红细胞血型抗原的遗传
红细胞血型抗原的遗传
红细胞血型抗原的遗传是免疫遗传学的经典内容。血型抗原都由特异性抗血清检出。所用最主要的血清学方法是红细胞凝集试验。血型抗体有完全抗体和不完全抗体之分。完全抗体可使具有对应抗原的红细胞在盐水悬液和蛋白质胶体悬液中发生凝集。不完全抗体则只能使具有对应抗原的红细胞在蛋白质胶体悬液中发生凝集,或在抗人球蛋白试验中发生凝集。血型抗体又有天然抗体和免疫抗体之分。天然抗体为完全抗体,大半限于IgM,但有些抗体虽属天然抗体,却为不完全类型。免疫抗体为不完全抗体,限于IgG。对某一血型抗原来说,完全抗体和不完全抗体的特异性是一致的。
自1900年Landsteiner发现ABO血型以来,迄今已检出15~17个红细胞血型系统(表1)。其中MNSs血型、Kell血型,可能还有Rh血型为复合系统,即受控于2个或3个密切连锁的位点,其他血型系统刚受控于单
表1 人体红细胞血型抗原遗传

ABO血型系统 1900年Landsteiner取6份血样,将各人的红细胞和血清做交叉凝集试验,发现三种反应格局,分别称之为A型、B型和O型。他推断在人体红细胞表面,A型者有A抗原,B型者有B抗原,O型者两者俱缺; 在人体血清中则有针对本身红细胞所缺抗原的天然凝集抗体,A型者血清中有β凝集素(抗B),B型者血清中有α凝集素(抗A),O型者血清中有α和β凝集素。1902年,Decastello和Sturli发现有人红细胞兼具A抗原和B抗原,血清中无α和β凝集素,这些人属于AB型。因此用抗A和抗B两种抗血清可检出ABO血型中A、B、AB、O四种表型:红细胞在抗A血清中凝集者为A型,在抗B血清中凝集者为B型,在抗A和抗B血清中都凝集者为AB型,在抗A和抗B血清中都不凝集者为O型(表2)。
表2 ABO血型系统的表型和基因型
| 红细胞凝集反应 | 表型 | 基因型 |
| 抗A 抗B | ||
| + - - + + + - - | A型 B型 AB型 O 型 | IAIA,IAi IBIB,IBi IAIB ii |
1924年Bernstein揭示ABO血型系统受控于有三个等位基因的单一位点。IA和IB为共显性,两者对i均为显性,i为隐性。这三个等位基因构成六种基因型和四种表型:纯合体IAIA和杂合体IAi为A型,纯合体IBIB和杂合体IBi为B型,杂合体IAIB为AB型,纯合体ii为O型(表3)。
表3 我国五民族ABO血型系统的表型频率和基因频率(上海市中心血站)
| 民族 | 调查 人数 | 表型频率 | 基因频率 | |||||
| A | B | O | AB | IA | IB | i | ||
| 汉族 | 40980 | 0.3131 | 0.2806 | 0.3086 | 0.0977 | 0.2325 | 0.2117 | 0.5558 |
