减数分裂meiosis
染色体数目减半的特殊有丝分裂过程。发生在有性生殖细胞成熟过程中,故亦称成熟分裂。不同生物类群出现减数分裂的时期不同,可分3种类型:❶合子减数分裂(始端减数分裂),发生在受精卵开始卵裂时,形成具有单倍体的生物体,如紫菜;
❷孢子减数分裂(中间减数分裂),发生在孢子体与配子体世代之间,是高等植物特有的形式;
❸配子减数分裂(终端减数分裂),发生在配子形成时,结果产生精子和卵,是一般动物具有的形式。
减数分裂过程复杂,其持续时间比有丝分裂长得多,包括两次连续的细胞分裂,即减数分裂Ⅰ和减数分裂Ⅱ。染色体减数一般发生在Ⅰ期内,但少数类群可发生在Ⅱ期。两期之间,有一短暂的间期,不进行DNA合成,也不发生染色体复制。由于细胞分裂两次,而染色体只复制一次,所以子细胞内的染色体数目减少一半,变成单倍体(见图)。
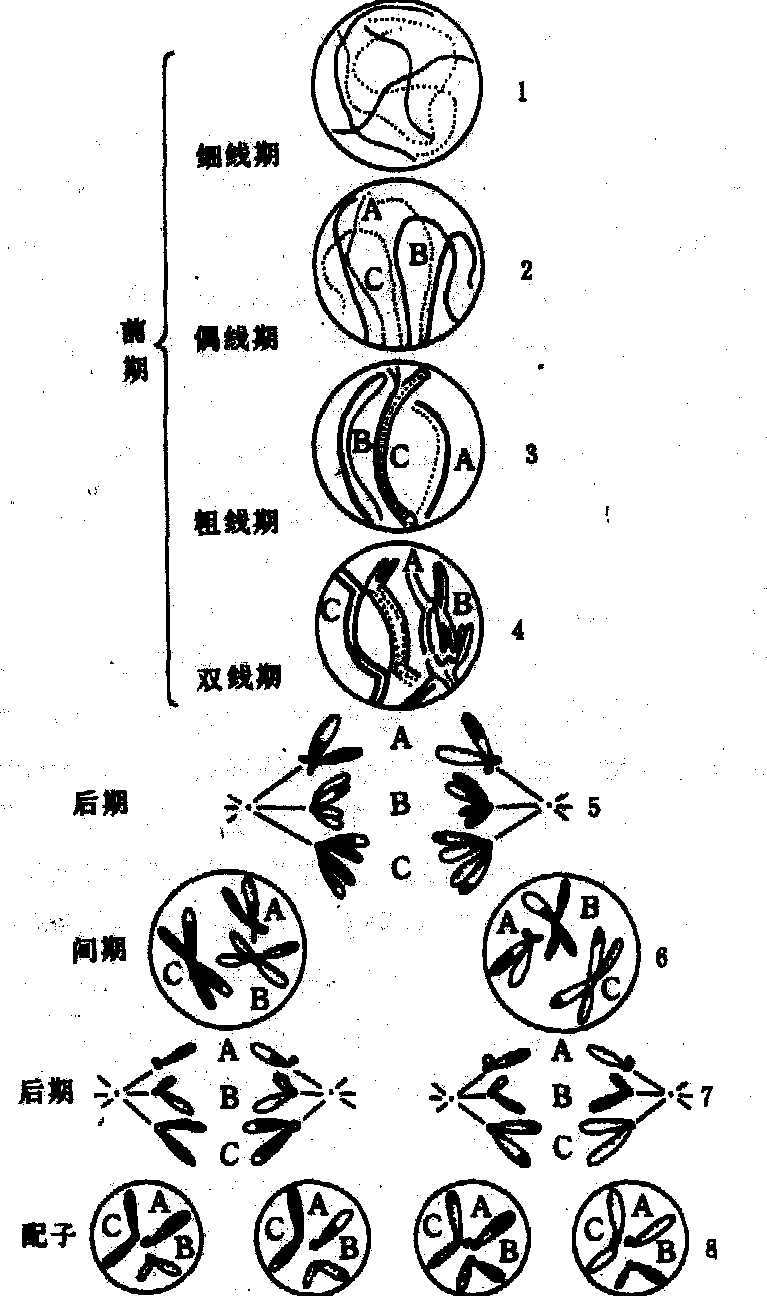
减数分裂示意图
减数分裂前间期 从有丝分裂进入减数分裂的准备阶段,也分为G1期、S期和G2期。此期特点是S期较长,能合成全部染色体DNA的99.7%,其余的0.3%在偶线期内合成(以麝香百合为例)。
减数分裂Ⅰ
前期Ⅰ 减数分裂特有过程一般发生在此期。时间特别长。
❶细线期,核体积增大,核仁也较大,染色体呈细线状,局部可见念珠状的染色粒,可能是染色线紧密围绕而成。每一染色体有两条染色单体构成,因其很细,故看不出结构的双重性。
❷偶线期,亦称合线期,此时发生同源染色体纵向锁合的配对过程,称联会。联会一般从染色体靠近核膜的一端或在其全长的若干位点上开始,象拉链一样沿着整条染色体进行,最后达到完全配对。染色体配对靠联会丝复合体实现,其微细结构在各种动植物大体相同。电镜下,可见联会丝复合体由三股平行的线状结构组成,两侧是平行的电子致密度高的侧生组分,两者之间是电子致密度低、显得明亮的中间区,中央是一纤细的中央组分。联会丝复合体的末端与核膜相连,侧生组分的外侧纤维与染色体的基本纤维紧接。宽度在不同物种间差别很大,但在一个物种中却相对稳定。侧生组分宽约40nm,中间区约100nm,中央组分宽约30nm。侧生组分富含精氨酸的蛋白质、核糖核酸及脱氧核糖核酸的敏感成分; 中央组分主要是非组蛋白,近来发现其中含有肌蛋白,可能有使染色体靠近、保持密切配对的作用。
双线期时,联会丝复合体的两条侧生组分分开,只在一些短的片段上保持相连,这可能是交叉位置的标志。联会丝复合体在早双线期开始消失,其解体过程可从中段或末端、中央组分或侧生组分处开始,以后整体消失。联会丝复合体形成开始于偶线期,完成于粗线期,消失于双线期。它的形成标志着同源染色体联会的结束,它的解体则引起同源染色体的分离。联会是减数分裂中最主要的事件,对其研究已有一定进展。
