细胞间的相互作用
动物发育中细胞间的相互作用是各式各样的,可以是诱导作用,也可以是抑制作用。就作用方式来说,有的作用需要细胞的直接接触,另一些所需要的可能是间隔一定距离的化学物质的扩散。
诱导作用 在两栖类躯体的建成中,胚孔的背唇起关键性的作用。如果把蝾螈原肠胚期的背唇移植到另一同时期胚胎的腹部,不仅移植的背唇本身继续分化为脊索和肌节,还可以影响周围寄主的发育方向:原来将形成上皮的细胞,分化为神经组织,发育成神经系统,并且和寄主细胞一起共同组成次级的中轴器官,最后形成一个具有两套中轴器官的怪胎(图1)。移植的背唇创造了一个环境,改变了周围细胞发育的方向,使它们在腹部形成次级的神经系统。像这样的一部分胚胎细胞影响另一部分并决定其分化方向的作用称为诱导作用。
胚孔背唇能够诱导神经管的形成,神经管又能诱导其他胚胎结构。例如,在发育的过程中,中脑向左右各长出一个突起,即眼泡。眼泡又能诱导覆盖在它上面的表皮形成晶体,眼泡本身形成视网膜等结构。诱导形成的结构又能继续诱导其他结构的形成,像这样的一系列的诱导作用,在动物的胚胎发育中起了重要的作用。
从实验的结果来看,诱导作用不是由于细胞间的简单接触,而是由于诱导组织产生的某种物质。例如,被杀死的背唇细胞还保留它的诱导性能,这就指示从杀死的背唇可能有一种化学物质扩散到周围的细胞,促使后者向特定的方向分化。有一些实验观察支持这种看法,譬如,在背唇和外胚层之间放置一层薄膜,诱导作用照样能够发生。另外一个事实更直接地证明了这一点。将胚胎中胚层小块培养在生理盐溶液的“悬液”中,一星期之后,取出中胚层,放进一小块外胚层,外胚层将分化为神经细胞和色素细胞,但是,外胚层在没有放过中胚层的“悬液”中,不能进行这样的分化。可见中胚层细胞在 “悬液”中释放了某种物质,诱导外胚层分化为神经细胞。多年来,虽然进行过许多实验,企图获得这种物质,但是,到目前为止,它们的化学性质还有待进一步探索。
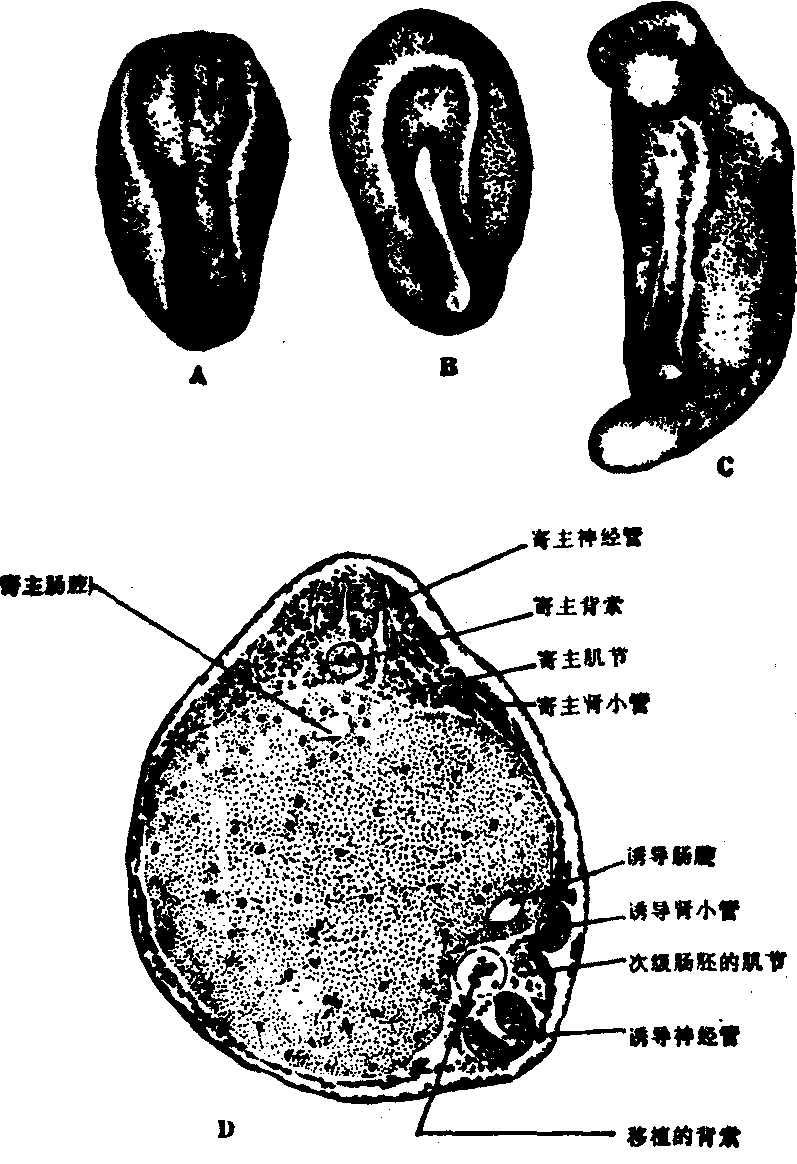
图1 移植背唇诱导次级蝾螈胚胎的形成
