信使核糖核酸(mRNA)
信使核糖核酸在细胞中的含量约占总RNA的5%以下,其量虽少,但在蛋白质生物合成中却起着关键性的模板作用。它在细胞核或拟核区域由DNA转录生成,并经加工处理后,进入细胞质可以自由存在,但主要是与多个核蛋白体结合成串珠样的多核蛋白体的形式而存在。mRNA的种类繁多,从理论上讲,真核细胞中含多少种蛋白质或肽链,就必然含多少种mRNA,但在原核细胞中的种类却要少得多。各种mRNA的分子大小颇不一致,相差悬殊,一般由数百个至数千个核苷酸组成,沉降常数为8S至30S,分子量约为20万至200万道尔顿,但个别mRNA可能更大或更小,例如蛋丝纤维蛋白mRNA的分子量可大至600万以上。mRNA一般都不稳定,代谢活跃,更新迅速,寿命较短,原核mRNA的此种现象较真核mRNA尤为显著,前者半寿期只有数分钟,但后者的也不过数小时或数天。mRNA的基本组成单位也都是腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)四种碱基的相应核苷酸,分子中一般不含稀有核苷酸(原核mRNA)或含量极少(真核mRNA);这些核苷酸的含量没有一定比例,它们按照一定的排列顺序,通过3′,5′磷酸二酯键彼此相连而形成一条很长的多核苷酸链,链本身可有多处折叠造成发夹式结构,并形成许多小双螺旋。但不同mRNA所含发夹式结构的数目、长短、相对位置等各不相同,即其二级结构没有共同的形式和规律。一级结构与功能关系 原核生物mRNA与真核生物mRNA的分子结构组成具有很大差别。绝大多数原核细胞的mRNA,包括嗜菌体RNA,都是多顺反子型信使,即一个原核mRNA分子中带有几种蛋白质的信息,可作为合成几种蛋白质的模板,它们都是能为几种蛋白质编码的操纵子的转录物或自身复制物。例如大肠杆菌中由 糖操纵子转录来的mRNA带有能合成三种蛋白质的信息,由色氨酸操纵子转录来的mRNA带有能合成五种蛋白质的信息; 又如由噬菌体MS2自身复制的mRNA也带有合成三种蛋白质的信息; 等等。与此相反,所有已知真核细胞中的mRNA都是单顺反子型信使,即一分子真核mRNA只携带一种蛋白质的信息,只能作为合成一种蛋白质的模板。此外,真核细胞mRNA和原核细胞mRNA的生成方式也不一样,前者都是由其前体即非均一性核RNA(HnRNA)经加工处理后转变而来,而后者则没有前体,一般是边直接转录生成,边直接进行翻译,了解比较清楚的多顺反子型mRNA是一些RNA噬菌体的RNA,它们既是复制自己的基因,又是合成其所含蛋白质的模板,例如大肠杆菌噬菌体R17、MS2、f2、QB等的RNA都是由三种顺反子组成,带有能合成A蛋白(成熟蛋白)、外壳蛋白及复制酶(RNA聚合酶)三种蛋白质的信息。实际上每种这类顺反子都是一种特异蛋白质的编码区,也就是一个特异的翻译区,各个翻译区之间借一段无编码及翻译功能的核苷酸序列相连。