| 维吾 尔族 | 1513 | 0.2922 | 0.3192 | 0.2750 | 0.1136 | 0.2284 | 0.2468 | 0.5248 |
| 壮族 回族 | 1487 1355 | 0.2125 0.2723 | 0.2757 0.2834 | 0.4728 0.3594 | 0.0390 0.0849 | 0.1352 0.1983 | 0.1727 0.2052 | 0.6921 0.5965 |
| 哈萨 克族 | 885 | 0.2282 | 0.2983 | 0.3797 | 0.0938 | 0.1645 | 0.2072 | 0.6283 |
A抗原和B抗原不仅存在于红细胞表面,也存在于胃、肠、毛发、骨胳等其他组织中。有些人在唾液、胃液、胆汁、精液等分泌物中也有A抗原和/或B抗原,称为分泌型。在分泌物中没有A抗原和B抗原的人称为非分泌型。分泌型和非分泌型受控于另一对等位基因Se和se。基因型Se Se和Se se为分泌型,基因型se se为非分泌型。
A抗原和B抗原的出现需以存在H抗原为前提。i基因为无效等位基因,O型红细胞有H抗原但不能由此产生A抗原或B抗原。因此ABO血型有时称为ABH血型。1952年,Bhende在印度孟买首次发现一种特殊O型,这种红细胞既无A抗原和B抗原,也无H抗原,称为孟买型。这表明存在一对等位基因H和h。基因型HH和Hh能产生H抗原,由此再经IA和IB基因的作用而出现A抗原和B抗原。基因型hh不能产生H物质,即使有IA和IB基因也无从形成A抗原和B抗原。Hh位点和ABO位点是彼此独立的,因此孟买型可按有无IA和IB基因而记为Oh、OAh、OBh、OABh。
红细胞膜上的血型物质是脂溶性的糖脂,分泌物中的血型物质是水溶性的糖蛋白。两者糖链的主链的化学结构特点如下:
❶红细胞膜上的血型物质:D-半乳糖—N-乙酰葡糖胺—D-半乳糖—葡萄糖—鞘脂;
❷分泌物中的血型物质: D-半乳糖—N-乙酰葡糖胺—D-半乳糖—N-乙半乳糖胺。对水溶性血型物质的分析表明,糖蛋白的肽链赋予血型物质以抗原性,末端多糖赋予血型物质以特异性。A物质和B物质经过各自的特异性酶降解后,由A物质可得H物质和N-乙酰半乳糖胺,由B物质得H物质和半乳糖。两者差别仅在一个单糖残基。H物质可进一步分解为Ⅰ型和Ⅱ型前体链及岩藻糖。现知H基因的产物是α-L-岩藻糖转移酶(1),此酶催化在Ⅰ型和Ⅱ型前体链的末端半乳糖残基上接上一个岩藻糖,由此构成H物质。IA基因的产物是α-N-乙酰半乳糖胺转移酶,经此酶催化,再在H物质末端半乳糖残基上接上一个N-乙酰半乳糖胺,由此构成A抗原特异性。IB基因的产物是α-D-半乳糖转移酶,经此酶催化,再在H物质末端半乳糖残基上接上一个D-半乳糖,由此构成B抗原特异性(图1)。在孟买型中,Oh者缺乏α-L-岩藻糖转移酶(1),因而不能形成H物质,OAh、OBh、OABh虽有相应受控于IA、IB基因的酶,但因为无作用底物,所以只能成为孟买型。
