病毒复制
病毒的复制也称为增殖,因为病毒的基本要素——核酸是通过复制的方式增殖的。病毒的复制是病毒学研究的中心问题之一,是与研究病毒如何感染,如何致病等问题有着密切关系的。
病毒的增殖和细菌不同,不是有丝分裂,而是复制,好象照像一样,先拍出一种反体的物象,和原来的图象是立体异构的,称为底板或“负本”,从这个“负本”再冲洗 (复制)出这个“负本”的立体异构物,即和原来物体完全相同的物体或“正本”。复制可以反复进行多次。
各种病毒的复制原理基本相同,但是,由于所含核酸类型的不同而有所差异(参见“病毒核酸”)。正链RNA病毒、负链RNA病毒、双链RNA病毒及双链DNA病毒等的具体复制程序都有各自的特殊性。
病毒的RNA或DNA进入细胞后,一般先进行信息核糖核酸(mRNA)的合成,这个过程叫做转录。mRNA的转录是由一种多聚酶(也叫转录酶)来完成的。有些病毒颗粒内含有自身的转录酶,如正粘病毒、副粘病毒及痘病毒等,能以病毒RNA或DNA为模板转录自身的mRNA。有些病毒不具备这种酶,须依靠细胞的依赖于DNA的RNA多聚酶来转录病毒的mRNA,如乳多泡病毒和腺病毒等。还有一些病毒,其RNA本身就具有mRNA的性质,不需要先进行转录,如小RNA病毒和披膜病毒等。病毒的转录以其RNA或DNA基因组上的有意义密码子,依照Watson-Crick的碱基配对原则合成mRNA。如下式所示:
DNA上有意义密码子 3'TACAACAAA5'm
RNA密码子 5'AUGUUGUUU3'
转录成的mRNA便可以进行病毒蛋白的转译或翻译(即合成蛋白)。
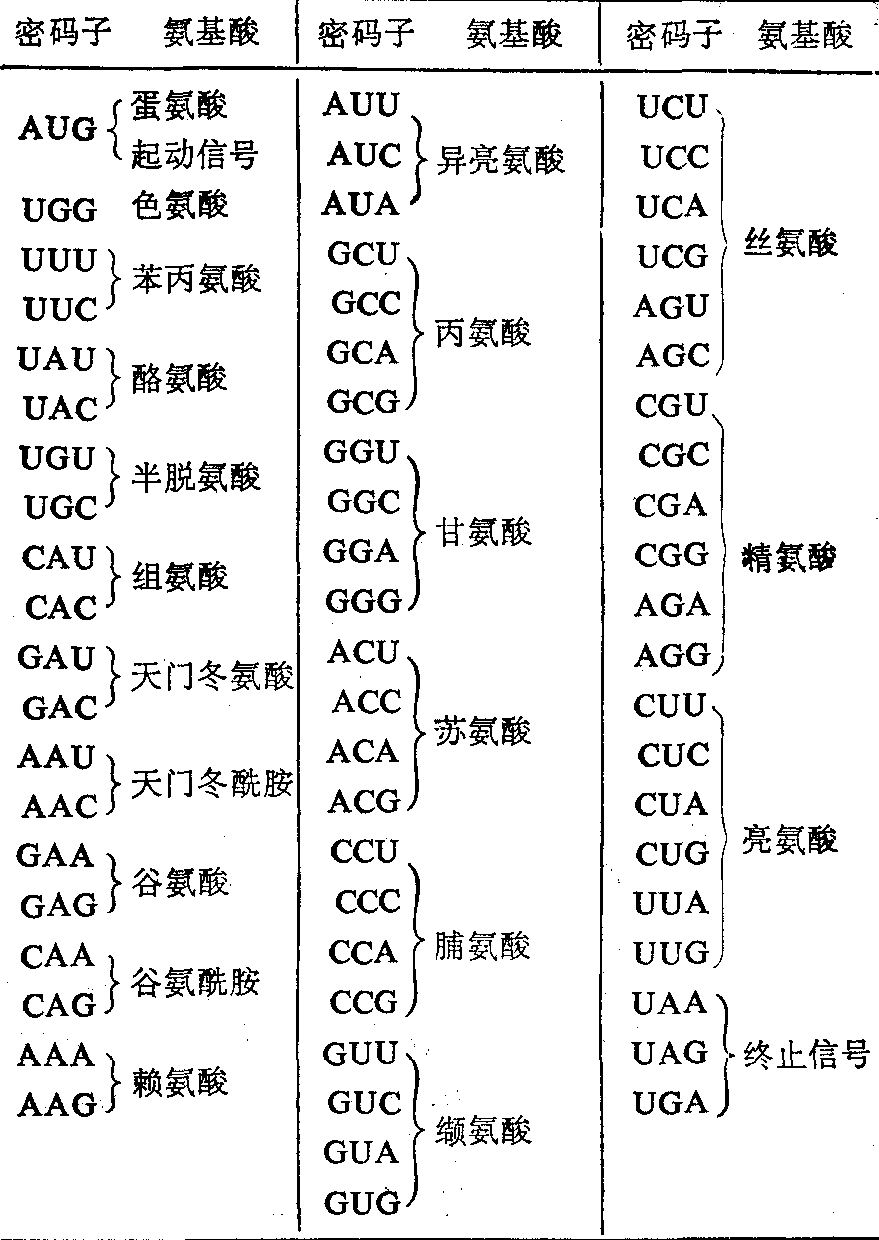
蛋白质的转译是按照三联密码进行翻译的(表)。
转译的蛋白质有的最后结合到新病毒颗粒中去,称为结构蛋白。有的虽不结合到病毒颗粒中去但对病毒的复制具有特殊作用,称为非结构蛋白。
由于病毒mRNA转译的结果,合成了另一种病毒的多聚酶,又称为复制酶,开始进行病毒RNA或DNA的复制。不论病毒RNA或DNA的复制都以原来的病毒RNA或DNA为模板。按照Watson-Crick的碱基配对原则合成与原来RNA或DNA相互补的链,称为互补链 (cRNA或cDNA)。如果原来链上的密码是有意义密码,复制的互补链上的密码即为无意义的,反之亦然。如下式所示:原来DNA链(无意义密码子) 5'ATGTTGTTT3'复制互补DNA链(有意义密码子) 3'TACAACAAA5'不论是有意义的密码子或无意义密码子都可以被复制,但是只有有意义密码子才能被转录为mRNA。
mRNA链上氨基酸的三联密码子及起动和终止信号
当细胞内病毒的核酸和蛋白达到一定的浓度后,即按其特殊结构结合起来。这个过程叫做装配或组装,并成熟为新的病毒颗粒即子代病毒。
病毒的整个复制周期,一般包括下列几个阶段:吸附和穿入;脱壳;转录病毒mRNA、转译病毒蛋白和复制病毒DNA或RNA;装配、成熟和释放。病毒复制的各个阶段不能截然分开。
