骨胳肌组织
骨胳肌组织是由骨胳肌纤维构成的一种肌组织,肌纤维间有结缔组织、丰富的毛细血管和神经。因其肌纤维有横纹而属于横纹肌。骨胳肌组织构成解剖学所见的肌肉;还分布在舌、咽、食管和喉;使肢体进行运动,参与机械性消化、呼吸和发声。
骨胳肌纤维的微细结构 骨胳肌纤维是细长圆柱状有横纹的多核细胞,直径约10~100μm,长度不一,最短的约1mm,一般长约3~40mm,有的更长,两端钝圆。在短肌肉中,肌纤维可与肌肉长度相等,在长肌肉中,肌纤维达不到肌肉末端,中途以结缔组织相连。肌纤维表面有肌膜,即细胞膜(图1)。细胞核有多个,在长的肌纤维中多达数百个。胞核呈卵圆形,染色质较少,核仁明显。胞核一般位于肌纤维周边,靠近肌膜;但可因肌纤维的类型和动物种属的不同而异,如红肌纤维和蛙骨胳肌纤维的胞核可位于肌纤维内部。肌纤维的胞质常称肌浆,其中有大量肌原纤维、丰富的线粒体、发达的肌浆网及高尔基复合体、溶酶体、糖原颗粒和肌红蛋白等。
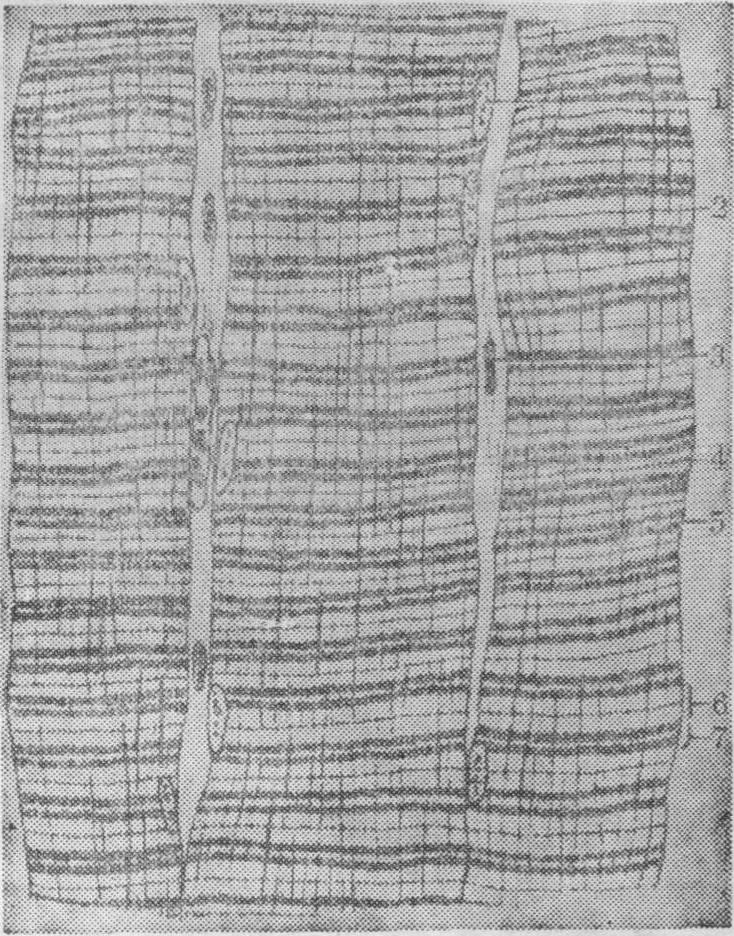
图1 骨胳肌纤维纵切面
1.细胞核 2.肌膜 3.成纤维细胞 4.间线 5.H带 6.明带 7.暗带
肌膜 是肌纤维表面的细胞膜,在电镜下,由单位膜构成,其外面附有一层基板和网状纤维(图2)。躯体运动神经纤维的终末附于每条肌纤维表面,形成运动终板。该处肌膜稍凹陷形成小槽,还向肌浆方向下陷形成许多小皱褶,在肌膜上有乙酰胆碱受体(N受体)(参见“神经末梢”条)。肌膜和基板一起从肌纤维表面凹入细胞内,形成垂直于肌纤维长轴的小管,称为横小管或T小管。横小管伸到肌原纤维之间,在同一水平面上的横小管分支相连环绕每一肌原纤维(图2)。哺乳动物的横小管位于明、暗带交界水平,而鱼类、两栖类和鸟类的位于间线水平。从细胞表面观察,在上述水平的肌膜上可见一排排横小管开口。横小管是肌膜向内凹陷所成,直径约20~40nm,管腔与细胞外间隙相通,内含细胞外液。横小管的广泛存在,使神经刺激产生的去极化波迅速由肌膜传到肌纤维内部,使整个肌纤维一致地收缩。
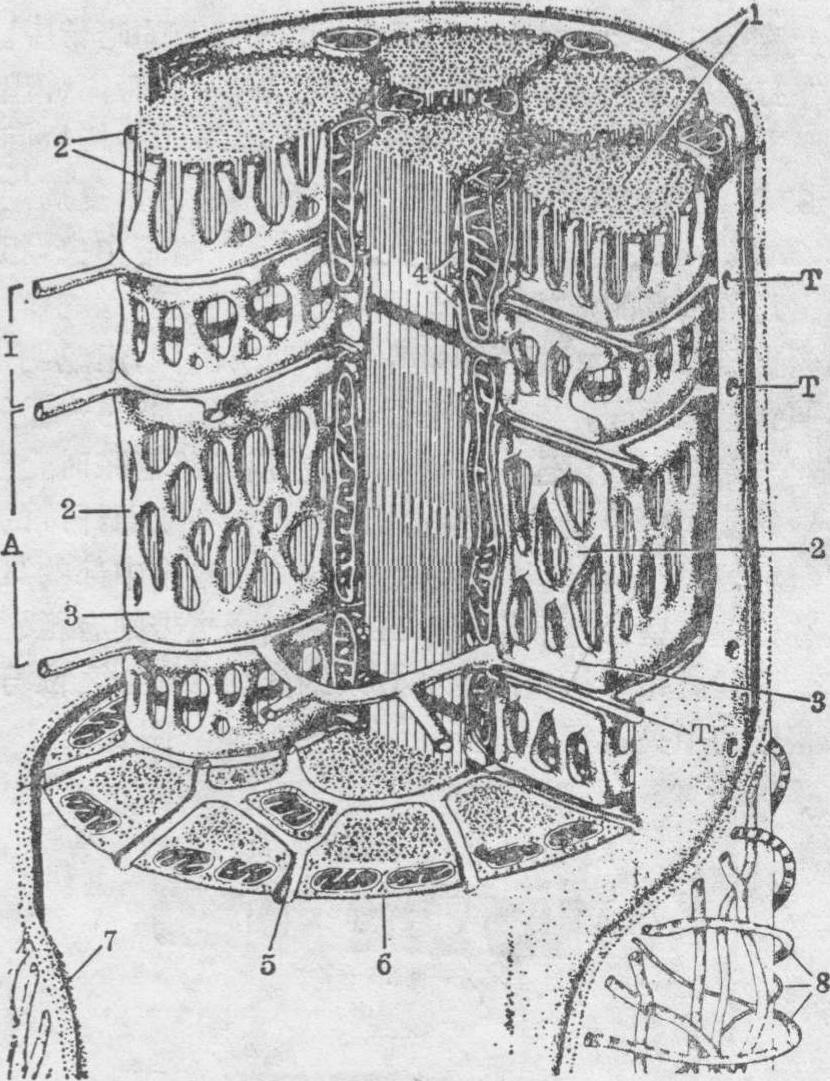
图2 骨胳肌纤维超微结构模式图
1.肌原纤维 2.肌浆网 3.终池 4.三联体 5.横小管 6.肌膜 7.基板 8.网状纤维 I.明带 A.暗带 T.横小管开口