此外在5′末端及3′末端也各有一个非翻译区,而且5′端的第一个核苷酸一般都是三磷酸鸟苷(pppG),3′端的最后一个核苷酸多为腺苷酸,而且每个顺反子与核蛋白体都有一个独立的结合部位(图1)。至于以单顺反子形式存在的真核细胞mRNA分子,实际上是由三部分组成(图2),位于分子中间的主体部分多数为由几百个至几千个核苷酸构成的编码区,也就是带有某种特异蛋白质信息的翻译区; 另两部分是连在翻译区两边的无编码功能的5′非翻译区及3′非翻译区,此两区段的中心往往各含一个稀有核苷m6A,而且前者的5′末端都是含1~3个稀有核苷的特殊结构,后者的3′末端都是一段完全由腺苷酸聚合成的聚A序列。
糖操纵子转录来的mRNA带有能合成三种蛋白质的信息,由色氨酸操纵子转录来的mRNA带有能合成五种蛋白质的信息; 又如由噬菌体MS2自身复制的mRNA也带有合成三种蛋白质的信息; 等等。与此相反,所有已知真核细胞中的mRNA都是单顺反子型信使,即一分子真核mRNA只携带一种蛋白质的信息,只能作为合成一种蛋白质的模板。此外,真核细胞mRNA和原核细胞mRNA的生成方式也不一样,前者都是由其前体即非均一性核RNA(HnRNA)经加工处理后转变而来,而后者则没有前体,一般是边直接转录生成,边直接进行翻译,了解比较清楚的多顺反子型mRNA是一些RNA噬菌体的RNA,它们既是复制自己的基因,又是合成其所含蛋白质的模板,例如大肠杆菌噬菌体R17、MS2、f2、QB等的RNA都是由三种顺反子组成,带有能合成A蛋白(成熟蛋白)、外壳蛋白及复制酶(RNA聚合酶)三种蛋白质的信息。实际上每种这类顺反子都是一种特异蛋白质的编码区,也就是一个特异的翻译区,各个翻译区之间借一段无编码及翻译功能的核苷酸序列相连。此外在5′末端及3′末端也各有一个非翻译区,而且5′端的第一个核苷酸一般都是三磷酸鸟苷(pppG),3′端的最后一个核苷酸多为腺苷酸,而且每个顺反子与核蛋白体都有一个独立的结合部位(图1)。至于以单顺反子形式存在的真核细胞mRNA分子,实际上是由三部分组成(图2),位于分子中间的主体部分多数为由几百个至几千个核苷酸构成的编码区,也就是带有某种特异蛋白质信息的翻译区; 另两部分是连在翻译区两边的无编码功能的5′非翻译区及3′非翻译区,此两区段的中心往往各含一个稀有核苷m6A,而且前者的5′末端都是含1~3个稀有核苷的特殊结构,后者的3′末端都是一段完全由腺苷酸聚合成的聚A序列。
翻译区及其功能 无论是真核mRNA或原核mRNA的翻译区在蛋白质生物合成中是真正起模板作用的部位,通过转录过程它们由DNA接受特异的遗传信息,再通过翻译作用将此遗传信息转送给蛋白质,从而决定蛋白质的结构和功能。mRNA翻译区携带的遗传信息是以其分子中核苷酸的排列顺序来体现的,具体说就是在其分子中每三个核苷酸组成一组而成为一个三联体,可视为一个信息单位,通称一个密码子,由于mRNA翻译区系由四种不同的许多个核苷酸分子组成,那么这些核苷酸必可排列出43=64种不同的三联体或密码子。