吸附和穿入 病毒吸附或粘着在敏感细胞表面上是病毒进行复制的第一步。吸附过程一般可分为两个阶段。首先,病毒由于分子运动和细胞相互碰击而与敏感细胞接触,继而因细胞与病毒颗粒之间的静电吸引作用或范德瓦耳斯(Von den Waals)吸附作用相互吸引。这种吸附一般不依赖于温度,在细胞表面上也没有一定的部位。这个阶段是可逆的。换句话说,这时还可以从细胞回收完整的、仍然具有感染性的病毒颗粒。
以后吸附过程很快进入到第二个阶段——不可逆阶段。由于大多数病毒颗粒表面具有特殊酶活性的蛋白,例如小RNA病毒的一种称为VP4的蛋白,正粘病毒的血凝素(HA),副粘病毒的具有血凝素及神经氨酸酶活性的一种粘蛋白(HN),弹状病毒的G蛋白,腺病毒的六邻体等与细胞上特异的受体的亲和作用,形成更紧密的结合,发生化学变化。病毒的酶分解细胞受体的结果,使局部细胞膜的通透性发生改变,病毒与细胞的胞膜接触使病毒的包膜损坏,便造成病毒与细胞膜融合,因此成为吸附的不可逆阶段。
有些病毒对细胞受体的要求十分严格,而另一些病毒则相反。例如,脊髓灰质炎病毒只能被灵长类动物的细胞受体所吸附,而不能被啮齿类动物细胞所吸附。流感病毒、副流感病毒则不甚严格,能吸附于多数鸟类及哺乳类动物的细胞上。
病毒进入敏感细胞的机理有两种不同情况。一种是病毒包膜与细胞膜由于酶的作用遭受某种程度的降解而互相融合,病毒即进入细胞。这种过程叫做穿入。另一种是吞饮,病毒接触并附着于细胞表面后被细胞吞饮到空泡内。严格说来,这时病毒还处于细胞外。然后病毒仍须进一步和细胞质膜相融合。小RNA病毒、副粘病毒、正粘病毒、痘病毒、疱疹病毒、腺病毒等都有吞饮和穿入的实验报告。
脱壳 病毒感染的第二步是在细胞质内脱去包围病毒核酸(或核蛋白)的外壳,使病毒核酸(或核蛋白)能够在细胞内裸露并进行复制。病毒的脱壳过程在某些病毒很难和吸附阶段的后期截然分开。
小RNA病毒,有包膜病毒的脱壳过程比较简单,但目前还没有足够的实验资料说明其机理。然而,从它们的增殖速度来看,病毒和细胞融合的后期,病毒的RNA便游离到细胞内,它们的脱壳过程很短。
有些病毒如痘病毒和呼肠孤病毒不需要完全脱壳,转录过程就开始进行。另一些单链负链RNA病毒如正粘病毒、副粘病毒等在完全脱去核壳之后,才开始转录。
痘病毒的脱壳过程比较复杂。痘病毒具有包膜和外壳,在敏感细胞中由于细胞-病毒相互作用而解除包膜时,可能解离出一种对紫外线敏感的蛋白。这种蛋白能够抑制宿主细胞中另一种蛋白。后者的作用是封闭宿主细胞中原来存在的能产生脱壳酶的基因。
病毒基因的转录、转译和复制
(1) DNA病毒: 这类病毒可分为三类。