肌原纤维 光镜下,肌原纤维呈细长丝状,直径1~2μm,沿肌纤维的长轴平行排列。肌原纤维上都显有明暗相间的带,而且各肌原纤维的明带、暗带均整齐地排列在同一水平上,故整个肌纤维显有明暗相间的横纹(图1)。用铁苏木素染色时,暗带着色深,明带着色浅。用偏振光显微镜观察时,暗带呈较明亮的双折光性或各向异性(ani-sotropy),故又称A带; 而明带呈较弱的双折光性或各向同性(isotropy),故又称I带。肌肉舒张时,明带宽约0.8μm,暗带宽约1.5μm。肌肉收缩时,明带变窄,暗带不变。暗带中有色浅的H带。H带中又有一条深色的中线,也称M线。明带中有一条染色深的间线,也称Z线、Z膜或Z盘。相邻两间线之间的一段肌原纤维,称为肌节。每个肌节包括1/2明带+暗带+1/2明带。肌节是肌原纤维的结构和功能的基本单位,其长度在肌肉舒张时约为2.3μm,收缩时则变短。在肌纤维的横切面上,肌原纤维呈点状,由于它们在肌浆中配布不均匀,故集成许多大小及形状不一的区域,称Cohnheim区(参见“肌肉”条图)。
在电镜下,肌原纤维由两种很细的肌丝即粗丝和细丝构成,两种肌丝均顺肌原纤维的长轴平行排列,在肌节内各居一定位置,彼此保持一定距离,又互相穿插(图3)。粗丝直径10~20nm,长约1.5μm,位于肌节的暗带中。粗丝在H带表面光滑,在H带两侧则以相等的距离伸出许多横突,称横桥。相邻横桥间距约为14.3nm,横桥间角为60度,每旋转一周伸出6个横桥,横桥的重复周期为42.9nm。每条粗丝约有216个横桥。在暗带中央,粗丝伸出几条垂直的丝状突,形成中线,有固定粗丝的作用。细丝直径5~7nm,长约1μm,位于肌节两侧,组成明带,并伸到粗丝之间,与粗丝平行,共同组成暗带的一部分。细丝一端与间线相连,另端游离,在肌纤维舒张时,止于H带外侧。由于上述肌丝配布的特点,在肌原纤维的横切面上,明带只有细丝,呈六角形排列;暗带中部的H带只有粗丝,呈三角形排列; H带两侧的暗带则有粗、细两种肌丝。每条粗丝周围有6条细丝;每条细丝位于3条粗丝的中央。粗、细丝相距10~20nm(图3)。
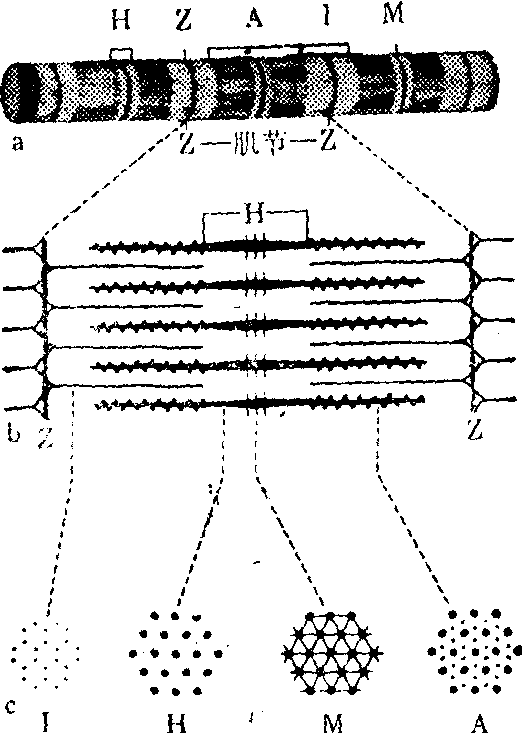
图3 骨胳肌纤维的肌原纤维
a.肌原纤维纵切面低倍电镜示意图,示肌节的各带
b.肌原纤维纵切面高倍电镜示意图,示粗丝和细丝在肌节内的分布
c.肌节横切面高倍电镜示意图,示肌丝在各带的位置关系
Z.间线 I.明带 A.暗带 H.H带 M.中线