六十年代初期以来通过大量实验,已经完全确定了哪三个核苷酸组成的密码子相当于哪种氨基酸,并编出一套适用于一切生物体的遗传密码表(见“遗传密码”条)。64个密码子中除UAA、UAG和UGA三个外,其他每个密码子都代表一种氨基酸,但用以合成蛋白质的氨基酸一般只有二十种,显然一种氨基酸可被几种密码子 (1~6种) 所代表,这样的密码子常被称为同义密码子或简并密码子,其特点是它们的前两个核苷酸都相同,而第三个则不同,UAA、UAG及UGA三个密码子都不代表任何氨基酸,常被称为无意义密码子,但实际上它们在终止蛋白质合成的作用上具有决定性意义,故被称为终止密码子。它们都位于mRNA非翻译区的起始部位。在原核mRNA中,AUG和GUG不仅分别代表蛋氨酸和缬氨酸,而且还是蛋白质生物合成的起动信号,称为起动密码子,它们均位于mRNA翻译区的起始部位。起动密码子中以AUG为主,GUG为辅,二者的机率一般为18:1,当它们作起动密码子使用时,就不再分别代表蛋氨酸和缬氨酸了,而是统统代表甲酰蛋氨酸。在真核mRNA中只能以AUG为起动密码子,GUG则不行。应当指出,并非处于mRNA分子中任何部位的AUG和GUG都可作起动密码子,只有那些位于mRNA分子的起动部位者才有此作用,也就是说起动密码子的起动作用与其附近其他特定核苷酸的排列顺序和立体结构有关。至于上述三种终止密码子,无论在原核mRNA或真核mRNA中都可通用。有些mRNA翻译区的核苷酸序列已经分析清楚,例如MS2RNA的三个翻译区分别由1179个、390个和1635个核苷酸组成,其序列都已搞清,可分别译出成熟蛋白、外壳蛋白和夏制酶三种蛋白质。又如兔α珠蛋白mRNA和β珠蛋白mRNA都各只含一个翻译区,分别由423个及438个核苷酸组成,可分别译出含141个氨基酸的珠蛋白α链和含146个氨基酸的珠蛋白β链。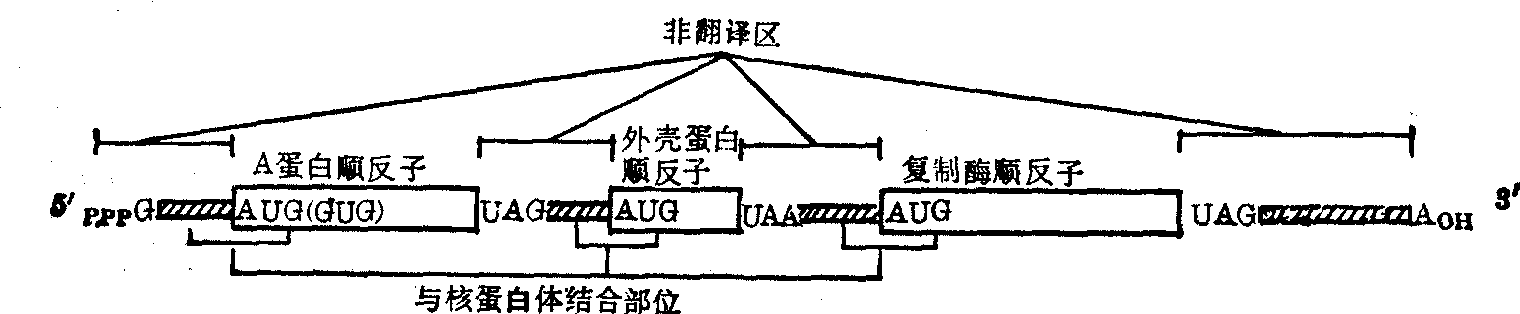
图1 原核mRNA的结构模型(噬菌体MS2的结构)

图2 真核mRNA的结构模型(哺乳动物)

图3 原核mRNA(噬菌体R17A蛋白顺反子)起动部位与核蛋白体30S亚基的16S RNA的碱基互补结合
非翻译区及其功能 mRNA分子中的非翻译区首先是在噬菌体MS2的RNA中发现的,它含有四个非翻译区,即5′非翻译区、3′非翻译区以及位于其翻译区之间的另外两个非翻译区,它们分别由129个、174个、26个及36个核苷酸组成,核苷酸的排列顺序都已全部确定,它们之中有的含有几个AUG或几个GUG,但无一具有起动信息。mRNA非翻译区虽无编码和模板作用,但在蛋白质生物合成中仍然非常重要,就噬菌体MS2或R17者来说,此区至少有下列两种功能:
❶因含有终止信号,可保证蛋白质的翻译过程按时停止。