配对后的染色体称二价体。同源染色体配对结果,细胞中染色体由2N条单价体变成N条二价体。
❸粗线期,联会完成后的染色体明显缩短变粗,由于两条同源染色体结合很紧密,所以不易区分。每一配对的同源染色体含有四条染色单体,称四分体。玉米粗线期的每个二价体,可明显看到着丝点、染色粒、异染色质区和核仁组织者区,因此可作染色体的组型分析。此期内同源染色体的非姐妹染色体间发生局部交换。
❹双线期,联会的两条同源染色体开始分离,但在许多交叉点上仍保持相连。一般认为交叉发生在同源染色体的非姐妹染色单体间,是交换的结果。植物的双线期比细线期、偶线期和粗线期都短得多; 在人和其他许多动物,双线期停留时间非常长,如5个月龄人胚的卵母细胞已达到双线期,一直停留到排卵时止。部分脊椎动物如鱼类、两栖类、爬行类和鸟类的卵母细胞和一些昆虫的精母细胞,双线期的二价染色体解螺旋而成似灯刷的构型,特称灯刷染色体。
❺终变期(浓缩期),染色体进一步螺旋化,变得更粗而短,并向核的四周移动,较均匀地分散于核内,这是观察染色体组型的适宜时期。此时,核仁开始消失,四分体明显并向赤道移动。
中期Ⅰ 核膜和核仁完全消失,二价染色体排列在赤道面上,两条同源染色体通过牵引丝各自与两极相连。
后期Ⅰ 二价体的两条同源染色体随机地分别向两极移动,每极只得到N条染色体,经过减数分裂Ⅰ后,染色体数由2N减为N。
末期Ⅰ 与有丝分裂末期相同,移向两极的染色体逐渐解旋变成细丝状,核膜和核仁重新形成,同时细胞质也分裂。许多植物在减数分裂Ⅰ只发生核分裂,而细胞质分裂要到减数分裂Ⅱ的末期进行,使四个核同时分开。
间期 历时很短,有些生物甚至缺少间期,由末期Ⅰ直接转入前期Ⅱ。
减数分裂Ⅱ 在两个新形成的子细胞中同时进行,其过程与普通有丝分裂基本相同。前期Ⅱ时间较短;中期Ⅱ时,染色体又整齐排列于赤道面上;后期Ⅱ时,两条染色单体分别由纺锤丝牵引移向两级;末期Ⅱ时,移向两极的染色体又逐渐解旋,核膜和核仁重新形成,经过胞质分裂,减数分裂过程结束。
个体进行有性繁殖时,如果只靠有丝分裂形成配子,则其配子仍与体细胞一样具有双份遗传物质。每一代的精卵结合后,新个体细胞内的遗传物质含量必将逐代倍增,以致成为生物进化过程中无用且有害的负担。减数分裂使染色体数目减半就可防止因受精而出现染色体数目加倍,从而使亲代和子代之间的染色体数目保持恒定,为后代的正常发育和性状遗传提供物质基础,保证物种的相对稳定性。减数分裂还通过非同源染色体的重新组合,同源染色体间部分交换,丰富了生物的遗传变异,使配子的遗传基础多样化,为生物的变异及其对外界环境的适应性创造条件。