间线位于明带的中央,也常称间膜,是肌节的分界和细丝附着处,细丝分别垂直地固定在间线两侧。间线由Z丝和间膜基质组成,含α辅肌动蛋白。根据两栖类肌肉的观察,在间线两侧都是每条细丝连接4根Z丝,而这4根Z丝斜穿间线至间线另一侧,又各与一条细丝相连(图4)。如此,Z丝在间线部位构成一个网。从间线侧面观,Z丝连成锯齿状(图3、图4)。哺乳类肌肉间线的构造更为复杂。
(1)粗丝: 主要由肌球蛋白集合而成,故又称肌球蛋白丝。肌球蛋白分子的分子量约为500,000,分子全长约150nm,可分为球状头和杆状尾两部分,形似豆芽。每一肌球蛋白分子由2条分子量各约200,000组成的长多肽链即重链,和4条分子量各约20,000组成的短多肽链即轻链共同组成。两条重链互相缠绕成α双螺旋,形成分子的杆部,长约140nm,直径2~3nm。在重链的一端与轻链盘折一起形成一对豆瓣状部分,称为球头,长约10nm,宽约20nm,分子量约120,000~180,000。用胰蛋白酶水解后,可将肌球蛋白分解为两个组分:即轻酶解肌球蛋白(light meromyosin,LMM) 和重酶解肌球蛋白 (heavymeromyosin,HMM)。LMM为杆部的大部分,长约90nm。HMM长约60nm,可被木瓜酶水解成两个亚单位,即球头(HMMS-1)和与其相连的一小段杆部(HMMS-2)(图5)。
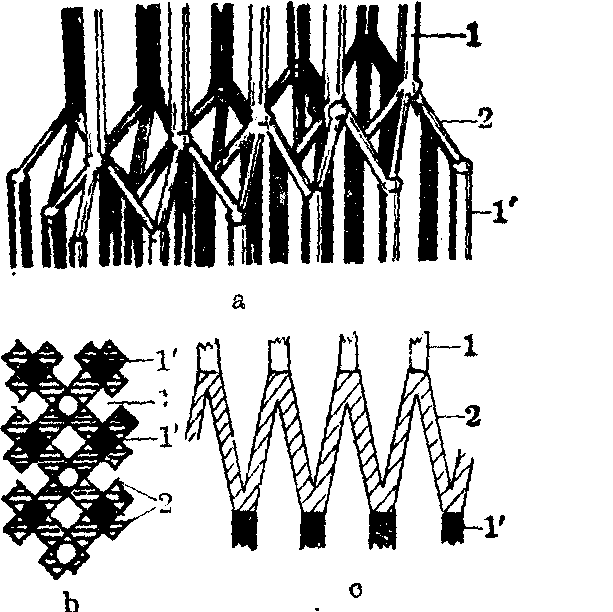
图4 间线结构示意图
a.间线的立体结构,示间线两侧的细丝与Z丝的关系 b.间线的表面观 c.间线的侧面观
1.间线一侧的细丝 1′.间线另一侧的细丝 2.Z丝
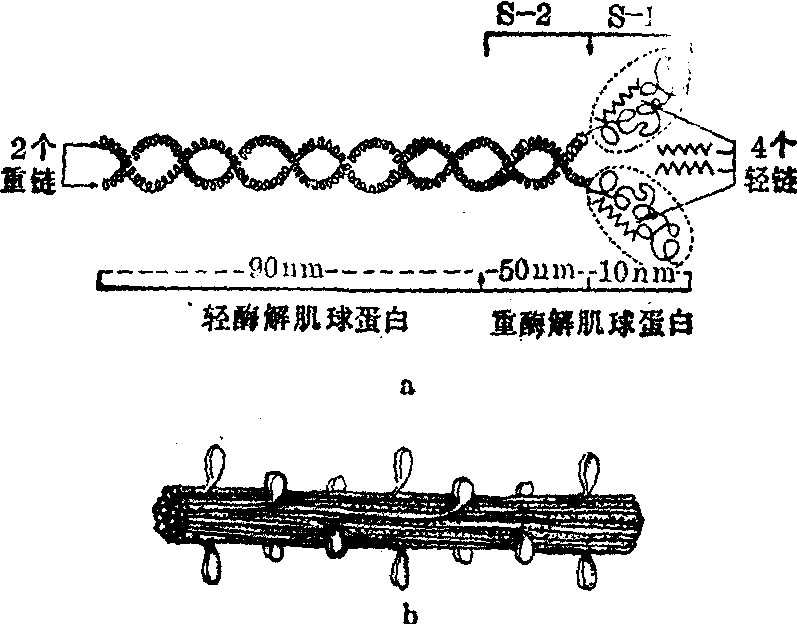
图5 粗丝分子结构示意图
a.肌球蛋白分子 b.由肌球蛋白分子组成的粗丝(示一段)
许多肌球蛋白分子互相规则集合一起构成粗丝 (图5)。各分子的LMM杆部组成粗丝骨架;在中线两侧的各分子球头端分别朝向粗丝两端,球头突出粗丝表面构成横桥头部,HMMS-2杆部则构成横桥臂。每个球头均含有ATP酶,还有一个与ATP结合的位点和一个与细丝的肌动蛋白结合的位点,球头和HMMS-2与LMM相接处还可弯曲,这些特点对肌肉收缩有重要作用。在粗丝上,每隔42.9nm的间隔有由C蛋白组成的开口环,箍在粗丝表面。