A和B.神经胚期的受主胚胎;A.背面,示寄主神经板;B.腹面,示诱导神经板C和D.尾芽期的同一胚胎; C.外观,示诱导的次级胚胎于左下方; D.横切面
诱导作用不但存在在中轴器官的形成中,以后器官的发生也离不开诱导现象。脊椎动物的许多器官是在间质细胞对上皮的诱导作用下形成的。
对于脊椎动物各种上皮结构(包括牙齿、毛发、羽毛、鳞片和皮腺)的形成和分化,间质细胞在上皮附近的聚集常常是必不可少的。但是两者之间的依赖关系在不同情况并不一样。上皮的命运高度依赖于衬垫其下的、专一的间质细胞。
鸡的皮肤有两种附加结构: 鳞片和羽毛。腿下部的皮肤上有鳞片,而羽毛则覆盖身体的其他部位。已经知道,这些上皮的附加结构是在中胚层的诱导下产生的。用鸡胚皮肤进行的实验进一步指出,皮肤附加结构的不同是因中胚层的区域性差异而决定的。如果将大腿中胚层移植到胚胎翅膀外胚层下面(图2,A),翅膀羽毛将按大腿羽毛的形态和排列而分化。如果将羽毛区域的中胚层和无羽毛区域的外层混合(图2,B)。会分化出羽毛。如果羽毛区的外层和鳞片区的中胚层混合(图2,C),外胚层则会形成鳞片。最后一个实验特别值得重视,因为它指出,在正常形成羽毛的外胚层中保存了形成鳞片的能力。异纲移植的实验还指出,上皮细胞的反应受本身遗传性质的限制,移植小鼠腹侧真皮到鸡的去除眼睛的角膜上皮之下,结果长出羽毛。小鼠中胚层的作用本来是诱导上皮形成毛发,但是,鸡的上皮对这一诱导刺激按它本身的遗传特性形成羽毛,而不形成毛发。
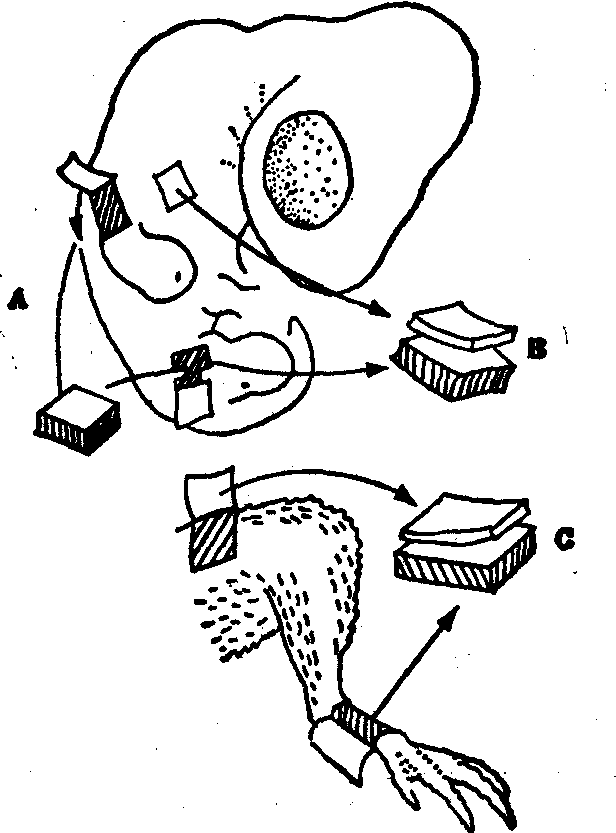
图2 证明中胚层控制上皮附加结构的实验
体内各种腺体(包括甲状腺、胸腺、肺、唾腺和胰腺)的形成和腺体细胞的生化特性的出现,间质细胞的作用和对皮肤的不同。这些腺体对间质细胞的依赖程度也不一样。在离体条件下,胰腺原基在发育的一定时期只要有间质细胞存在就可以继续发育。可是唾液腺上皮对间质细胞的要求就严格些,来自其他部位的间质细胞不一定能完全取代唾腺自身的; 另一方面,在鳃下腺芽体和不同来源的间质细胞混合培养的实验中,大多数间质细胞不能使上皮分化,而小鼠肺的间质细胞却能使鳃下腺上皮分支并形成腺体。但是,所用的肺间质细胞的量,必须大于鳃下腺间质细胞的。可见唾腺上皮对间质细胞虽然要求严格些,但在一定程度上也可为来自其他部位的间质细胞所取代。乳腺上皮与自身的间质细胞一起培养,形成窄细的管状结构,但与唾液的间质细胞一起时,则形成较多分支,而末端较膨大的结构外形上很像唾腺。