第一类是腺病毒、乳多泡病毒及疱疹病毒。其IUR(开始复制的单位) 是双链DNA(±DNA)。腺病毒和疱疹病毒是双链线状DNA,乳多泡病毒是双链环状DNA。这类病毒一部分DNA带着有意义的可供转录mRNA的密码子,另一部分带着无意义的密码子,不能转录mRNA。这类病毒颗粒内未证明含有RNA多聚酶。因此,病毒mRNA的转录只能利用细胞内的依赖于DNA的RNA多聚酶来完成,其转录及复制均在细胞核内进行。腺病毒和疱疹病毒从双链DNA中每一股的一部分(带有意义的密码子部分)转录成mRNA。乳多泡病毒如SV40则从每股DNA的一半(带着有意义的密码子)转录mRNA。这类病毒只能在胞核内进行复制。
第二类是痘病毒。其IUR是线状双链脱氧核蛋白(±DNP)。痘病毒颗粒内含有RNA多聚酶以及胸腺嘧啶激酶和多核苷酸连接酶等,所以不需要细胞的RNA多聚酶来转录病毒的mRNA。病毒mRNA又转译了病毒DNA多聚酶以复制病毒DNA。以前认为,痘病毒全部复制周期包括转录都在胞质内进行。但最近的实验证明,痘病毒同时也能在胞核内进行复制。
第三类是小DNA病毒。其IUR是+DNP (有些是-DNP。只有当人工抽提时其核酸才成为双链的±DNA形式)。小DNA病毒可分有缺陷的和无缺陷的两种。有缺陷的小DNA病毒在复制时必须同时有其他病毒的帮助才能生成完整病毒颗粒,如腺病毒伴随病毒。另一种是无缺陷的小DNA病毒 (如人H-3病毒),在复制时不需要其他病毒的帮助,但却需要细胞在生长的静止期的功能给以援助。这类病毒只在细胞核内进行复制。
从病毒基因组 (在DNA病毒是病毒的±DNA或-DNA)转录mRNA(±mRNA或+mRNA)时必须由依赖于DNA的RNA多聚酶来启动。这种起转录作用的多聚酶又称转录酶。依赖于DNA的RNA多聚酶能识别基因组上一定的核苷酸序列而与之相结合才能开始转录,这种序列又称为启动子(参见“病毒核酸”)。由±DNA转录的mRNA的极性均为正的,为方便起见称为±mRNA。
在启动子序列后面接着是开始转录mRNA的信号——起始序列。同一病毒的不同蛋白基因的起始序列的核苷酸数目并不一样。例如噬菌体φ×174 A蛋白基因的起始序列共有18个核苷酸,而D蛋白基因的起始序列共有32个。起始序列的长短可能与转录mRNA的速率及效率有关。起始序列之后便是供mRNA开始转译蛋白的信号——如果+DNA的起始信号密码子是ATG,终止信号的密码子是TGA和TAA,那么其互补链DNA的开始密码子相应为TAC,终止密码子为ACT和ATT。
DNA基因组一般有2个以上供转录mRNA的起始信号和相同数目的终止信号。一般在前一个终止信号出现以后始出现另一个起始信号。这种基因组称为不重叠基因组。有时在前一个终止信号未出现之前第二个起始信号就出现了,因而在基因组上有一段核苷酸序列是二种mRNA所共同的,称为重叠基因组。DNA病毒如腺病毒、乳多泡病毒等的基因组都有数个重叠基因。