(2)细丝: 由肌动蛋白、原肌球蛋白和肌钙蛋白组成。肌动蛋白是细丝的主要成分,故细丝又称肌动蛋白丝。肌动蛋白分子的单体称G-肌动蛋白,呈椭圆形,直径约4.5~5.6nm,分子量约为45,000。球形单体不对称,每个单体都分前、后两极。两排球形单体前、后相接,聚合成两条念珠状链条,互相扭转成a-双螺旋,形成有极性的细丝状的F-肌动蛋白,构成细丝的主体(图6)。每个球形单体均有一个能与肌球蛋白结合的位点。
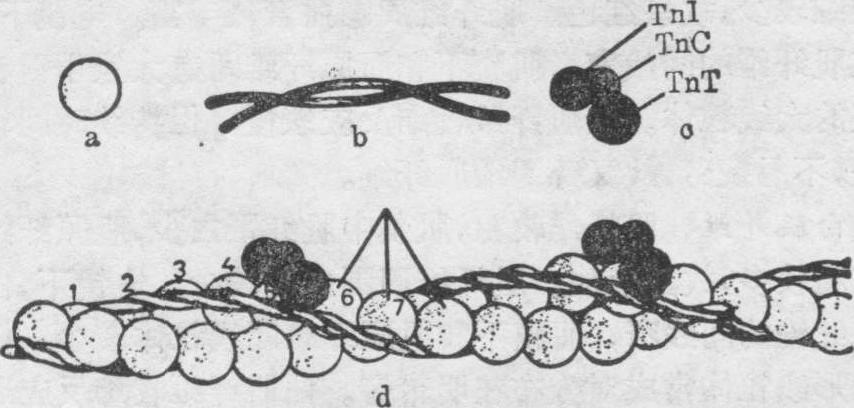
图6 细丝分子结构示意图
a.肌动蛋白分子球形单体 b.原肌球蛋白分子c.肌钙蛋白分子,分三个亚单位:Tn T肌钙蛋白T、TnC肌钙蛋白C、TnI肌钙蛋白I d.由三种蛋白分子组成的细丝(示一段)
原肌球蛋白分子呈线状,由两条多肽链拧成α双螺旋,长约40nm,分子量约70,000。原肌球蛋白分子也有极性,分子首尾相接,形成原肌球蛋白丝,位于肌动蛋白的双螺旋沟内(图6)。一个原肌球蛋白分子跨越7个肌动蛋白单体。原肌球蛋白与肌钙蛋白以1:1的比例形成复合体。
肌钙蛋白呈椭圆形,在细丝上,每隔40nm有一肌钙蛋白分子结合在一个原肌球蛋白分子上(图6)。肌钙蛋白分三个亚单位:
❶肌钙蛋白T(TnT):分子量约37,000~38,000,它与原肌球蛋白有很强的结合力,是肌钙蛋白与原肌球蛋白相连接的亚单位;
❷肌钙蛋白I(TnI):分子量为24,000,有抑制肌球蛋白与肌动蛋白相互作用的能力;
❸肌钙蛋白C(TnC):分子量最小,约为17,000~18,000,是肌钙蛋白的主要成分,与Ca的亲和力很强。每个TnC分子可与两个Ca离子结合,对肌肉收缩和舒张起重要的调节作用。
肌浆网 也称肌质网,是由特化的滑面内质网组成的膜管系统,在骨胳肌纤维内特别发达,具有调节肌浆内钙离子浓度的作用。肌浆网在两条相邻的横小管之间包绕每条肌原纤维。据肌浆网的分布和形状不同可分为互相连通的三部分:
❶小管网: 位于H带周围,呈小管状,互相吻合成密网,网孔较小;
❷纵小管: 位于暗带和明带周围,这些小管的走向与肌原纤维的纵轴大致一致;
❸终池:在哺乳动物骨胳肌纤维中位于明、暗带交界水平,在横小管两侧各有一终池,是分别由明带或暗带的纵小管吻合而成的横行膨大部分(图2)。横小管和它两侧的终池合称三联体。哺乳动物的骨胳肌纤维中,因三联体位于明、暗带交界水平,故每个肌节范围有两套三联体。而鱼类、两栖类和鸟类的三联体位于间线水平,每个肌节平均只有一组三联体。
