腺垂体分泌的调节
腺垂体的分泌功能,一方面受中枢神经系统,特别是下丘脑的调节,另一方面也受外周靶腺所分泌的激素的反馈调节。
中枢神经系统对腺垂体分泌的调节
下丘脑-垂体系统 哺乳动物的下丘脑含有一些特殊的神经细胞,它们象内分泌腺细胞一样具有分泌激素的功能。这种能分泌激素的特殊神经细胞称为神经分泌细胞。例如下丘脑促垂体区(包括弓状核、室周围前核腹侧部分、视交叉后区内侧部分和正中隆起)的神经分泌细胞能分泌调节性多肽。下丘脑调节性多肽可以通过下丘脑垂体门脉系统的血液循环到达腺垂体,调节腺垂体的分泌功能。下丘脑与神经垂体的关系更为直接。下丘脑视上核和室旁核中的神经分泌细胞能分泌加压素和催产素,它们所制造的加压素和催产素可沿下丘脑垂体束中神经纤维的胞浆流动,到达位于神经垂体内的神经末梢,在身体需要时再释放入血液。由于下丘脑与垂体在解剖结构和生理功能方面都有密切的联系,因此生理学者将它们看作一个功能单位,称为下丘脑-垂体系统。
肽能神经元 末梢能分泌肽类激素的神经分泌细胞称为肽能神经元。因为下丘脑调节性多肽、加压素和催产素都属于肽类激素,所以下丘脑分泌这些激素的神经分泌细胞又称为肽能神经元。除加压素和催产素早已确定是由视上核和室旁核中的肽能神经元所分泌的以外,近年来应用免疫细胞化学的技术在其他肽能神经元的定位方面也取得了一定的进展。初步看来,分泌不同肽类激素的肽能神经元在下丘脑的分布区域也有所不同。现已确定,下丘脑分泌黄体生成素释放激素(LRH)的肽能神经元的胞体位于视前区、下丘脑前区及弓状核,其轴突则集中于正中隆起处。分泌GIF的肽能神经元在动物体内分布特别广泛,不仅存在于中枢神经系统的许多部位,甚至还存在于胃、肠、胰等神经系统以外的部位。但末梢在正中隆起处分泌GIF的肽能神经元的胞体则位于视前区及下丘脑前区。末梢在正中隆起处分泌促甲状腺素释放激素 (TRH) 的肽能神经元的胞体主要位于背内侧核,弓状核亦有之。至于分泌其他调节性多肽如促皮质素释放因子(CRF)的肽能神经元的定位,因为这些激素的化学结构尚未查明,所以还不能用免疫化学的技术精确地定出它们的存在部位。
下丘脑的调节性多肽 下丘脑一些区域的特殊神经细胞发出神经纤维到达正中隆起,其末梢可以释放一些具有生物活性的多肽,称为下丘脑调节性多肽(HRP)。这些HRP直接进入附近的毛细血管,经垂体门脉系统到达腺垂体,调节腺垂体激素的合成与释放。HRP作用于腺垂体中的靶细胞后,可立即引起分泌反应,将HRP除去时分泌反应立即停止。体内HRP的消除主要是通过肾的清除和血液中酶的降解作用。此外,HRP经过腺垂体进入体循环时立即被稀释,这也是一种及时消除HRP的有效途径。现在知道,HRP至少有9种:
❶TRH:1968年从羊的下丘脑提取物中分离提纯了TRH,并确定为三肽。TRH是第一个被人工合成的HRP (1969)。TRH能与分泌促甲状腺素的腺垂体细胞的质膜受体结合,激活腺苷酸环化酶,使细胞内cAMP合成增加,从而促进促甲状腺素的分泌。此外,TRH对生乳素的分泌也有强烈的刺激作用。