mRNA的转录还有一种现象叫做mRNA的剪接,即在mRNA开始转录一段长度后,由于在DNA的分子上有一段剪接序列,上面有剪接信号,它以3′GU……开始而以……AG5′结束,而细胞内则有一种小核内RNA(snRNA)能够认别这个信号,因而把剪接序列中的插入部分剪掉,但其转录的密码子可能错位,而与原来不经剪接的mRNA不相同。腺病毒、泡沫病毒和流感病毒的mRNA转录时都有剪接现象。mRNA的转录同时可以又有剪接又有重叠现象。
mRNA一经转录,便由鸟便嘌呤核苷酰转移酶和鸟便嘌呤核苷酸7甲基化酶在5末端加上一个m7G (5′)ppp(5′)Ap或m7G(5)ppp(5)GmpGp的“帽子”,转录终止后又由腺嘌呤核苷酸多聚酶在其3′末端接上一个含50~200个(不同的病毒数目不同)的聚腺嘌呤核苷酸的 “尾”(多聚A)。“帽子”的功能是加强mRNA的转译能力及其转译速率; “尾”的功能是使细胞核内合成的mRNA能够迅速地转移到细胞质中,以便进行转译。
蛋白的转译是从mRNA的起始密码AUG信号开始的。mRNA由基因组的3'末端向5'末端转录,合成的mRNA则由mRNA的5'末端向3'末端进行转译。病毒的mRNA进行转译与动物细胞的mRNA一样,必须在细胞核糖体40s和60s两种亚基的共同作用下按照上述三联密码子转译氨基酸。核糖体含有能识别mRNA的起始位点而与mRNA结合,这些能识别起始位点的细胞成分称为起始因子;其次是蛋氨酰tRNA与mRNA上的起始密码AUG结合以开始肽链的合成; 然后是其他各种氨基酰tRNA含有不同的反密码子和延长因子以进行肽链的延长。当转译进行到mRNA链上的终止密码UAA,UAG或UGA时,细胞的终止因子能识别这种信号,蛋白的转译就停止。释放因子使新合成的蛋白从核糖体上脱离,最后进行转译后加工(如酶解,糖化,碱酸化等)。全部转译过程中所需的能量由细胞供给。
从病毒的mRNA转译用以复制病毒DNA所需的DNA多聚酶(DNA复制酶)后 (有些DNA病毒能利用细胞的DNA多聚酶),就开始用病毒DNA为模板复制新的DNA。双链±DNA同时复制互补的正负两股链,单链+DNA或-DNA先复制互补的-DNA或+DNA链,再复制原来的DNA链。双链±DNA的复制形式,是半保留复制,复制由解链酶带头,将±DNA双链解开成 +DNA及-DNA二股单链,同时复制酶在被解开的链上复制了相互补的-DNA及+DNA两股互补链,合成新的±DNA双链。这样原来的±DNA双链就分为两股,分别分配到二个子代的±DNA双链中。单链DNA的复制则是全保留复制形式。+DNA(或-DNA)模板,复制了其互补的-DNA(或+DNA)互补链,再以互补链为模板复制新的+DNA (或-DNA)单链。这样,原来的单链DNA复制后都完全地保留下来。
病毒DNA在复制开始时,需要一个带有3′末端的引物,在这个引物上,以病毒DNA为模板,从DNA的3'末端起向5'末端复制,被复制的互补DNA即相应地从5'末端开始至3'末端完成。
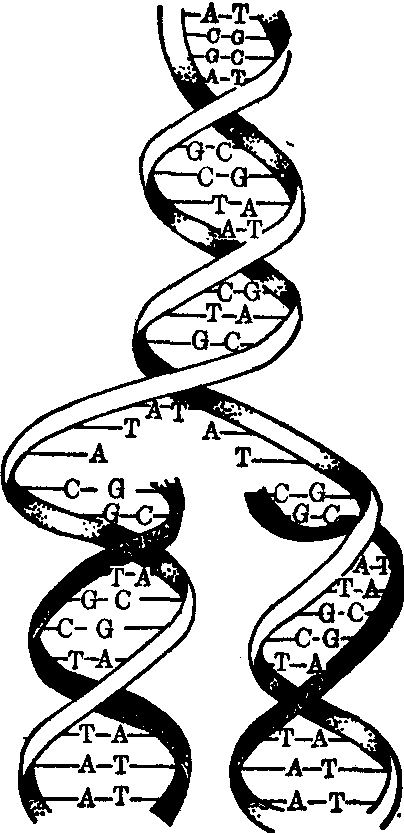