肌浆网的单位膜上镶嵌的蛋白质中的70~80%是钙泵蛋白,它的分子量约100,000,是一种ATP酶,此酶必须有Ca2+和Mg2+存在才有活性。酶分解ATP时释放出能量,使钙泵蛋白分子构形发生变化,将肌浆内的Ca2+输送到肌浆网内,从而使肌浆网内Ca2+浓度比肌浆内高数千倍。肌浆网的膜上还有受磷酸蛋白和收钙素。受磷酸蛋白的分子量为22,000,与钙泵蛋白相邻接,能与磷酸结合,可加速钙泵自肌浆中摄取Ca2+的速度,故对钙泵运输速度起调节作用 。收钙素约占肌浆网膜蛋白的15%,其分子量为55,000。肌浆网内贮存的Ca2+约80%是与收钙素结合的。
肌浆的其它成分 包括
❶线粒体: 肌浆内有丰富的线粒体,位于肌膜内面,胞核附近及肌原纤维之间,其长轴与肌原纤维平行。在肌原纤维间的线粒体,常成对整齐地排列在间线两侧的明带旁。线粒体嵴既多又密。线粒体的主要功能是为肌肉收缩提供能量。线粒体的数量因肌纤维的类型而有差异;
❷高尔基复合体:多位于胞核两端的肌浆中,在成年的骨胳肌纤维中不甚发达,在肌纤维发生和再生时,较为明显;
❸溶酶体: 在正常肌纤维内很少,在病理应激情况下,如切断支配肌纤维的神经后,可见肌浆中有许多溶酶体;
❹糖原颗粒:肌浆中含大量糖原颗粒,多分布在肌原纤维间和肌浆网间;
❺脂滴: 分布在肌原纤维间;
❻肌红蛋白: 是肌纤维特有的一种蛋白质,弥漫分布在肌浆基质中,呈红色。不同类型的纤维因肌红蛋白含量不同而显不同颜色。肌红蛋白与氧的亲和力较强,当肌肉缺氧时可放出氧,以供肌肉收缩的急需。
骨胳肌纤维收缩机理 光镜早已证明,骨胳肌收缩时肌节变短是由于明带变窄,而暗带长度不变,但对肌纤维收缩的机制尚不了解。电镜观察发现,肌节是由粗、细肌丝相互穿插而成。在此基础上,Hanson和Huxley(1953,1957)提出 “肌丝滑动学说”,主张肌肉收缩时,粗、细丝的长度和暗带的宽度均无改变,肌肉收缩是由于细丝在粗丝之间向中线滑动的结果。细丝向中线滑动,导致粗、细丝互相穿插的部分增加和肌节两侧细丝的游离端互相靠拢,于是明带变窄、H带变窄以至消失,间线被牵拉靠近暗带,结果肌节和肌原纤维变短,从而肌肉缩短(图7)。