❷黄体生成素释放激素(LRH):1971年从猪的下丘脑提取物中分离提纯了LRH并确定为十肽。LRH已能人工合成。由于腺垂体分泌的两种促性腺激素有时有不平行的变化,因而曾设想下丘脑可能是通过两种释放激素分别调节两种促性腺激素的分泌,但是这种设想没有得到证实。目前认为两种促性腺激素均受LRH的调节,因此LRH又称为促性腺素释放激素(GnRH),但促进黄体生成素(LH)分泌的作用较强。LRH与腺垂体内靶细胞的受体部位结合后,激活腺苷酸环化酶,使细胞内cAMP增加,再促进黄体生成激素和卵泡刺激素的分泌。
❸生长激素释放抑制激素(GIH)和生长激素释放因子(GRF):1972年从羊的下丘脑提取物中分离提纯了GRF,现已能人工合成。GIH是由14个氨基酸组成的多肽,不仅能抑制生长激素的分泌,还能抑制促甲状腺素的合成和释放。除GIH外,下丘脑的提取物中还有GRF的存在,可以促进生长素的分泌。有人从猪的下丘脑提出一种十肽,认为可能就是GRF。切断下丘脑与垂体的联系后,生长激素的分泌减少,说明在正常情况下生长激素的分泌主要由GRF调节。
❹促皮质素释放因子(CRF):下丘脑存在一种能促进ACTH分泌的物质称为CRF。CRF是第一个被证实存在的HRP,但由于技术上的困难,其化学本质至今仍不清楚。加压素在体内及体外实验中都能刺激ACTH的分泌,但它是否就是生理性CRF还不能确定。
❺催乳素释放抑制因子(PIF)和催乳素释放因子(PRF): 下丘脑的提取物中含有一种能抑制催乳素分泌的物质,称为催乳素释放抑制因子 (PIF),可能是一种小分子肽。损伤下丘脑或切断垂体柄均可使催乳素的分泌增加,说明催乳素的分泌经常受到PIF的控制。儿茶酚胺和胆碱能物质都能抑制催乳素的分泌,其中多巴胺的作用最强。多巴胺也可直接作用于垂体,降低催乳素的分泌,因此有人认为多巴胺可能就是PIF,尚未最后决定。在某些条件下可以看到下丘脑还有促进催乳素分泌的作用。下丘脑的这种能促进催乳素分泌的物质就称为PRF,可能也是小分子肽,在人类,TRH是催乳素分泌的强烈刺激物,但在哺乳或其他伴有催乳素释放的情况下,TRH并不升高,而且在大白鼠,TRH却没有促进催乳素分泌的作用。因此,TRH不象是PRF。
❻促黑色细胞激素释放抑制因子 (MIF)和促黑色细胞激素释放因子(MRF):腺垂体的促黑色细胞激素(MSH)的分泌受下丘脑的调节。实验证明,下丘脑可分泌促黑色细胞激素释放因子,加强MSH的释放,还可分泌促黑色细胞激素释放抑制因子,抑制MSH的释放。目前已发现MRF为5肽,而MIF有3肽和5肽两种。3肽的活性大大强于5肽。
肽能神经元活动的调节 腺垂体所分泌的激素和腺垂体的靶腺激素可以分别通过短反馈途径或长反馈途径调节相应的肽能神经元的活动。下丘脑调节性多肽本身还能通过超短反馈途径调节肽能神经元本身。内外环境的变化也能通过高级中枢影响肽能神经元的活动。参与肽能神经元活动调节的神经介质的化学本质目前尚不十分清楚,但一般认为可能包括多巴胺、去甲肾上腺素和五羟色胺等单胺类递质。
(1) 多巴胺(DA):下丘脑多巴胺能神经元发出短纤维到达正中隆起部的外层,其神经末梢分泌DA,可直接作用于垂体,显著抑制催乳素的分泌。在下丘脑提取物中含有DA,若用氧化铝吸附后,这种下丘脑提取物对催乳素分泌的抑制作用便消失。所以,有些学者认为DA就是PIF。但是,另一些学者认为DA不是PIF,而具有促进PIF释放的作用,从而能抑制催乳素的分泌。多巴胺能神经元还可以调节LRH释放,促进FSH和LH的分泌。此外,DA可影响GIH或GRF分泌而调节生长激素的释放。