❷与核蛋白体小亚基结合,构成mRNA的起动部位,其结合机制已被基本确定,主要根据是: 多种原核mRNA及噬菌体RNA的各非翻译区序列中,在距离相邻翻译区起动密码子AUG前的7~10个核苷酸处,都有一段富于嘌呤核苷酸的序列; 而一切细菌核蛋白体小亚基(30S)中16S rRNA的3′末端的12个核苷酸,都是一段富于嘧啶核苷酸的序列,即……AUCACCUCCUUAOH,此二者可以互补结合,至少生成4~7个连续的碱基对(图3)。原核mRNA与核蛋白体小亚基如此相互结合的机制常被称为Shine-Dalgarno定则,但有些事实说明它未必是唯一的结合方式。真核细胞mRNA分子中的非翻译区包括5′端及3′端两部分,其功能可能类似原核mRNA的非翻译区,但与原核mRNA相比,真核mRNA的非翻译区具有两大特点:
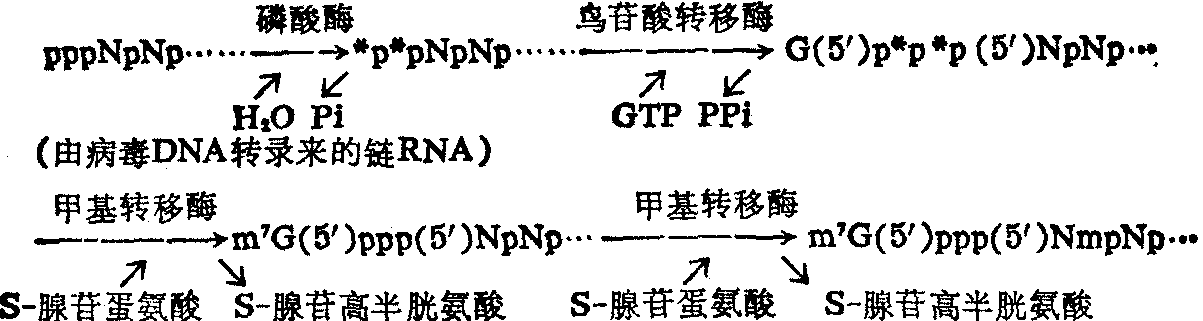
❶5′非翻译区的5′末端为由1~3个稀有核苷酸形成的一个特殊结构区,常称为“帽子”。
❷3′非翻译区的3′端为由30~200个腺苷酸聚合成的所谓 “多聚A尾巴”。根据所含甲基化稀有核苷的数目,5′帽子可有下列三种类型,即m7GpppNp……、m7GpppN1mpNp…和m7GpppN1mpN2mpNp…; 其中7-甲基鸟苷(m7G)中有一个质子解离上去而形成偶极离子; N1m和N2m代表四种2′-o-甲基核苷中的任何一种,即Am、Gm、Cm或Um,但N1m也可能是m6Am; m7G通过三个磷酸基与Np或Nmp以5′,5′键相连,其结构式如下。
5′帽子的存在对于mRNA的翻译作用即促进蛋白质生物合成的能力至关重要,这可能是由于:
❶它能阻止5′外切核酸酶对mRNA的降解作用,使mRNA的稳定性增加。真核mRNA的半寿期何以普遍长于原核mRNA者,可能就是由于此故。
❷它是核蛋白体小亚基识别mRNA的信号并与之结合的部位,因而参与蛋白质的起动作用。真核mRNA的3′聚A尾巴最初是在哺乳动物mRNA中发现的,随后在多种昆虫、植物、酵母等mRNA分子中也都发现。3′聚A中腺苷酸数目不等,一般为30~200个,越高级的真核细胞mRNA所含聚A中的A数越多,反之越少。真核mRNA的前体HnRNA也含3′聚A,但此聚A不是由DNA转录而来。而是HnRNA在转录完成之后于3′末端一个A一个A加上去的,催化此反应的酶为聚A聚合酶,所需原料为ATP,但GTP、CTP、UTP对此酶则有抑制作用。各种真核mRNA的3′非翻译区中,在距离3′聚A约20个核苷酸处有一段共同的序列AAUAAA,可能是聚A聚合酶的识别及结合部位。至于3′聚A的功能虽有多种推测和说法,但实验结果极其混乱矛盾,至今仍无定论,总的估计它可能与维持并稳定mRNA的二级结构有关,从而有利于蛋白质合成的进行。
二级结构 无论真核mRNA或原核mRNA的一级结构都是单股多核苷酸链,但当加热后都可引起变性,表现增色效应;若用核糖核酸酶A或T1处理时,链的某些部位比较敏感,但有些部位则不太敏感,这表示mRNA链可有多处能够自身折叠而形成发夹式结构,并为X线衍射所证实。但不同mRNA所含发夹式结构的数目、长短、相对位置等都不相同,即不同的mRNA具有不同的二级结构形式; 而且即使是同一种mRNA,由于折叠位置的不同也可能形成多种不同形式的二级结构,当然其中必定有的形式更为稳定更合理。