌质膜对糖的磷酸酯是不通透的,这样就在细胞内进行积聚。糖要进行磷酸化反应也需要供给能量,所以这种运送也属于主动运送。但在这里,不是利用ATP作为磷酸基的供体而系利用磷酸烯醇丙酮酸,后者经磷酸烯醇丙酮酸转磷酸化酶系统催化转移磷酸基。这个系统的总反应是:
糖外侧+磷酸烯醇丙酮酸→糖-磷酸内侧+丙酮酸
内吞作用 也是跨细胞质膜进行物质运送的一种方式。它是通过细胞质膜内陷形成囊泡,将外界物质裹进并输入细胞内的一个过程。按输入物的大小,物质状态以及特异性程度等,一般将内吞分为吞噬、胞饮以及经受体的内吞作用等三种。
吞噬作用 细胞内吞较大的固体颗粒或分子复合物(直径可达几微米),称为吞噬作用。例如,原生动物摄取细菌和其他食物颗粒,高等动物免疫系统的巨噬细胞裹吞入侵的细菌等。吞噬可分吸附和吞进两个相对独立的过程。总的来讲吞噬是一个需能的主动运送过程,但吸附并不需要能量。一般认为,被吞噬的颗粒在细胞表面的吸附并不具有明显的专一性。
胞饮作用 将周围的微滴状液体(微滴的直径一般小于1μm)吞入胞内,称为胞饮。被吞进的微滴液体经常含有离子或小分子。胞饮作用也不具有明显的专一性。绝大多数细胞都能胞饮,即使是哺乳动物的正常成熟红细胞,只要条件合适也能进行胞饮作用。一个巨噬细胞1小时饮入的液体可达细胞体积的20~30%。
经受体的内吞作用 是专一性很强的内吞作用,被内吞物(称为配体,它们或是蛋白质或是小分子)与细胞表面的专一受体相结合。结合后随即引发细胞膜的内陷,形成的囊泡将配体裹入并输进细胞内。
铁传递蛋白是肠、肝细胞摄取铁的一个重要载体。载有二个铁离子的铁传递蛋白与细胞表面的专一受体相结合,随后受体、铁传递蛋白复合物通过内吞进入细胞。有些激素(如胰岛素)也是与受体结合后进行内吞而将信息传给细胞的。胰岛素分子与靶细胞的专一受体结合后即引发内吞作用。胰岛素-受体复合物内吞进入细胞后引起一系列的代谢反应,产生生理效果。
内吞后的变化 广义的内吞过程不仅指被吞物质如何被细胞质膜内陷形成的囊泡所裹进并输入细胞内部,还包括含有内吞物的囊泡进入细胞内以后如何进一步变化的过程。无论哪一种内吞类型(吞噬、胞饮或经受体进行内吞),这些囊泡可能会经历膜表面的变化、相互融合、与溶酶体融合等步骤,最后使被吞进物质从囊泡内释放或被溶酶体内含的水解酶所水解。对每一种内吞物来说,不一定都经历上述各步骤,每一步骤的具体细节也可能有差异,但是整个内吞过程是一个连续的、快速的生物膜重排和融合过程却是共同的。
外排作用 一个与内吞作用相反的运送过程。细胞内物质先被囊泡裹入形成分泌泡,之后与细胞质膜接触、融合并外排至胞外。外排作用在细胞分泌时经常发生。许多因素能引起细胞分泌,其中至少有些是改变了细胞膜对Ca2+的通透性使之进入细胞内而引起的。例如,神经因素能引起腮腺和肾上腺髓质细胞分泌,血浆中的葡萄糖促进胰岛细胞的分泌,都是通过细胞膜的去极化,让钙离子进入细胞而引起的。在外排过程中,胞内含外排物质的分泌囊泡往往先与细胞质膜发生融合,这也需Ca2+的参与。例如,当神经脉冲传至突触前膜时,由于膜的去极化,让Ca2+进入从而使突触细胞质内大量的储存泡与突触前膜相融合而将所含的乙酰胆碱释放至突触隙内,之后乙酰胆碱与突触后膜上的受体相结合,使神经兴奋继续传递下去。又如,海胆卵受精时,皮质颗粒通过与细胞质膜的融合而将其内含物排至细胞外。这一外排过程也需Ca2+参与。
在外排过程中,当分泌囊泡与细胞膜相互融合并将内含物排出以后,分泌囊泡膜的组份就成为细胞膜的一部分。这部分也可能降解或‘回流’而被重新利用。
内吞和外排作用与其他主动运送一样也需要能量供应。例如,如果氧化磷酸化作用被抑制,那么巨噬细胞的吞噬作用就会被阻止。同样,如果分泌细胞中的ATP合成被阻遏,外排作用也不能继续进行。
蛋白质运送通过膜