肌纤维收缩时细丝向中线滑动是受粗丝的横桥牵引所致,在此过程中发生一系列分子水平的变化(图8、图9).肌肉静止时,粗丝肌球蛋白头部 (即粗丝横桥的头) 的ATP酶处都结合着一分子ATP,简称为待发肌球蛋白,但此时肌浆内的Ca2+浓度很低,细丝肌动蛋白单体上与肌球蛋白头部结合的位点被原肌球蛋白遮盖,故肌球蛋白不能与肌动蛋白接触。当肌纤维受到神经刺激时,肌膜的去极化沿横小管传到肌纤维内部,通过三联体使肌浆网的膜去极化,引起肌浆网贮存的Ca2+释放与细丝的肌钙蛋白C结合,改变了肌钙蛋白的空间构型,驱使原肌球蛋白陷入肌动蛋白的螺旋沟内,而将肌动蛋白上的肌球蛋白结合位点暴露出来。肌球蛋白头部随即,与肌动蛋白接触,同时头部的ATP酶在Ca2+存在下被肌动蛋白激活,迅速分解ATP,放出能量。这些能量促使肌球蛋白头和HMM与LMM连接处向中线方向旋转,于是牵引细丝向中线滑动(图8、图9)。球头每次旋转可使细丝移动5~10nm的距离。粗丝上虽有多个球头,但每一瞬间只有少数与肌动蛋白接触。当先与肌动蛋白相接的球头牵移细丝后,另一些待发肌球蛋白又与肌动蛋白接触,而先与细丝相接的球头则结合上一新ATP,与肌动蛋白脱离并恢复原位,准备再次与肌动蛋白接触。如此重复,终使粗丝与细丝大部重叠,导致肌肉收缩。当神经刺激停止后,肌浆网的钙泵迅速将肌浆的Ca2+泵回肌浆网内,使肌浆的Ca2+浓度降低,于是粗、细丝脱离接触。
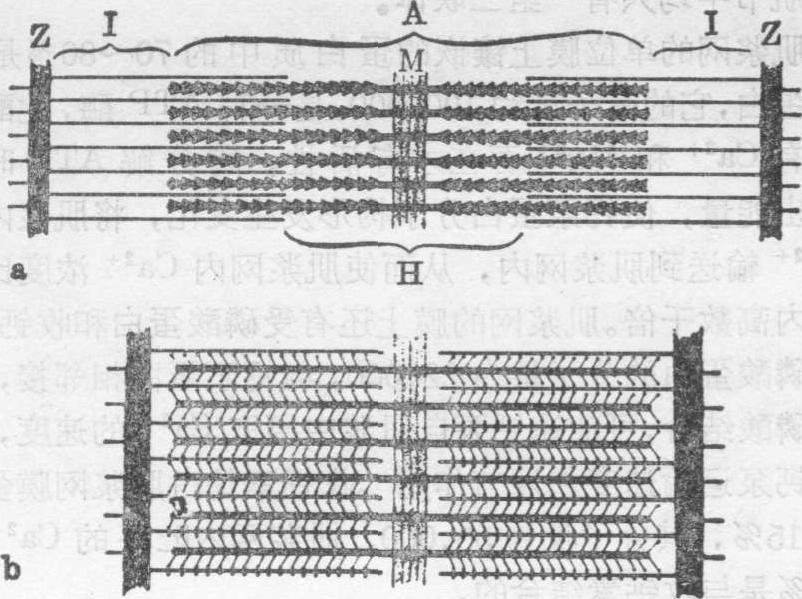
图7 骨胳肌收缩时粗、细肌丝位置变化示意图
a.舒张状态 b.收缩状态
Z.间线 I.明带 A.暗带 H.H带 M.中线
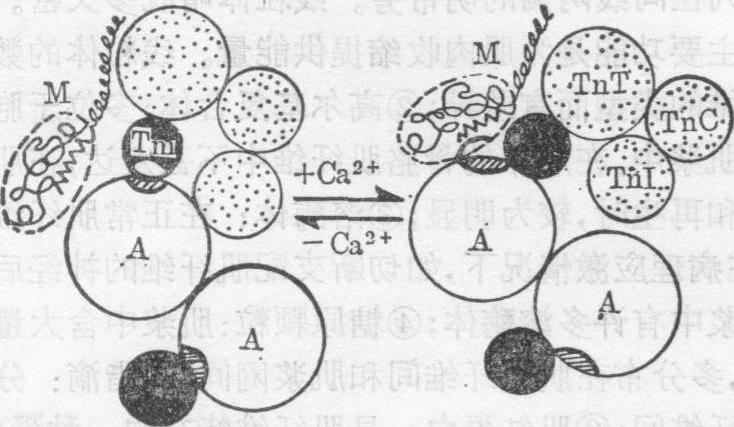
图8 骨胳肌纤维收缩(右)与舒张(左)时粗、细丝中各种蛋白分子的位置变化
A.肌动蛋白单体 M.肌球蛋白头 Tm.原肌球蛋白TnT.肌钙蛋白T TnC.肌钙蛋白C TnI.肌钙蛋白I
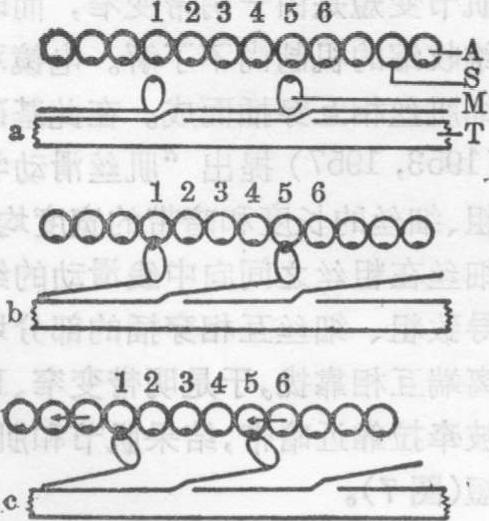
图9 肌丝滑动机理示意图
a.舒张时,示粗丝肌球蛋白头未与细丝肌动蛋白接触。A.细丝1~6示球形肌动蛋白单体; S.与肌球蛋白头结合的位点;M.肌球蛋白头T.粗丝
b.收缩开始时,粗丝肌球蛋白头与细丝肌动蛋白单体接触
c.粗丝肌球蛋白头向中线方向旋转,牵引细丝向中线滑动(箭头示滑动方向)
骨胳肌纤维的类型 依据肌浆中肌红蛋白和肌原纤维的多少及其它特点,可将肌纤维分为红肌纤维、白肌纤维和中间型肌纤维。以红肌纤维或白肌纤维为主组成的肌肉,颜色深浅不同,分别称为红肌和白肌。