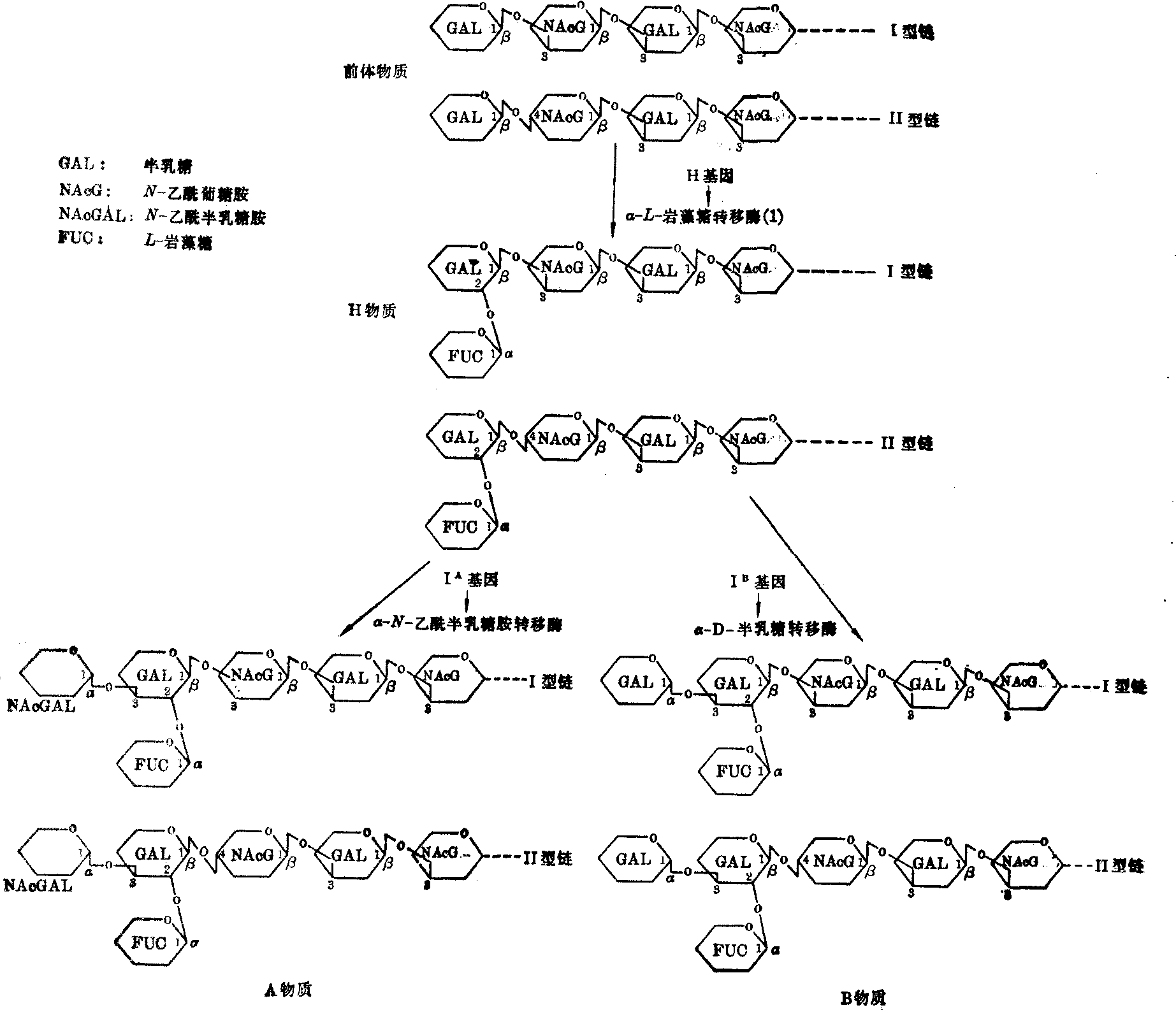
图1 水溶性A物质和B物质的形成
MNSs血型系统 1927年,Landsteiner等以兔抗人红细胞血清检出红细胞抗原M和N,受控于一对等位基因M和N,计有三种基因型MM、MN和NN,决定三种表型M(+),M(+)N(+),和N(+)。1947年,Walsh等在一新生儿溶血症患儿的母血中发现抗S抗体; 1951年,Levine等找到抗S抗体,两者检出的抗原受控于另一对显性基因S和隐性基因s。MN位点和Ss位点密切连锁,组成四种基因复合体MS、Ms、NS和Ns,由此产生十种基因型和六种表型(表4)。
据上海市中心血站报道,我国人(民族不详)Ss血型的表型频率为S(+)0.0863,s(+)0.9137(表5)。
P血型系统 1927年,Landsteiner等用兔抗人红细
表4 MNSs血型系统的表型和基因型
| 红细胞凝集反应 | 表 型 | 基因型 | |||
| 抗M | 抗N | 抗S | 抗s | ||
| + + + + - - | - - + + + + | + - + - + - | - + - + - + | M(+)S(+) M(+)s(+) M(+)N(+)S(+) M(+)N(+)s(+) N(+)S(+) N(+)s(+) | MS/MS,MS/Ms MS/Ms MS/NS,MS/Ns,Ms/Ns Ms/Ns NS/NS,NS/Ns Ns/Ns |
表5 我国五个民族MN血型的表型频率和基因频率(上海市中心血站)
| 民族 | 调查人数 | 表型频率 | 基因频率 | |||
| M(+) | N(+) | M(+)N(+) | M | N | ||
| 汉 族 维吾尔族 壮 族 回 族 蒙 族 | 1788 1513 1170 1355 1112 | 0.2220 0.3893 0.5487 0.2516 0.2959 | 0.2964 0.1395 0.0821 0.2502 0.2302 | 0.4816 0.4712 0.3692 0.4982 0.4739 | 0.4628 0.6249 0.7333 0.5007 0.5329 | 0.5372 0.3751 0.2667 0.4993 0.4671 |
表6 P血型系统的表型和基因型
| 抗P | 抗Tja | 表型 | 基因型 |
| + - - | + + - | P1 P2 p | P1P1;P2P2;P1p P2P2;P2p pp |
表7 我国五个民族P血型系统的表型频率和基因频率(上海市中心血站)
| 民族 | 调查人数 | 表型频率 | 基因频率 | ||