DNA的半保留复制图中黑色带为新生的子代DNA,白色带为亲代DNA。
在整个病毒复制过程(mRNA的转录,蛋白的转译以及病毒DNA的复制)中,都受到一些物质的调控。其中有属于细胞成分的,例如上述起始因子、终止因子等。也有病毒基因编码的特殊酶类,如痘病毒基因编码的RNA多聚酶、多聚A聚合酶、鸟酰转移酶、甲基转移酶及蛋白激酶等都和病毒复制的调控有关。但对这方面的研究尚待深入。
(2) RNA病毒:这类病毒比较复杂,按基因组和复制过程的不同可以分为五类。绝大多数RNA病毒的转录、复制和转译均在细胞质内进行。RNA病毒的mRNA在转译时和DNA病毒一样需要细胞质内的转译系统,并由细胞提供能量。
第一类——含有双链±RNA基因组如呼肠孤病毒。这类病毒的IUR是病毒的核蛋白核心(±RNP)。病毒颗粒含有依赖于RNA的RNA多聚酶。先从原来±RNA中的-RNA转录出+RNA,这种RNA和原来病毒颗粒中的+RNA的5'末端随即被戴上m7GpppGmpG的“帽子”,而成为mRNA。呼肠孤病毒的mRNA是一种不需要多聚A“尾”端的哺乳动物的病毒。
呼肠孤病毒的±RNA是分节段的,可分成10~12个节段,因此它的mRNA也是分节段的。mRNA转译时先转译合成更多的多聚酶 (这种后合成的多聚酶又称为复制酶) 以及一些构成核蛋白的蛋白亚基和其他调控酶类如核苷酸磷酸化酶、鸟便嘌呤转移酶和甲基转移酶等。+RNA随即与一些新生的病毒蛋白形成+RNP,然后作为复制-RNA的模板合成了-RNP,最后产生±RNP,其外层立即包以核壳蛋白,形成病毒颗粒。
呼肠孤病毒在病毒颗粒内含有大约3200个寡核苷酸,都具有pppA-或pppG-的结构,核苷酸的数目从2到20不等。据研究,它们是在后期被转录的不完全RNA节段上。
第二类——正粘病毒。如流感病毒,其IUR是-RNP。这类病毒的-RNA分为8个节段,本身没有感染性,不能起mRNA的作用。其病毒颗粒含有病毒特异的RNA多聚酶,在细胞质中转录的第一步是利用病毒颗粒内的RNA多聚酶以各节段-RNA为模板,转录成相应的+RNA,+RNA迅速被戴上宿主的甲基化“帽子”并接上多聚A尾端成为+mRNA。+RNA有两种功能,一种是作为复制病毒-RNA的模板,另一种是作为mRNA转译相应的病毒蛋白。用病毒的-RNA在试管中进行转录以合成+RNA时,需要一种特异的二核苷单磷酸引物,因此认为,流感病毒转录早期,即转录+RNA阶段需要细胞的依赖于DNA的RNA多聚酶提供一种RNA引物,这可能是流感病毒的复制之所以对于抗DNA合成的药物敏感的原因,流感病毒的基因也有重叠的现象。