细胞分化不只是依赖于某些细胞对另一些细胞起作用,而且依赖于细胞之间的相互作用,不只是依赖于专一的物质从一部分细胞进入另一部分,而且依赖于两者之间物质的相互交换。在鸟类和哺乳类,泌尿系统的发育包括三个阶段,它们出现的顺序是从前向后。最早出现的是前肾,包括扭曲缠绕的前肾小管和直的、通入泄殖腔的前肾导管。在前肾导管的刺激下,中肾形成,前肾导管即成为中肾导管。后肾发育的开始是在中肾导管的后段长出一个芽体,芽体伸长,后段成尿道,顶端周围有间质细胞聚集,逐渐分支,以后形成肾脏的收集小管(图3,A)。聚集在收集小管尖端的间质细胞先形成上皮,然后折叠扭曲成管状,是肾小管的前身(图3,B)。用实验的方法已经证实,如果没有周围的间质细胞,尿道芽的分支就不会形成;相反,如果尿道芽的生长受到阻碍,达不到间质细胞,后者也不能折叠扭曲形成肾小管。这里尿道和间质细胞都是来自中胚层,在它们的相互作用下,分化才能正常进行。
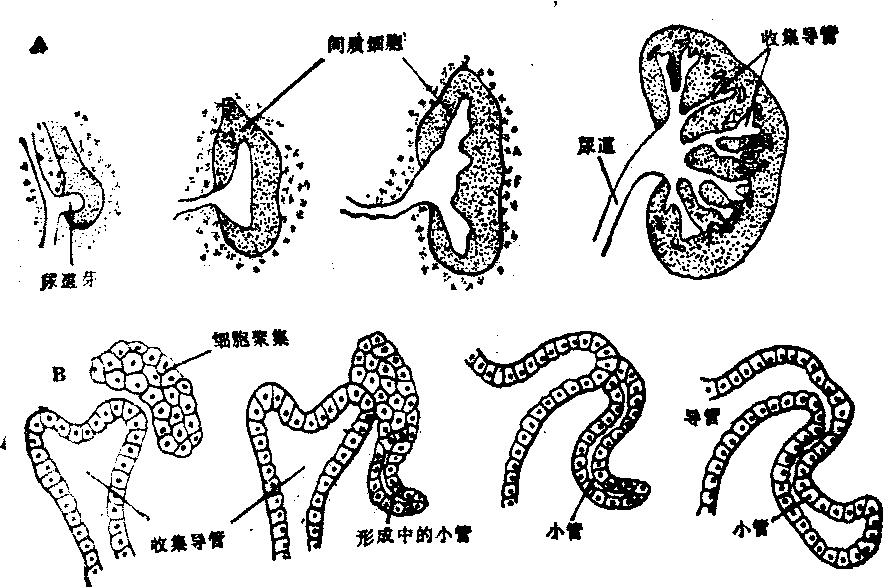
图3 哺乳类肾脏的发育
抑制作用 细胞间的相互影响还包括抑制作用,蝾螈晶体的再生是一个例子。在蝾螈幼虫或成体摘除晶体,可以从背部的虹膜再生出一个新的晶体(图4)。虹膜本来主要是由色素细胞组成,摘除晶体之后,背方边缘的色素细胞的色素颗粒减少,变得透明,同时细胞分裂增多,形成细胞团,以后分化为晶体。晶体再生的研究指出,再生晶体的能力局限在虹膜背部的边缘。不是虹膜的任何部分都具有这种能力,摘掉晶体后把虹膜的其他区域移到背部,不出现再生;而且,虹膜背部的再生能力在其他位置也可以表现出来,因为把这部分组织移到另一只摘除晶体的眼睛,不是位于背部,而是使它位于腹部,仍旧可以由它再生出晶体。
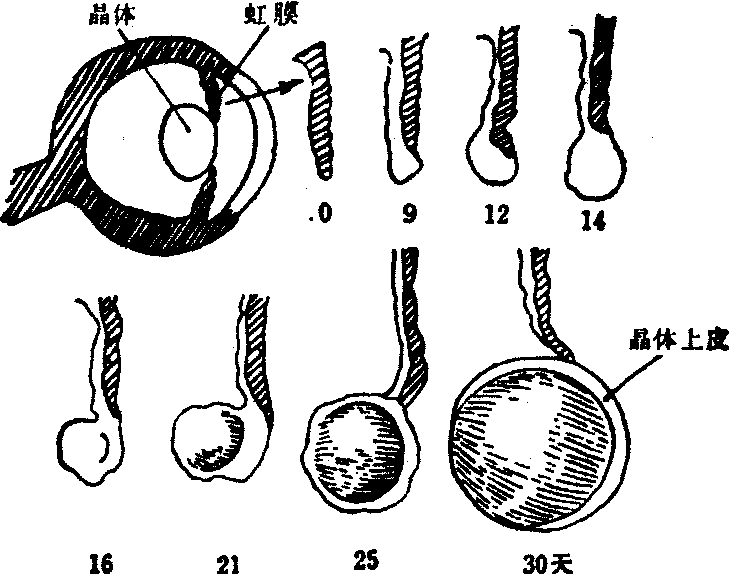
图4 蝾螈晶体从背部虹膜的再生
