生物大分子的能量转移
能量转移就是指能量以同一种形式从一处向另一处的传递。能量的传递可以是宏观的,也可以是微观的。这里能量转移是指微观的能量传递,也就是生物大分子吸收光子能量处于激发态时,能量从最初的吸收部位向其它部位传递的过程。能量转移有时也叫做能量迁移。严格地讲能量迁移是指能量的多次转移。
最早发现能量转移的是Cario与Franck(1922),他们用汞吸收的光照射汞与铊的混合气体,可以观察到两者的发光光谱,因为铊不能吸收这种激发光,所以它的发光必定是间接地从汞原子得到的能量,他们把这种现象叫做敏化荧光。1949年Forster用量子力学方法处理溶液中的能量转移,提出了关于能量转移的一般理论,使它适用于供体和受体间相互作用的所有形式。
能量转移包括多种形式,主要有激发能转移、激发电子的转移 (或迁移)、激子转移、电荷转移与质子转移。Forster提出的激发能转移主要是指电子激发能的非辐射共振转移。
非辐射共振转移 非辐射共振转移是指供体和受体分子间不经过辐射而进行的激发能的转移,一般简称为共振转移。要进行这种转移必须满足三个条件: 第一是能量供体必须能发荧光,在没有受体存在时的荧光量子效率愈高,能量转移过程也愈有效;第二是供体的荧光光谱必须与受体的吸收光谱有重叠,重叠愈多转移效率也愈高;第三是要发生有效的能量转移,供体与受体间的距离不能超过50~100Å。
Forster认为这种能量转移是靠供体和受体二个偶极子之间的电磁耦合作用而实现的。他提出了共振转移和距离关系的著名Forster方程式:
R06=8.8×10-25K2n-4φDτ
式中τ称为重叠积分,和供体的荧光光谱与受体的吸收光谱的重叠积分有关; n是介质的折射系数; K是和一对偶极子之间的夹角有关的取向因子; φD是能量供体的量子效率。他还进一步推导出距离与能量转移效率(E)的关系。设r是供体与受体之间的实际距离,R0是能量转移效率为50%时的距离(另外50%的能量通过其他去活化作用转化),叫做临界距离,
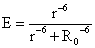
Forster的理论认为这种能量转移是发生在单线态-单线态之间的,转移效率是与r的六次方成反比。Stryer等人则进一步证明,三重态-单线态之间也存在能量转移,这是一种延迟荧光,可以作为单线态-单线态能量转移的补充,转移距离也可达到65A。至于三重态-三重态的能量转移则只能发生在<15A的距离范围内,它的作用机理不是偶极-偶极的相互作用,而是电子交换作用。Stryer与Haugland总结了所有这些结果,提出了一个新名词叫做“光谱尺”,它是指通过光谱测定并利用Forster公式测量生色团之间的距离。这也是目前能量转移实际应用的主要方面。例如免疫球蛋白是由二条轻链和二条重链结合成为“Y”形结构,在分叉的二端点部位标记荧光分子,就可以利用光谱尺的原理测出两端点之间的距离。此外能量转移还可用于研究蛋白质构象特性以及变性或与底物结合时的分子构象的变化。
对蛋白质中能量转移已有很多研究。从能量转移来讲,三种芳香氨基酸的转移次序是: 苯丙氨酸→酪氨酸→色氨酸。在蛋白质内部也是如此,只要蛋白质中有色氨酸,蛋白质表现出来的就是色氨酸的荧光光谱,若蛋白质中没有色氨酸,则表现出酪氨酸的荧光。已经肯定可以在不同分子间进行能量转移,但对相同分子间的能量转移特别是色氨酸→色氨酸之间的能量转移尚有争论。
在多核苷酸中,在天然DNA的同一股上相邻以及下一个相邻的几个碱基之间可以发生共振转移,这是振动弛豫前的单线态能量转移,转移率为1012/S,所以它可与振动弛豫竞争。在多聚腺苷酸(poly A)中可在50~100个碱基中进行三重态的能量转移。在低温下,二核苷酸不论是哪一种碱基吸收光子,都可将其激发成二聚体。相邻碱基间可发生单线态的能量转移,最后转移到二聚体上。在多聚脱氧腺苷酸胸苷酸(polyd AT)中也是从单线态转移能量到二聚体上。在DNA中,二聚体可直接由激发的单线态形成,也可从来自单线激发态的较高振动能级的三重态形成。在室温下DNA的激发能转移,不论是单线态或三重态的转移效率都比低温下小一些,在较高温度时激发态的寿命减少很多。
激子转移 激子转移也是一种激发能的转移,它发生在分子聚集体上。当分子聚集体中一个电子被激发时,它和留下的一个空穴相互联系成为一个激子(exciton),由于分子聚集体本身的性质,使激子可以是非定域的,而能在整个聚集体上转移,这种现象就称为激子转移。由于激子中包括一个带负电的电子和一个带正电的空穴,整个激子呈中性,所以转移的结果不影响聚集体的导电性。
电子转移 激发电子转移简称电子转移,是指借电荷进行的能量转移。这是一种半导体导电的理论,当电子被激发到半导体的导带上时,电子可在导带上运动。由于导带上的电子已不受空穴束缚,在电场作用下电子和空穴将反向运动从而增大导电性。