第三类——副粘病毒和弹状病毒。其IUR是不分节段的-RNP。这类病毒的-RNA也没有感染性。水泡性口腔炎病毒也属于这一类,由病毒颗粒内的NP蛋白和P蛋白构成RNA多聚酶,催化+RNA的转录。首先转录的是5种长度不同的+RNA,可能是由一起始点转译成大+RNA分子,再裂解为5段。每种+RNA随即被戴上5′端m7G (5′)ppp及5′XYp (X代表m6Am,Y代表A,Am或m6A′m)“帽子”,并接上3′多聚A“尾”端。5种mRNA为5种蛋白编码。当mRNA开始转译了N蛋白 (与RNA组成核蛋白),就开始复制全长的+RNA,与上述合成的蛋白组成核蛋白。因此,如果抑制了N蛋白的转译就导致对+RNA合成的抑制。
仙台病毒及其他副粘病毒的mRNA为6种蛋白编码,其中的F0蛋白与转录的起始有关。F0及NP蛋白以后又进行酶解,衍生了另一些病毒颗粒的结构蛋白。
第四类——小RNA病毒及披膜病毒。其IUR是+RNA,病毒RNA(VRNA)基因组是有感染性的,可以利用细胞核糖体进行转译。这类病毒如脊髓灰质炎病毒的+RNA不需要先戴上“帽子”,就能在细胞内起mRNA的作用,转译病毒特异的RNA多聚酶,然后反过来转录病毒RNA的互补链(cRNA也即-RNA),因而在原来的VRNA链上形成对RNA酶有抵抗的双链结构,但不含对RNA酶敏感的尾端。这种形式称为复制型(RF型)。稍后,原来的VRNA随着复制的进行逐渐脱落,继而在cRNA上复制新的VRNA,也表现对RNA酶不敏感的双链RNA性质,称为复制的中间型,同时含有对RNA酶敏感的VRNA尾端(或RI型)。由病毒多聚酶以cRNA(-RNA)作为模板反过来转录病毒的mRNA。
小RNA病毒只产生一种新生的mRNA,这种mRNA只转译一种病毒的大分子多肽,在病毒颗粒形成的过程中集结起来,并逐渐裂解产生若干小多肽。披膜病毒产生二种mRNA: 一种是分子量与原来的VRNA相同的mRNA,其功能是转译病毒的酶;另一种是分子量较小的mRNA,功能是转译病毒的大分子肽。但是只有分子量较大的+RNA能组成病毒颗粒中的RNA。
病毒颗粒中的RNA具有多聚A尾端,尾端的长短随病毒而异,在Mengo脑炎病毒,有15个多聚A,而脊髓灰质炎病毒则有90个多聚A。有些披膜病毒如乙型脑炎病毒不含多聚A尾端。
第五类——反录病毒。其IUR是+RNP,内含2个35S的+RNA和反转录酶,以及RNaseH和宿主tRNA。此外亦含有RNA多聚酶活性。二条RNA链在其5′末端相连接。病毒颗粒内的宿主tRNA具有在RNA指导下起转录DNA引物的作用。反录病毒的+RNA的5′端有m7G(5′)ppp(5′)GmpGp的“帽子”。虽然反录病毒RNA是没有感染性的,但从它转录的cDNA则是有感染性的。
反录病毒复制时需要另一种DNA的合成通道,利用病毒的+RNA为模板,借助病毒自身的反转录酶,在胞核内合成一种35S的—DNA的中间产物,然后再用这种—DNA为模板合成10S的+DNA节段,并借连接酶的作用连接成分子量为5.7×106道尔顿的±DNA前病毒。在许可性细胞内,前病毒受细胞特定基因的控制而整合到细胞染色体中,进行复制,同时转录mRNA,包括35S的+mRNA,还有22S和14S的+mRNA。mRNA转移到细胞质中进行蛋白的转译,主要是转译病毒的核壳蛋白、膜蛋白和反转录酶等。