红肌纤维 肌纤维较细,肌浆中肌红蛋白多,肌原纤维较细较少,故呈红色。肌纤维周围血管较丰富。电镜下,红肌纤维的间线宽,肌膜下和肌原纤维间有大量嵴很密集的大线粒体。红肌纤维收缩反应缓慢,但持续时间较长,不易疲劳,故又称慢缩肌纤维。
白肌纤维 肌纤维较粗,肌浆中肌红蛋白少,肌原纤维较粗较多,故颜色淡。肌纤维周围血管较少。电镜下,白肌纤维的间线窄,肌膜下和肌原纤维间线粒体较少,椭圆形线粒体常成对分布在明带旁。白肌纤维收缩反应迅速,但持续时间较短,容易疲劳,故又称快缩纤维。
中间型肌纤维 形态特征介于红、白肌纤维之间。电镜下,间线较窄,线粒体比红肌纤维的较小,嵴也较少。
显示线粒体酶和肌原纤维ATP酶的组织化学方法,可清楚地区分三型肌纤维。如显示线粒体琥珀酸脱氢酶时,红肌纤维的呈色反应最深,白肌纤维的反应最弱,中间型肌纤维的反应界于二者之间。许多哺乳动物如大白鼠、小白鼠、家兔等的红肌和白肌明显可辨,甚至可由一型肌纤维构成一块肌肉。人的肌肉中,三型肌纤维混杂存在,但它们在各肌肉中的数量比例不同。有的肌肉如小腿腓肠肌以白肌纤维为主,颜色略淡,适于敏捷的运动。经常处于运动的眼肌、咀嚼肌和呼吸肌等,则以红肌纤维为主。
骨胳肌组织的生长和再生 在骨胳肌纤维的基膜内面,见扁梭形细胞位于肌纤维表面的浅凹中,称肌卫星细胞。此细胞胞质少,核长圆形,光镜下看不清细胞界限。它们可能是贮备的干细胞,能增生分化为成肌细胞,参与肌纤维的生长、肥大和再生。骨胳肌纤维是高度分化的细胞,一般认为童年后骨胳肌纤维的数目不再增多,但随个体的发育生长,肌纤维的体积可渐增大,包括增粗和增长。肌纤维增粗是由于肌浆增多,游离核糖体形成新肌丝,使肌原纤维变粗,以至形成新的肌原纤维。增长是由肌纤维两端的游离核糖体产生新肌丝蛋白,形成新肌节,衔接在肌原纤维末端,从而使肌纤维变长。也有人认为,肌卫星细胞与肌纤维融合而使之增长。肌肉的发达程度与肌肉的运动有密切关系。长期停止运动,肌肉发生萎缩;长期高强度活动,肌肉则明显发达,这种现象称为适应性肥大。
低等脊椎动物的骨胳肌有明显的再生能力。人骨胳肌的再生能力很低。肌卫星细胞与骨胳肌再生有关。肌纤维受损伤后,肌卫星细胞分裂增殖,变为纺锤形的成肌细胞,许多成肌细胞彼此融合而成管状的多核细胞,称肌管。肌管内生成肌丝增多,胞核移向细胞周缘,最后分化成有横纹的肌纤维。有人认为,肌肉损伤后,在最初2~3天出现退行性变化,中性粒细胞和巨噬细胞侵入其中,清除坏死组织,损伤部位附近的肌纤维残端肌浆形成芽状突起,称为肌芽。