| P(+) | P(-) | p1 | P2 | ||
| 汉 族 维吾尔族 壮 族 回 族 蒙 族 | 1619 1513 1487 1355 1112 | 0.3428 0.5757 0.2334 0.5343 0.4839 | 0.6572 0.4243 0.7666 0.4657 0.5161 | 0.1893 0.3486 0.1244 0.3176 0.2816 | 0.8107 0.6514 0.8756 0.6824 0.7184 |
Rh血型系统 1940年,Landsteiner和Wiener发现约85%白人的红细胞可为兔抗猕猴红细胞血清所凝集。当时认为有些人的红细胞有与猕猴共有的抗原决定簇,称之为Rh因子。在这之前一年,Levine和Stetson报道了一个O型产妇娩出死胎,又在输入其O型丈夫血时发生严重输血反应,此产妇血清可使104例O型红细胞中83例发生凝集,后来证明此产妇血清的特异性与兔抗猕猴红细胞血清相同。紧接着Race等(1943,1944),Levine(1945),Mourant(1945)相继发现人体血清可检出属于Rh血型系统的另外四种特异性。因此检测Rh血型共有五种抗血清。
对Rh血型系统的遗传有下列两种学说,至今还不能肯定谁是谁非(图2)。
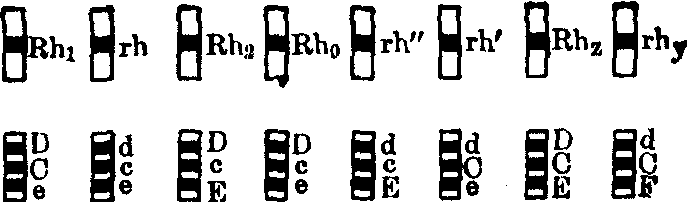
图2 Rh血型位点的命名
上行示Weiner命名系统 下行示Fisher-Race命名系统
Fisher和Race认为,Rh血型系统决定于三个密切连锁的独立位点,各有一对等位基因:C和c;D和d;E和e。五种Rh血清称为抗C、抗c、抗D、抗E和抗e;至今未曾发现抗d血清,d仅表示无D抗原。D是最初发现的,也是最重要的Rh抗原。D阳性即Rh阳性,D阴性即Rh阴性。这三对连锁基因组合成8种Rh基因复合体:cDe、CDe、cDE、cde、Cde、cdE、CDE、CdE。Fisher认为,这三个位点在染色体上的位置是D-C-E。虽然Fisher假设稀有基因复合体CDE和cdE来自交换,但事实上至今未曾发现发生交换的家系。
Wiener认为,Rh血型系统决定于单一位点上的8个等位基因: Rh0、Rh1、Rh2、rh、rh′、rh″、Rhz、rhY。每个等位基因控制一种凝集原,它有若干抗原决定簇或称抗原因子,各自可为特异血清检出。
上述两种学说命名的相互关系见表8
表8 Rh血型系统Fisher-Race和Wiener命名对照
| Rh基因复合体 (Fisher-Race命名) | Rh等位基因:抗原因子组合 (Wiener命名) |
| cDe CDe cDE cde Cde cdE CDE CdE | Rh0(R0):hr′,Rh0,hr″ Rh1(R1):rh′,Rh0,h″ Rh2(R2):hr′,Rh0,rh″ rh(r):hr′,hr″ rh′(r′):rh′,hr″ rh″(r″):hr′,rh″ Rh2(R2):rh′,Rh0,rh″ rhY(rY):rh″,rh″ |