1. 信号假说: 1972年Milstein等在研究免疫球蛋白轻链合成时,发现在它的N-末端含有~15个氨基酸的额外肽段。他们认为,这段氨基酸序列对在多核糖体上合成的分泌蛋白穿越膜时可能具有信号作用。在此基础上提出了‘信号假说’,认为像分泌蛋白合成后需通过膜的一类蛋白质(甚至可能包括一般的膜蛋白),在编码它们合成的结构基因中含有一种信号序列。通过翻译,信号序列形成信号肽。信号肽与粗型的内质网膜的识别序列相结合,从而引导合成的多肽能穿越膜。信号肽一旦穿越膜即被多肽酶水解,而蛋白质的合成仍继续进行。这一假说认为, 这类蛋白质合成的翻译过程与它们穿越膜的过程是同时进行的(图2)。
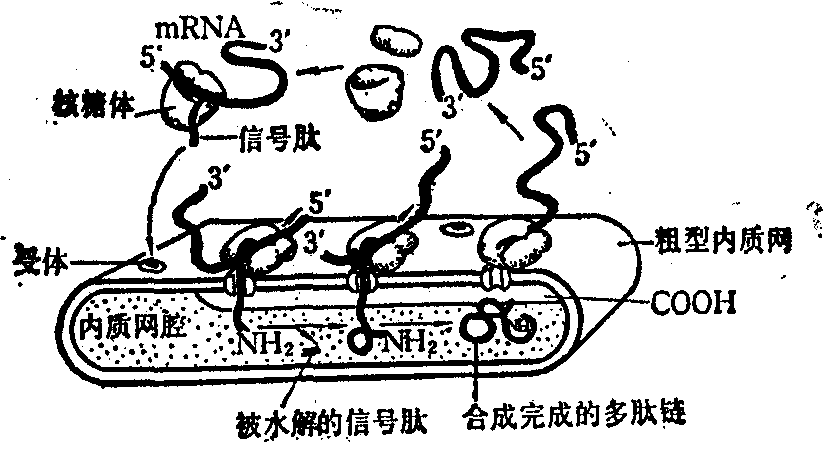
图2 信号假说示意图(自B.Alberts)
2. 蛋白质运送通过线粒体膜:线粒体内含的蛋白质绝大多数系由细胞核的DNA提供信息,并在细胞质中的核糖体内合成,之后再通过线粒体膜运送至内部进行更新或组装。近年来,很多研究结果表明,细胞质合成的蛋白质插入内膜(如H+-ATP酶或细胞色素氧化酶的亚单位、D-β-羟丁酸脱氢酶等) 或通过内膜进入基质(如L-谷氨酸脱氢酶、苹果酸脱氢酶等)或进入内、外膜的间隙(如酵母线粒体的细胞色素b2)的过程大多具有如下的特点:
❶通过线粒体内膜的多肽或蛋白质起始都是先合成较大的前体,通过膜时进行酶促水解形成’成熟’的形式。与信号假说中正在合成的蛋白质通过膜的情况不同,新生多肽或蛋白质通过线粒体内膜时,合成过程已经终止,因而它们通过膜的过程并不伴随蛋白质合成的翻译过程。
❷蛋白质通过线粒体内膜是需能的主动运送过程。
❸蛋白质通过线粒体内膜时可能都有相应的受体与之相结合。
多肽进入线粒体内、外膜间隙的过程比运送至内膜或基质的过程更为复杂。例如,酵母线粒体细胞色素b2,它在细胞质内合成的首先是一个分子量为68,000的前体,与线粒体内膜结合并穿越之后被基质内的蛋白水解酶裂解成一种中间形式(分子量为65,000),之后它又返越内膜,当大部分进入膜间隙时又遭第二种蛋白酶作用才变成‘成熟’形式(分子量为58,000),并释放至膜间隙内。但是,并非所有多肽或蛋白质输入膜间隙都分两步进行。而且,像细胞色素c的合成也没有发现有较大的前体存在,它先以不含血红素的脱辅基蛋白的形式运送入内,然后在线粒体内加上血红素才转变为细胞色素c。
多肽运送到线粒体外膜的过程与上述情况不同,它既不是一个需能的主动运送过程,也不需要经过酶水解的步骤。
☚ 生物膜与能量转换 生物膜与信息传递 ☛
- 莫雄回忆录是什么意思
- 莫雷是什么意思
- 莫雷亚斯是什么意思
- 莫雷利是什么意思
- 莫雷尔法通过是什么意思
- 莫雷斯·安德拉德是什么意思
- 莫雷洛斯是什么意思
- 莫雷洛斯是什么意思
- 莫雷蒂是什么意思
- 莫雷蒂是什么意思
- 莫雷西嗪是什么意思
- 莫雷西嗪是什么意思
- 莫雷诺是什么意思
- 莫雷诺是什么意思
- 莫雷诺是什么意思
- 莫雷诺是什么意思
- 莫雷诺·希门尼斯是什么意思
- 莫雷诺社交测量法是什么意思
- 莫雷诺-杜兰是什么意思
- 莫霍人是什么意思
- 莫霍钻孔计划是什么意思
- 莫非是什么意思
- 莫非是什么意思
- 莫非仙是什么意思
- 莫非仙是什么意思
- 莫非仙是什么意思
- 莫非麦考酚酯是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫须有是什么意思
- 莫顿是什么意思
- 莫顿,杰是什么意思
- 莫马迪是什么意思
- 莫高是什么意思
- 莫高是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟是什么意思
- 莫高窟千佛洞是什么意思
- 莫高窟唐代壁画是什么意思
- 莫高窟年表是什么意思
- 莫高窟的由来是什么意思
- 莫高窟窟前殿堂遗址是什么意思
- 莫高窟第45窟菩萨象是什么意思
- 莫高窟藻井图案是什么意思
- 莫鲁克是什么意思
- 莫鲁瓦是什么意思
- 莫鼎榕是什么意思
- 莰是什么意思
- 莱是什么意思
- 莱是什么意思
- 莱是什么意思