(2) 去甲肾上腺素(NE):下丘脑去甲肾上腺素能神经元释放的NE通过α-受体起作用,抑制CRF神经元分泌CRF,结果使ACTH的分泌减少。NE还可通过α-受体作用于分泌TRH、LRH、GIH和GRF的神经元,促进促甲状腺素,促性腺素和生长激素的分泌。NE对雌激素诱发的生乳素分泌有刺激作用。
(3) 五羟色胺(5-HT):五羟色胺能神经元能刺激催乳素的分泌,特别在吸吮或雌激素诱发的快速分泌中,这种刺激作用更明显。五羟色胺能神经元的这一作用是由于刺激PRF的分泌还是抑制PIF的分泌仍不清楚。给动物第三脑室注射5-HT,血浆LH和FSH水平降低,而直接注入垂体前叶则无此效应,说明5-HT对LH和FSH分泌的抑制效应不是对垂体的直接作用,而是通过对下丘脑分泌LRH的抑制而实现的。
(4) 组胺: 给犬脑室注射组胺可通过H1-受体的作用而刺激ACTH的分泌。用药物使H2受体兴奋时,则抑制ACTH的分泌。组胺刺激ACTH分泌的作用可能是通过加压素的释放。此外,刺激下丘脑H1受体也可增加催乳素的分泌,而刺激H2受体则降低催乳素的分泌。
肽能神经元释放激素的原理 一般认为下丘脑肽类激素的释放是由肽能神经元细胞体的动作电位引起的。当动作电位到达神经末梢时就使末梢细胞膜去极化,引起肽类激素的释放。可以利用大白鼠的排乳反射来说明这一点。在幼鼠吸吮乳头引起排乳反应出现之前,视上核和室旁核中有些肽能神经元能规律性地发生放电活动增强的现象,说明神经分泌细胞的电活动参与激素分泌的过程。此外,还可看到当改变细胞外液中K+的浓度使肽能神经元去极化时可以引起加压素、催产素、CRF和PIF的分泌。可能是神经末梢膜的去极化引起膜的Ca2+通道开放,Ca2+进入细胞内,再导致激素的释放。
腺垂体分泌的反馈调节 腺垂体的分泌可通过“长反馈”、“短反馈”和“超短反馈”三种途径进行反馈调节。
长反馈途径是指靶腺激素作用于腺垂体或下丘脑,从而调节腺垂体分泌的途径。腺垂体的各种促激素都有促进外周靶腺激素分泌的作用,当靶腺激素在血中的浓度过高时,又能反过来抑制相应促激素的分泌,称为负反馈调节。在一定的条件下,适量的靶腺激素也可促进相应促激素的分泌,称为正反馈。反馈调节是使血液中激素维持相对稳定的一种重要调节方式(见图)。
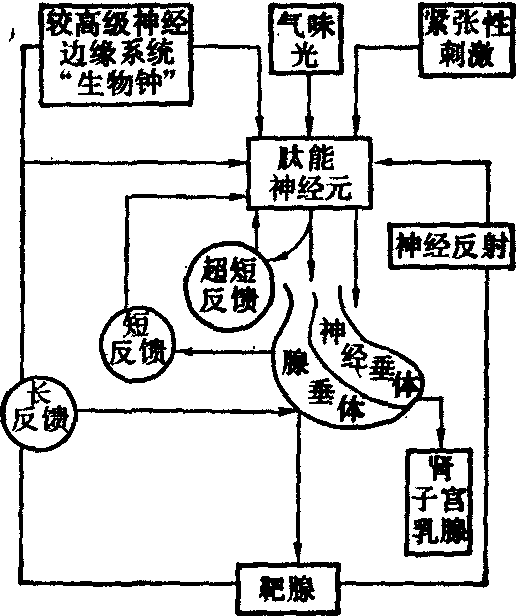
下丘脑肽能神经元系统的调节
靶腺激素可以直接作用于腺垂体,调节腺垂体促激素的分泌,也可作用于下丘脑,通过对HRP分泌的影响而调节腺垂体促激素的分泌。
❶甲状腺素主要作用于腺垂体,抑制促甲状腺素的分泌。甲状腺素还有与TRH竞争腺垂体细胞受体部位的作用,预先用甲状腺素处理的腺垂体,对TRH的反应能力显著降低,甚至丧失。至于甲状腺素对下丘脑的反馈控制,目前还没有一致的看法。有人将T4埋藏于下丘脑视前区,观察到对促甲状腺素的分泌有抑制作用。另一些人发现甲状腺素过多可刺激TRH合成酶的合成,甲状腺素过少则表现为抑制作用。于是认为甲状腺素对下丘脑TRH的分泌有正反馈作用。
❷肾上腺糖皮质激素主要作用于下丘脑,抑制CRF及MRF的释放,从而抑制ACTH及MSH的分泌。此外,糖皮质激素也可直接作用于腺垂体,阻断CRF对腺垂体的作用,从而使ACTH及MSH的分泌减少。