Szent-Gyorygi在1941年首次提出在生物体系中有电子导电。他认为蛋白质具有半导体性质,所以能量迁移可通过激发电子(及空穴)进行。但许多实验测定证明,蛋白质的禁带宽度太大,不能用可见光把电子提高到导带进行传导,蛋白质的禁带宽度一般在2~3eV之间甚或更高,3eV就相当于70kcal的热量亦即相当于3000Å左右的紫外线光子能量。所以用紫外线激发可以产生半导体导电,而用可见光激发不能导电。但如果蛋白质结构中有缺陷或杂质存在时,激发所需能量可以减少,仍可以进行电子转移。
至于核酸,Gardy等人用顺磁共振谱仪早已证明它具有半导体的性质,但是未作直接的测定。Duchesne(1960)在273~313°K下测定了核酸的禁带宽度 (即能隙△E),如鲱鱼精子DNA的△E=1.80eV; 小牛胸腺DNA的△E=1.78eV。不同来源的DNA禁带宽度都小于2eV,所以认为DNA具有半导体性质。关于蛋白质和核酸半导体性质的研究仍有待深入。
电荷转移络合物 电荷转移络合物在生物大分子中的重要性,是Szent-Gyorgyi在“生物能力学”(1957)一书中强调以后才被引起重视的。电荷转移络合物是指在一络合物内,在两个组成成分间进行单电子转移的络合物。给出电子的叫电子供体,接受电子的叫电子受体。络合物中的二个分子并未分开,所以电子只是部分转移,只有当络合物中的两个分子完全分离时才各具有一个非偶电子,而以离子自由基形式出现。
要进行电荷转移必须具备两个条件: 一是两个分子要靠得很近。有些电荷转移过程可以在活体中进行,而在溶液状态反而不能进行,其中一个重要原因就是两个分子相距太远不能形成络合物; 二是要求受体的电子亲和势大于供体的电离势,供体的离解能愈小愈易给出电子,受体的亲和力愈大愈易接受电子。这样供体基态上的电子就能自发地转移到受体分子的最低激发能级上。
当电荷转移络合物发生电子转移后,会出现新的红移吸收带。例如色氨酸-腺嘌呤的冰冻水溶液,激发后出现的红移吸收带在362nm。在嘌呤与氨基酸的相互作用中,嘌呤既可作为芳香氨基酸的受体,也可作为电子受体,而给出电子的是氨基酸尚未离解的NH2基团。
目前已有愈来愈多的人用电荷转移络合物来解释许多重要的生命过程,例如光合作用、视觉过程和致癌物质的作用等。各种活性不同的致癌物质都能和蛋白质形成电荷转移络合物,同时出现络合物所特有的红移吸收带,并发现致癌能力的强弱和红移吸收带的大小有相互平行的关系,这种关系有助于致癌物作用机理的探索。
质子转移 质子转移最早是Теренин发现的,他发现当溶液中溶质分子吸收能量后,将质子转移到溶剂中,或者是在一个络合物中的两个组分之间,产生氢质子的转移过程。后来对于质子转移的研究,更多的是与水结构联系起来(见“水结构”条)因为大分子周围的水,也象冰一样是有结构的,不同的是它的结构较疏松,距大分子愈远结构性愈小,质子就在水的有序结构中穿梭。当质子受到能量活化时,就从一个水分子中逸出,而当另一个水分子有了多余的质子和能量时,也可以把另一个氢质子推出去,被推出去的自由质子又可重复上述过程。质子转移的结果就是氢键的断裂,结构的重排,同时转移能量。
所谓氢键就是一个质子共享两对孤电子,每对孤电子相当于一个深的势阱,所以两对孤电子就相当于二个势阱,中间形成一突起,叫做位障。在经典力学中,氢质子要从这一势阱跳到另一势阱中,需要活化能才能越过位障。而在量子力学中,质子可看成是一束波,它可从一个势阱“穿过”位障到另一势阱中,毋须具有越过位障的活化能,这种作用就是质子转移的隧道效应。由于质子转移的隧道效应往往使配对碱基异构化,因而在DNA进行下一轮复制时有可能导致“差错”而发生突变。
- 平头式公共汽车是什么意思
- 平头戟是什么意思
- 平头数是什么意思
- 平头整脸是什么意思
- 平头末端是什么意思
- 平头末端标记是什么意思
- 平头枝是什么意思
- 平头桌席是什么意思
- 平头正脸是什么意思
- 平头熯是什么意思
- 平头甲子是什么意思
- 平头百姓是什么意思
- 平头箭是什么意思
- 平头貂毛画笔是什么意思
- 平头软毛大画笔是什么意思
- 平头辇是什么意思
- 平头部丞是什么意思
- 平头錾是什么意思
- 平头钉是什么意思
- 平头钳是什么意思
- 平头锤是什么意思
- 平头长鬃毛画笔是什么意思
- 平头鞋是什么意思
- 平夷是什么意思
- 平夷守捉是什么意思
- 平夷碑记是什么意思
- 平夷郡是什么意思
- 平奄奄是什么意思
- 平奏是什么意思
- 平奖是什么意思
- 平如油盎是什么意思
- 平如生是什么意思
- 平妖传是什么意思
- 平妖贼记是什么意思
- 平妥是什么意思
- 平妥贴贴是什么意思
- 平子是什么意思
- 平子四愁是什么意思
- 平子定情是什么意思
- 平子归田是什么意思
- 平子戏是什么意思
- 平子诗是什么意思
- 平子赋愁是什么意思
- 平子青是什么意思
- 平字五銖是什么意思
- 平字五铢是什么意思
- 平字桌是什么意思
- 平字见奇,常字见险,陈字见新,朴字见色是什么意思
- 平孙是什么意思
- 平孟起义是什么意思
- 平宁是什么意思
- 平安是什么意思
- 平安万应丸是什么意思
- 平安丸是什么意思
- 平安乡是什么意思
- 平安井是什么意思
- 平安京是什么意思
- 平安使,吴笺谩遣,欲寄愁难。是什么意思
- 平安侯漆圆盒是什么意思
- 平安保险公司是什么意思