这部分细胞具有生长晶体的能力,但在正常的眼睛不表现出来,这要归之于晶体的存在。如果把虹膜的背部移到另一只未摘除晶体的眼睛里,不管使它位于哪一部位,都长不出晶体。更说明问题的是,如果在摘除晶体的眼睛里,经常地注射完整的(带有晶体的)眼腔中的液体,在注射期间,虹膜背部的细胞长不出晶体。由此可见,虹膜背部的细胞本来具有产生晶体的能力,正常的晶体会产生一种物质,对此起抑制作用。这种抑制作用,随着晶体的摘除而消失,因此虹膜背部形成晶体的能力得以显示出来。
细胞间的相互关系在高等动物虽然积累了不少资料,但是,关于在相互影响中起作用的因素知道得非常少,相比之下,在某些低等生物研究得比较深入。
在粘菌(Dictyostelium discoidaum)已经证明,细胞间的作用是通过某种化学物质而实现的。这是一种简单的真核生物,生活在森林中腐烂的落叶下,以土壤细菌为食物,生活周期可以划为三期。在食物丰足的情况下,是单细胞生活的变形体期,独立生存,以有丝分裂的方式增殖。当食物短缺时,变形体相互吸引,以群流会合之势,成千上万的变形体聚集到一起,形成一个形似蛞蝓的假原质团,它由千万细胞组成,能够移行,通常由黑暗移向光明。当环境干燥并有足够的光亮时,则形成子实体(图5)。由变形体期的细胞聚集成假原质团,这种成千上万细胞的协调行为,提示周围环境中有某种化学物质在起作用。现在已经知道,使变形体聚合的化学物质是环磷酸腺苷(cAMP)和腺苷酸磷酸二酯酶(使cAMP转变为腺苷酸的酶)。大约在变形体饥饿后6—7h,聚集的现象就开始出现,先是某一些细胞自主地周期性地释放cAMP,其他细胞对cAMP的反应是向这个信号移动,并且释放它们自己的cAMP。在释放cAMP以后,变形体暂时地对它不敏感,就在这期间周围的cAMP被腺苷酸磷酸二酯酶破坏,使细胞对新到达的cAMP又能发生反应。如此,这个现象重复发生,cAMP作为细胞外信使,将信息传递给周围的细胞,使整个过程协调进行,因而细胞聚集而形成多细胞个体。
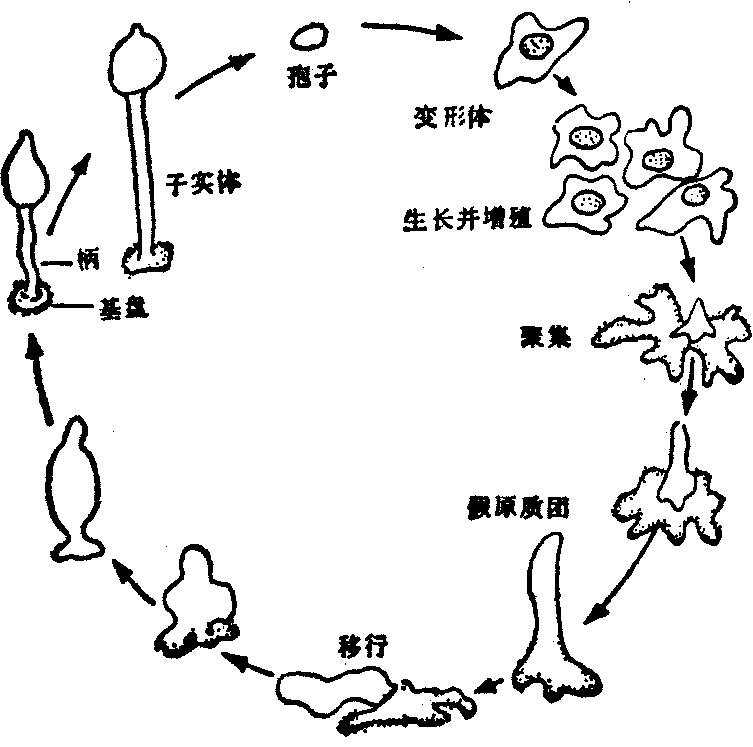
图5 粘菌的生活周期