例如由3569个核苷酸组成的噬菌体MS2RNA,其全部核苷酸的排列顺序已经测定清楚,仅以其分子中只含390个核苷酸的外壳蛋白顺反子来说,它可以折叠成几种形式的二级结构,分析其中各个发夹式结构的稳定性以及根据它们对核糖核酸酶敏感部位等特点,Fiers等提出此顺反子的二级结构如同一朵花,它主要含有九个长短不等的发夹式结构,而且是从同一圆心向外放射,有如九片花瓣一样。
帽子的生成机制可用下列反应式表示:
- 国际食品法典机构标准制定立法是什么意思
- 国际食品法典机构职责立法是什么意思
- 国际食品法典机构议程拟定立法是什么意思
- 国际食品法典机构费用立法是什么意思
- 国际食品法典机构预算立法是什么意思
- 国际食品法典机构领导成员立法是什么意思
- 国际食品法典范围立法是什么意思
- 国际食品法典附属机构立法是什么意思
- 国际食品添加剂使用定义立法是什么意思
- 国际食品添加剂标准承认立法是什么意思
- 国际食品科技合作组织是什么意思
- 国际食物、营养与社会经济发展讨论会论文集是什么意思
- 国际饭店是什么意思
- 国际饮水供应和环境卫生十年是什么意思
- 国际首饰市场是什么意思
- 国际马传染性子宫炎证明立法是什么意思
- 国际马传染性贫血证明立法是什么意思
- 国际马流感证明立法是什么意思
- 国际马脑髓炎证明立法是什么意思
- 国际马铃薯研究组织是什么意思
- 国际马鼻疽证明立法是什么意思
- 国际骆马保护管理立法是什么意思
- 国际骆马研究教育立法是什么意思
- 国际高发牛海绵状脑病国家进口牛肉证明立法是什么意思
- 国际高发牛海绵状脑病国家进口牛胚胎证明立法是什么意思
- 国际高发牛海绵状脑病国家进口牛证明立法是什么意思
- 国际高发牛海绵状脑病国家进口肉骨粉证明立法是什么意思
- 国际高等教育百科全书出版是什么意思
- 国际高致病性禽流感证明立法是什么意思
- 国际鲸类保护管制立法是什么意思
- 国际鸟类保护会议是什么意思
- 国际鸡伤寒和鸡白痢证明立法是什么意思
- 国际鸭病毒性肝炎证明立法是什么意思
- 国际鸭病毒性肠炎证明立法是什么意思
- 国际鹤类保护与研究是什么意思
- 国际麻醉品管制局是什么意思
- 国际麻醉品管制局是什么意思
- 国际黄麻组织是什么意思
- 国际黑格尔协会是什么意思
- 国际黑格尔学会是什么意思
- 国际黑海捕鱼立法是什么意思
- 国难会议是什么意思
- 国雅品是什么意思
- 国风是什么意思
- 国风是什么意思
- 国风是什么意思
- 国风是什么意思
- 国风今译是什么意思
- 国风尊经是什么意思
- 国风是否民歌是什么意思
- 国风民歌说是什么意思
- 国风的普通话翻译是什么意思
- 国风省篇一卷是什么意思
- 国风诗旨纂解是什么意思
- 国风诗旨纂解是什么意思
- 国风选译是什么意思
- 国风选译是什么意思
- 国风非民歌说是什么意思
- 国香是什么意思
- 国魂是什么意思