RNA病毒基因组的转录、复制和转译基本上与DNA病毒相似。
病毒的装配与释放 装配包括病毒增殖的成熟阶段,而释放是病毒颗粒最后脱离宿主细胞的阶段。人们早就发现许多病毒在某种条件下可以被拆散,在另一些条件下又可以重新装配成病毒颗粒。Fraenkel-Conrat和Williams在1955年即报道了TMV在试管内重建问题。此后许多学者对病毒在试管内重建问题进行研究,借以搞清病毒在细胞内的装配机理,搞清为什么只有病毒蛋白而不是宿主蛋白能以一定的种类和数目集结,装配成病毒的核壳,同时只有一套病毒核酸,而不是宿主核酸,能够装入这种病毒外壳之中。虽然有些实验说明病毒核酸在试管内也能够和非病毒蛋白结合,但是病毒颗粒的装配说明装配是一种特异的生化过程,因为病毒有一个能为自身蛋白编码的基因。
病毒颗粒的装配随病毒类型的不同而稍有差别。总的来说,可分为无包膜病毒和有包膜病毒两种。
无包膜病毒如小RNA病毒、腺病毒及小DNA病毒,有一个二十面体的外壳,其装配过程是先由基因转译的大分子多肽裂解成若干较小的肽,构成空心外壳前体(这种空心外壳又可称为前病毒颗粒)。此后病毒核酸或核蛋白核心装入空心外壳,同时继续裂解并与新合成的蛋白结合形成完整的病毒颗粒。关于病毒蛋白亚单位是如何集结为空心外壳的机理目前尚不大清楚,但是在对腺病毒及疱疹病毒的研究中看到,受染细胞缺乏精氨酸时,外壳蛋白的转译虽然不受影响,但是不能集结成病毒外壳,因而也无完整的病毒颗粒产生。可能精氨酸参加了某种蛋白的转译,其作用是使外壳蛋白集结,而缺乏精氨酸的细胞不能合成这种蛋白质。
有包膜的RNA病毒如披膜病毒、弹状病毒、副粘病毒及正粘病毒等没有二十面体的核壳而有包膜,由病毒的各个RNA节段与其核蛋白(NP)共同组成螺旋状对称的核壳。这些病毒的核壳都在胞质内形成。
有包膜的DNA病毒如疱疹病毒等,所含的蛋白亚单位的种类及数目很多。大约由4种大分子多肽构成空的二十面体外壳前体,然后塞进新合成的病毒DNA。以后外壳仍继续裂解并继续成熟,形成完整的病毒核壳。
RNA病毒核壳的装配在胞质中进行(肿瘤病毒还不大肯定),而DNA病毒除痘病毒可以在胞质中和胞核内进行外,其他病毒都在胞核内进行,其蛋白质也是在胞质内转译,然后转移到胞核内进行装配。
有包膜的病毒在生成核壳后病毒颗粒还没有成熟,还需要在其表面上再加上一个包膜。RNA病毒套上包膜的过程是芽生。在细胞质膜上有若干经病毒蛋白改变了的糖蛋白(即装配后病毒的包膜蛋白)的集结点,病毒RNA核蛋白和一些其他结构蛋白转移到这些地方时即向外突出,形成芽状,同时套上包膜。病毒颗粒即成熟而释放到细胞外。