在胚胎发育中,细胞迁移往往是分化的先驱。细胞先迁移到应该到达的位置,或者在环境的影响下进行分化,或者对环境施加某种影响。细胞迁移是一个复杂的现象,有时相同来源的细胞会发生分离,有时不同来源的细胞会相互聚合。这里既包含细胞的识别,也包含细胞间的粘着等等。
细胞聚合的观察最早是在海绵进行的。通过细小的网眼,把海绵挤碎,把分散的细胞相混,静止后会重新聚合。用这种机械的方法使两种颜色不同的种类——紫色的Heliclona occulata和红色的Microciona prolifera——分散后相混,结果特别明显。它们不会形成混合的聚集,而是分别形成两种不同颜色的团块。海绵细胞也可以用化学的方法分散,用缺钙镁的海水分散后,放回含钙镁的海水又能聚合。但是,用化学方法和用机械方法有所不同,用缺钙镁的海水处理后,分散的细胞不仅聚合得慢,而且在低温(5℃)下,根本不再聚合。而用机械的方法分散后,在低温聚合可以缓慢进行。这些结果提示,化学处理去除了某种聚合需要的物质,而必须在一定的温度下才能补足这种物质。同时,还发现用化学方法分散海绵细胞之后,海水中含有一种因子,在5℃时可以刺激分散的细胞聚合。这种因子是物种专一的,处理Heli-clona之后的海水能促使Heliclona细胞聚合,处理Microciona之后的海水只能促使Microciona细胞聚合。有工作指出,负责海绵物种专一的聚合因子可能是糖蛋白性质的,在Microciona,这种因子的分子量是几百万。
在脊椎动物,也可以利用细胞的聚合,对细胞之间的相互关系进行了解。用化学的方法处理两栖类胚胎组织,使成单个细胞的混悬液,然后把不同组织的细胞混合起来,使它们在正常生理溶液中重新聚合。起先,混合的细胞任意聚合。但是,随后它们相互分离,同类的聚在一起,形成组织专一的聚合,而且是按照正常胚胎中的情况排列。当上皮细胞和中胚层细胞混合时,中胚层细胞移入内部,形成间质细胞,而上皮细胞聚集在外部,逐渐在表面铺展,形成表皮(图6,A)。当上皮细胞、中胚层细胞和内胚层细胞一起混合的时候,内胚层细胞形成实心的球体,上皮细胞形成表面的表皮,而中胚层产生间质细胞,衬夹在表皮和内胚层之间(图6,B)。