C=rh′,c=hr′,D=Rh0,E=rh″,e=hr″
8种Rh基因复合体可构成36种基因型。表型ce只有cde/cde一种基因型。表型CcDEe有6种可能基因型,其确切基因型需由家系数据推导,由群体Rh基因复合体频率可估计最可能的是哪一种(表9)。
表9 我国四个民族的Rh基因复合体频率(上海市中心血站)
| 民族 | cDe (R0) | CDe (R1) | cDE (R2) | cde (r) | Cde (r′) | cdE (r″) | CDE (R2) | CdE (rY) |
| 汉族 壮族 回族 蒙族 | 0.0285 0.0268 0.0526 0.0643 | 0.6466 0.7962 0.5880 0.5906 | 0.2664 0.1173 0.2608 0.2719 | 0.0447 0.0367 0.0719 0.0521 | 0.0092 0.0152 0.0140 0.0152 | 0.0037 0.0000 0.0000 0.0000 | 0.0090 0.0078 0.0127 0.0059 | 稀 有 |
Kell血型系统 1945年Coombs等利用抗人球蛋白抗体使被不完全Rh弱抗体致敏的红细胞发生凝集。这就是免疫血液学中著名的抗人球蛋白试验,或称Coombs试验。在将此试验应用于临床几周之内,Coombs(1946)就在Kell夫人的血清中发现一种特殊抗体,称为抗K。1949年Levine等在新生儿溶血症母血中发现对应的抗K。Allon等人相继于1957年和1958年发现属于Kell复合体的抗Kpa和抗Kpb。接着Giblett(1958)和Walker等(1963)检出了与其他血型基因独立分离但与Kell复合体连锁的Jsa和Jsb,14~20%的美国黑人有之,其他人种罕见。Kell血型系统是一个受控于2~3个密切连锁位点的复合系统。家系材料证明K位点和Kp位点密切连锁,可检出基因复合体KKpb、kKpa和kKpb,但迄今未曾检出KKpa。已知Jsa和Jsb受控于复合Kell位点,白人中Kell血型组合最常见的是kKpbJsb,其次是KKpbJsb和kKpaJsb。Kell血型系统K和k抗原的免疫原性仅次于Rh血型的D抗原,在白人中与ABO、Rh血型一起被列为三大血型。