❸性腺激素特别是雌激素和孕激素,对下丘脑和腺垂体都有反馈作用。大剂量的雌激素可抑制LRH的分泌,降低垂体对LRH的敏感性,使FSH和LH的分泌减少。但FSH的分泌比LH的分泌更容易受雌二醇的抑制。在排卵前,成熟的卵泡所分泌的雌激素却能促进LRH的释放,提高垂体对LRH的敏感性,从而导致月经中期的LH峰。孕酮单独作用也能使LRH所引起的LH和FSH都受到抑制,而孕酮与雌二醇共同作用时,对LH的分泌的抑制作用就更为明显。
短反馈途径就是指腺垂体激素通过对下丘脑HRP分泌的反馈控制而调节自身的分泌,这种反馈途径较短,因而称为“短反馈”。例如,血中的ACTH达到一定浓度时能反馈作用于下丘脑,抑制CRF的分泌。血中促甲状腺素达到一定浓度时则反馈抑制TRH的分泌。此外,还有超短反馈途径,即指下丘脑分泌的HRP达到一定浓度时也能反馈抑制下丘脑分泌细胞本身。这一途径在HRP分泌的调节中可能具有一定的意义。
- 影响存款变化因素是什么意思
- 影响射精的药物是什么意思
- 影响居间成交的因素是什么意思
- 影响工业厂房开发的因素是什么意思
- 影响工业布局的因素是什么意思
- 影响 带动是什么意思
- 影响建筑业产品成本的因素是什么意思
- 影响建筑市场因素是什么意思
- 影响心理应激强度的因素是什么意思
- 影响心理行为性别差异的生物学因素是什么意思
- 影响心理行为性别差异的社会文化因素是什么意思
- 影响感情是什么意思
- 影响或意义很大是什么意思
- 影响房地产价格的因素是什么意思
- 影响技巧是什么意思
- 影响文学艺术发展是什么意思
- 影响斟酌的选择说是什么意思
- 影响普通住宅开发的风险因素是什么意思
- 影响期权费的因素是什么意思
- 影响母乳质量的因素是什么意思
- 影响气体交换的因素是什么意思
- 影响活动量大小的因素是什么意思
- 影响消费品流通的因素是什么意思
- 影响消费水平的因素是什么意思
- 影响消费者知觉的主要因素是什么意思
- 影响消费者行为的主要因素是什么意思
- 影响深远的功绩是什么意思
- 影响深远的徽剧是什么意思
- 影响甚大是什么意思
- 影响生长发育的因素是什么意思
- 影响生长发育的环境因素是什么意思
- 影响生长发育的遗传因素是什么意思
- 影响的忧虑是什么意思
- 影响矩阵是什么意思
- 影响社会宣传效果因素是什么意思
- 影响组织液生成的因素是什么意思
- 影响群众是什么意思
- 影响老人智力减退的因素是什么意思
- 影响老年人婚姻的因素是什么意思
- 影响老年人记忆的因素是什么意思
- 影响老年性功能的疾病是什么意思
- 影响肾小管与集合管功能的因素是什么意思
- 影响肾素血管紧张素药是什么意思
- 影响胆固醇合成的因素是什么意思
- 影响胆固醇吸收的因素是什么意思
- 影响能力发展变化的因素是什么意思
- 影响范围是什么意思
- 影响药物作用的心理因素是什么意思
- 影响营销组织决策的因素是什么意思
- 影响营销组织模式的因素是什么意思
- 影响衰老的先天因素是什么意思
- 影响衰老的后天因素是什么意思
- 影响视力的膜是什么意思
- 影响课堂教学的变量是什么意思
- 影响质量的九大因素是什么意思
- 影响量是什么意思
- 影响针灸效应的因素是什么意思
- 影响防御机制的因素是什么意思
- 影响静脉回心血量的因素是什么意思
- 影响静脉血回流的因素是什么意思