肌芽向损伤区延伸,成纤维细胞生成结缔组织性肌内膜管,引导肌芽长入其中,与相对端伸来的肌芽相互衔接,实现肌纤维再生。另有人认为,在骨胳肌受损的部位,未受损伤的肌纤维变为成肌细胞,然后成肌细胞的胞核一再分裂,形成多核细胞,继之出现肌原纤维,遂产生新的肌纤维。严重创伤引起的大范围肌肉缺损,由成纤维细胞产生纤维性瘢痕组织填补。
- 伤寒汇编 明 王翔是什么意思
- 伤寒汇解 清 蒋钟尹是什么意思
- 伤寒汇集一卷 清 邹大麟是什么意思
- 伤寒沙门菌脂多糖是什么意思
- 伤寒治例是什么意思
- 伤寒治例是什么意思
- 伤寒法祖是什么意思
- 伤寒津氏微是什么意思
- 伤寒活人书括指掌图论 (类编)是什么意思
- 伤寒活人指掌补注辨疑是什么意思
- 伤寒活人指掌补注辨疑是什么意思
- 伤寒活人指掌补注辨疑是什么意思
- 伤寒派是什么意思
- 伤寒流注是什么意思
- 伤寒海底眼是什么意思
- 伤寒温疫抉要是什么意思
- 伤寒温疫条辨是什么意思
- 伤寒温疫条辨是什么意思
- 伤寒温疫条辨是什么意思
- 伤寒温疫条辨是什么意思
- 伤寒温疫条辨是什么意思
- 伤寒源流是什么意思
- 伤寒源流是什么意思
- 伤寒溯源集是什么意思
- 伤寒溯源集是什么意思
- 伤寒溯源集是什么意思
- 伤寒溯源集是什么意思
- 伤寒潮热是什么意思
- 伤寒潮热是什么意思
- 伤寒点精是什么意思
- 伤寒片玉集三卷 宋 卢昶是什么意思
- 伤寒琐言是什么意思
- 伤寒生意四卷 元 熊景先是什么意思
- 伤寒疫苗是什么意思
- 伤寒病症笺释 清 文锦华是什么意思
- 伤寒病问答是什么意思
- 伤寒瘟疫条辨是什么意思
- 伤寒瘟疫条辨六卷 清 杨浚是什么意思
- 伤寒百证歌是什么意思
- 伤寒百证歌是什么意思
- 伤寒百证歌是什么意思
- 伤寒百问是什么意思
- 伤寒百问 明 唐椿是什么意思
- 伤寒百问歌是什么意思
- 伤寒百问歌是什么意思
- 伤寒百问歌是什么意思
- 伤寒直指 清 强健是什么意思
- 伤寒直格是什么意思
- 伤寒直格是什么意思
- 伤寒直格是什么意思
- 伤寒直格是什么意思
- 伤寒直格是什么意思
- 伤寒直格 元 郭忠是什么意思
- 伤寒直格方 伤寒标本心法类萃是什么意思
- 伤寒直格方三卷 金 刘完素是什么意思
- 伤寒直解辨证歌是什么意思
- 伤寒真方歌括是什么意思
- 伤寒真方歌括是什么意思
- 伤寒科是什么意思
- 伤寒科是什么意思