Lewis血型系统 1946年,Mourant发现Lewis血型系统的抗Lea抗体,1948年Andresen找出抗Leb抗体。表型主要是Le (a+b-)、Le(a-b+)和Le(a-b-),有些抗Leb血清加上敏感的技术偶可检出Le(a+b+),但有人认为表型Le(a+b+)是不存在的。Lewis血型的表现较为复杂,与A、B、H血型物质的分泌特性有密切关系。所有Le(a+)者均为A、B、H血型物质非分泌者,但在他们的唾液中有可溶性Lea抗原。另一方面,并非所有非分泌者都是Le(a+),有些可为Le(a-b-)。在非分泌者的唾液中很少有Leb抗原。此外还发现,彻底冲洗红细胞可以完全消除与抗Lea和抗Leb的反应性;相反,无Lea和Leb抗原的红细胞接触Le(a+)血浆,可转变为Le(a+)细胞; 接触Le(b+)血浆,可转变为Le(b+)细胞。在将Le(a+b-)血液输入于Le(a-b+)受者时,可检出供者红细胞转变为Le(a+b+)。于是,有人提出ABH和Lewis血型抗原受控于ABO、H、Se和Le四个独立位点,它们相互作用导致分泌物中出现适当的抗原,然后Lewis抗原再吸附到红细胞膜上,因此红细胞的表型取决于血浆中每种抗原的量。分泌物中有无Lea物质,决定于一对等位基因Le和le。基因型LeLe和Lele为Le(a+),lele为Le(a-)。现知Lea和Leb物质的出现,均以Ⅰ型前体链为前提。在第二个单糖残基N-乙酰葡糖胺上接上一个L-岩藻糖,构成Lea特异性。H基因和Le基因的产物〔α-L-岩藻糖转移酶(1)和(2)〕共同作用于前体链,分别催化在其末端半乳糖残基和末端第二位N-乙酰半乳糖胺残基上接上L-岩藻糖,这时就构成Leb特异性。因此Leb特异性是H基因和Le基因共同作用的产物(图3)。
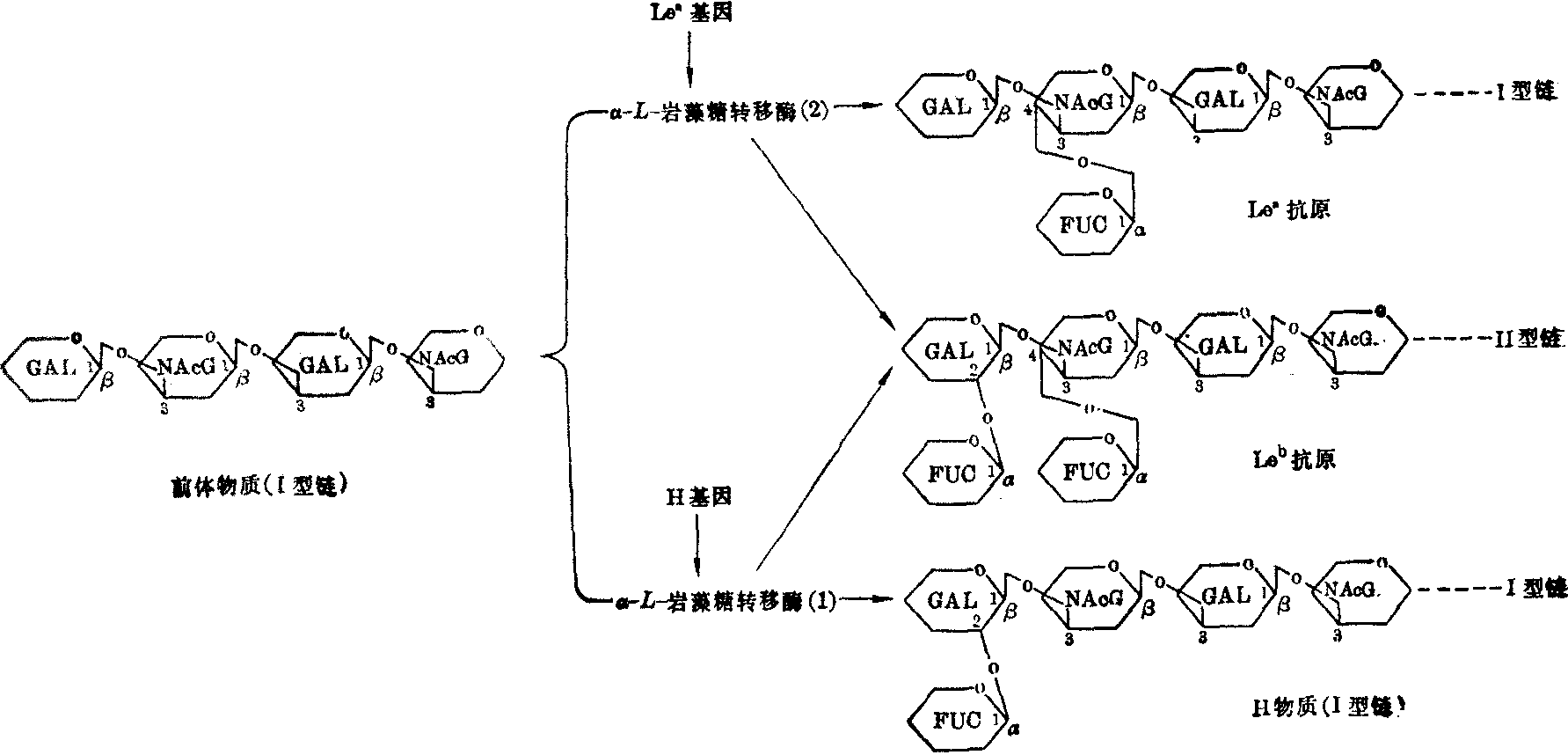
图3 H物质和Lewis抗原的形成
Xg血型系统 Xg是迄今所知唯一的X连锁遗传血型系统。1962年,Mann等在为一家族性毛细血管扩张症鼻出血患者作配血试验时,发现一种特异性血型抗体抗Xga,以此抗血清随机检查342个个体,发现表型Xg(a+)和Xg(a-)的分布有性别差异:

系谱分析证明Xga抗原受控于X染色体上Xga基因,其无效基因称Xg。Xg(a+)男性与Xg(a+)女性婚配。女儿不可能是Xg(a-),在有的家庭中可出现儿子为Xg(a-)。Xg(a+)男性与Xg(a-)女性婚配,儿子全为Xg(a-),女儿全为Xg(a+)。Xg(a-)男性与Xg(a+)女性婚配,儿子和女儿均有Xg(a+)或Xg(a-)的可能。
高频和低频血型系统 在已经鉴定的血型系统之外,还有多种抗血清检出的一系列不同的血型抗原,可分为高频和低频两大类。高频或“公共”血型抗原见于绝大多数人的红细胞(如Csa、Gya、Ata、Yka、Sda、Gna、Jra),低频或“私有”血型抗原仅见于少数人的红细胞,甚至仅见于特殊家系(如Bea、Bya、Chra、Swa、Tra、Pra、Bwa等)。