有包膜的DNA病毒用芽生方法套上包膜的只有疱疹病毒,是在核膜上获得包膜蛋白的。但是在经过细胞膜时又带上若干蛋白亚单位构成一层外膜。痘病毒不经芽生,是在细胞质内复制和转译各种病毒蛋白亚单位后,并在胞质内装配包膜而成熟的。
无胞膜的病毒和不经芽生成熟的病毒一般都是通过使细胞破裂的方式而释放到细胞外的。
- (二) 畜牧业跨入全省领先行列是什么意思
- (二)畜牧业项目是什么意思
- (二)畜牧厅系统是什么意思
- (二) 畜牧水产是什么意思
- (二)畜禽、水产良种选育是什么意思
- (二)畜禽存栏是什么意思
- (二) 畜禽粪便是什么意思
- (二)畜禽良种的引进与改良是什么意思
- (二) 畜禽饲养量是什么意思
- (二)疏林地是什么意思
- (二)疟疾的防治是什么意思
- (二) 疫病防治是什么意思
- (二)疫病防治是什么意思
- (二)疫病防治体系建设是什么意思
- (二)疾病防治是什么意思
- (二)疾病防治工作是什么意思
- (二)病虫害防治是什么意思
- (二) 百强供销社简介是什么意思
- (二)百货是什么意思
- (二) 皇嗣是什么意思
- (二)益虫是什么意思
- (二)益鸟是什么意思
- (二)盐碱地改良综合技术措施是什么意思
- (二)盐碱害是什么意思
- (二)监管手段和内容是什么意思
- (二) 直属企业单位是什么意思
- (二) 相持阶段到来时国民党的转守为攻方略和积极反共政策是什么意思
- (二)省内转移是什么意思
- (二) 省柴节煤灶是什么意思
- (二)省级农业丰收计划是什么意思
- (二)省级农业管理机构的历史沿革是什么意思
- (二)省级林业技术推广机构是什么意思
- (二)省级畜牧兽医技术推广机构是什么意思
- (二)省营机械农场是什么意思
- (二)着力搞好结构调整是什么意思
- (二)瞄准国内外两大市场,积极发展畜牧业是什么意思
- (二)石河子农学院是什么意思
- (二) 石油是什么意思
- (二)石油是什么意思
- (二)石马牌肉松、油酥肉松、猪肉脯是什么意思
- (二) 研究是什么意思
- (二)研究任务和成果是什么意思
- (二) 研究方向和任务是什么意思
- (二) 确定农副产品的合理定价是什么意思
- (二)确定开发项目是什么意思
- (二)确定资本的占有条件和为资本定价是什么意思
- (二)确定项目计划是什么意思
- (二)碘缺乏病防治是什么意思
- (二)磷肥工业是什么意思
- (二)社会主义初级阶段的基本路线教育是什么意思
- (二)社会主义市场经济体制的逐步建立是什么意思
- (二) 社会主义思想教育是什么意思
- (二)社会主义思想教育是什么意思
- (二)社会主义改造完成至改革开放之前是什么意思
- (二) 社会主义改造时期是什么意思
- (二)社会主义改造时期是什么意思
- (二)社会农业成人教育是什么意思
- (二) 社会化服务体系是什么意思
- (二) 社会化服务体系建设是什么意思
- (二) 社会发展水平是什么意思