甚至同一胚层不同发育命运的细胞也要彼此分离,神经板细胞和上皮细胞的团聚物也发生分离,上皮在外形成表皮,神经板在内部形成类似神经管的结构 (图6,C)。用晚期胚胎的组织进行的聚合实验,显示同样情况。形成心脏、软骨、肝脏、视网膜的细胞混合之后,起初可以毫无选择地团在一起,然后各自选择性地聚起来,形成各自应该形成的组织,并且各自在表面或者内部占据各自应有的位置。这些结果说明,不但每种细胞相互之间能粘着,而且和其他类型的细胞之间,还具有一种复杂的、和完整的胚胎中相同的关系。
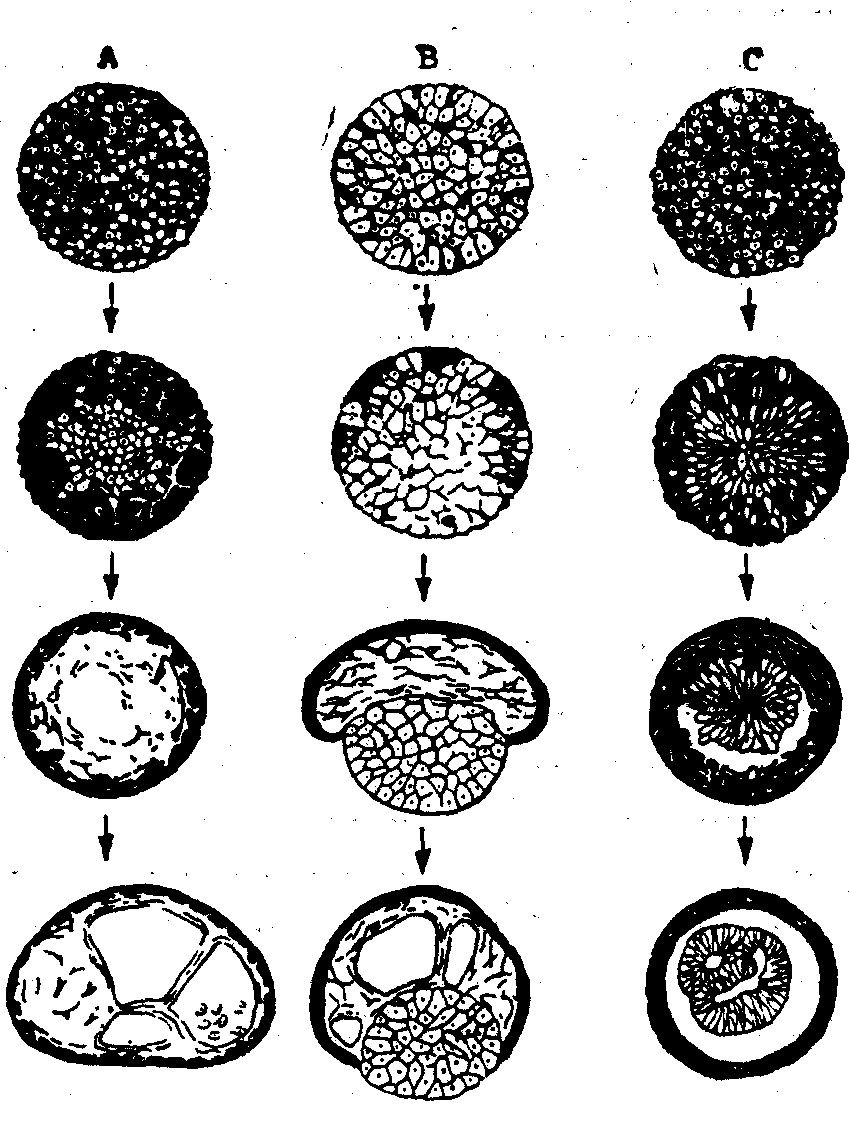
图6 分散后重聚的胚胎细胞的重新排列
A. 混合的上皮细胞和中胚层细胞;B.混合的上皮、中胚层和内胚层细胞; C.混合的上皮和神经板细胞
用鸡胚和鼠胚为材料进行的重聚合的实验,还说明另外一些问题。预先使鸡胚细胞和鼠胚细胞分别聚合,然后使分散的鸡胚细胞或鼠胚细胞在这样的细胞团上粘附,同种细胞的粘附,远比异种细胞为多,说明粘附有种间的差别,有一定的属专一性。但是,如果使鸡胚和鼠胚的视网膜神经细胞(这两种细胞的核可以根据组织学特点加以区分)相混,它们可以保持在一起,形成镶嵌的组织,以后也不再分离,说明不同种属的相同组织之间有组织的专一性,在种属专一性和组织专一性之间,大多数情况后者强于前者,所以不同种的细胞可以形成嵌合组织。
在细胞“识别”并和同种细胞聚合的现象中,表面分子可能起重要的作用。胚胎细胞的表面有组织专一分子的存在,主要是用免疫学的方法证实的。用视网膜神经细胞制备的抗血清,经过同种其他细胞的吸附,专一地和视网膜神经细胞起反应。同样,用肝细胞制备的抗血清专一地和肝细胞起反应。已经知道,专一的视网膜神经细胞聚合因子是一种糖蛋白,分子量约50 000;使肝细胞聚合的因子是低分子量的 (<1 000)。在另外一些实验中则发现一种分子量为140 000的细胞,粘性分子(CAM)对所有神经细胞的聚合和分离均起重要作用,而且,可能它在正常形态建成中也起一定作用。