血型不相容 临床溶血性输血反应和新生儿溶血症的原因都是两个个体的红细胞血型不相容或称不配合。事实上很多特异性抗体是在输血反应病人和新生儿溶血症母血中检出的。由于可能遇到某种未知的血型抗体,输血前必作常规交叉配血试验以确保安全。
新生儿溶血症是由于胎儿由父源染色体决定的血型抗原恰为母亲所缺。在胎儿红细胞通过胎盘出血进入母体循环时,母体即被致敏而产生免疫性抗体,此种免疫性抗体可通过胎盘屏障进入胎儿血流,导致新生儿溶血症(胎儿有核红细胞增多症)。
理论上说,凡母体因胎母不合而产生免疫性IgG者均可导致新生儿溶血症,除ABO和Rh血型抗体引起者外,曾报道Kell血型系统的抗K、抗k、抗Kpa、抗Kpb,Duffy血液系统的抗Fya,MNSs系统的抗S和抗M,Kidd系统的抗Jka和抗Jkb,Xg系统的Xga都可引起轻重不一的新生儿溶血症。
我国各族人民中,Rh阴性者约占3~8%,而O型者约占20~40%,因此ABO新生儿溶血症较Rh新生儿溶血症多见,但后者症状较为严重。上海市中心血站统计308例新生儿溶血症,ABO不相容者占63%,Rh不相容者占36%。
☚ 免疫遗传学 主要组织相容性复合体 ☛
- 课外阅读是什么意思
- 课外阅读指导是什么意思
- 课外音乐欣赏活动是什么意思
- 课外音乐活动是什么意思
- 课外音乐活动的任务是什么意思
- 课外音乐活动的原则是什么意思
- 课外音乐活动的因才施教性是什么意思
- 课外音乐活动的灵活性是什么意思
- 课外音乐活动的群众性是什么意思
- 课外音乐活动的自愿性是什么意思
- 课外音乐活动的趣味性是什么意思
- 课夫是什么意思
- 课头是什么意思
- 课子是什么意思
- 课子堂是什么意思
- 课孙草是什么意思
- 课学是什么意思
- 课安是什么意思
- 课室是什么意思
- 课工是什么意思
- 课巾是什么意思
- 课弟苦读 钱陈群是什么意思
- 课录是什么意思
- 课役是什么意思
- 课役户是什么意思
- 课征是什么意思
- 课徒是什么意思
- 课忠堂是什么意思
- 课息是什么意思
- 课户是什么意思
- 课户与不课户是什么意思
- 课托儿是什么意思
- 课捕是什么意思
- 课改:走在语文课例研修的路上是什么意思
- 课效是什么意思
- 课敛是什么意思
- 课文是什么意思
- 课文作业是什么意思
- 课文分析是什么意思
- 课文类型是什么意思
- 课文背诵是什么意思
- 课时是什么意思
- 课时补贴是什么意思
- 课时计划是什么意思
- 课时计划的作用与编写是什么意思
- 课易是什么意思
- 课易存商是什么意思
- 课最是什么意思
- 课本是什么意思
- 课本剧是什么意思
- 课材是什么意思
- 课核是什么意思
- 课桌是什么意思
- 课桌文化是什么意思
- 课桌文学是什么意思
- 课桌椅卫生是什么意思
- 课桌椅尺寸是什么意思
- 课桌椅标准是什么意思
- 课桌椅的卫生要求是什么意思
- 课桌椅距离是什么意思