细胞间的相互作用是各式各样的,可以是单方面的诱导或抑制作用,也可以是细胞间的相互影响。就作用方式来说,有的细胞分化需要细胞间的直接接触,另一些所需要的可能是在一定范围内扩散的化学物质。尤其在需要直接接触的情况,通过细胞的移动可以使不同的细胞遇到一起,彼此发挥应有的作用。
- 宽城满族自治县是什么意思
- 宽甸满族自治县是什么意思
- 宽胸利膈丸是什么意思
- 宾之初筵是什么意思
- 宾县是什么意思
- 宾川县是什么意思
- 宾得R-100系列全站仪是什么意思
- 宾阳县是什么意思
- 宿州市是什么意思
- 宿建德江(1)是什么意思
- 宿松县是什么意思
- 宿根福禄考是什么意思
- 宿根花卉施肥是什么意思
- 宿洞宵宫(1)是什么意思
- 宿济州西门外旅馆(1)是什么意思
- 宿渔家是什么意思
- 宿灵鹫禅寺(其一)(1)是什么意思
- 宿甘露僧舍(1)是什么意思
- 宿西山精舍(1)是什么意思
- 宿迁市是什么意思
- 寄内(1)是什么意思
- 寄外舅郭大夫(1)是什么意思
- 寄小儿是什么意思
- 寄江南故人是什么意思
- 寄洪与权(1)是什么意思
- 寄生性鱼病标本收集是什么意思
- 寄生性鱼病的来源和传播方式是什么意思
- 寄生性鱼病的特点是什么意思
- 寄生杂草是什么意思
- 寄生虫学检验是什么意思
- 寄生虫性皮肤病是什么意思
- 寄生虫病的防治是什么意思
- 寄苏内翰(1)是什么意思
- 寄衣曲(三首)是什么意思
- 寄隐居士(1)是什么意思
- 寄题广信军四望亭(1)是什么意思
- 寄黄几复是什么意思
- 寄黄几复是什么意思
- 寄黄几复(1)是什么意思
- 密云县是什么意思
- 密封性检验是什么意思
- 密尔顿,J.是什么意思
- 密山市是什么意思
- 密度是什么意思
- 密度、相对密度和浓度的概念及其单位是什么意思
- 密度单位换算系数表是什么意思
- 密度和相对密度的测定是什么意思
- 密度的调整是什么意思
- 密环菌是什么意思
- 密苏里妥协案是什么意思
- 密茨凯维支是什么意思
- 密蒙花是什么意思
- 密蒙黄是什么意思
- 密螺旋体是什么意思
- 密达是什么意思
- 密达是什么意思
- 密陀僧是什么意思
- 寇准是什么意思
- 寇松线是什么意思
- 寇松,G.N.是什么意思