减数分裂meiosis
又称成熟分裂。导致生殖细胞中染色体数目减半的细胞分裂方式。真核细胞分裂形式之一。为有丝分裂的一种特殊方式。由减数分裂Ⅰ和Ⅱ两次分裂组成。其特殊过程主要在减数分裂Ⅰ的前期,此期可细分为细线期、偶线期、粗线期、双线期和终变期5个不同的时期。两次分裂间有一短暂的间期,在此期间不进行DNA合成,不发生染色体复制。由于细胞核分裂两次,而染色体只复制一次,经分裂后染色体由二倍体变成单倍体。受精后又恢复为二倍体,这样使生物子代保持与亲本相同的二倍体染色体数目。在分裂过程中,非同源染色体重新组合,同源染色体间发生部分交换,使后代遗传基础多样化,对环境条件变化有更大适应性。
减数分裂
染色体数目减半的特殊有丝分裂过程,也称成熟分裂。减数分裂时染色体进行1次复制和连续2次分裂,减色体数目减半。分合子减数分裂、孢子减数分裂和配子减数分裂3种类型。减数分裂可分为细线期、偶线期、粗线期、双线期和浓缩期5个时期。经过2次分裂,可形成4个单倍体细胞。
减数分裂jianshufenlie
有性生殖的生物在生殖细胞形成前的某个时期进行的一种特殊方式的有丝分裂,其二倍体原始生殖细胞内染色体复制一次之后,细胞连续分裂两次,使生殖细胞中染色体数目比原始生殖细胞减少一半。可划分为前减数分裂期、减数分裂Ⅰ和减数分裂Ⅱ三个时期。前减数分裂期是指原始生殖细胞(如精原细胞或卵原细胞)进入减数分裂之前的间期,也分为G1期、S期和G2期。前减数分裂期的S期持续时间长于有丝分裂间期的S期,但只合成全部染色体DNA的99.7%,其余的DNA在偶线期合成。G2期是原始生殖细胞由有丝分裂向减数分裂发展的转变时期,也是原始生殖细胞由非同步性有丝分裂转变为同步性减数分裂的时期。减数分裂1是原始生殖细胞经过同源染色体分离实现染色体数目减半的过程。可划分为前期Ⅰ、中期Ⅰ、后期Ⅰ和末期Ⅰ。前期Ⅰ持续时间长,结构变化复杂,通常又细分为细线期、偶线期、粗线期、双线期和终变期。细线期染色质凝缩成的染色体呈带有若干染色粒的细线状,两条染色单体结合紧密。偶线期同源染色体进行配对,配对是靠两条同源染色体之间沿长轴形成联会复合体实现的,同源染色体的配对使细胞中染色体由2n条单价体变成n条二价体。此外,大约0.3%的DNA在偶线期合成,这部分DNA与同源染色体的精确配对有关。粗线期同源染色体配对完成,并进一步缩短变粗。每个二价体中的两条同源染色体紧密结合,共含有4条染色单体,故亦称四分体。在此期间,四分体中非姊妹染色单体间发生局部交换,并发生DNA修复合成,用于修复由于交换产生的染色单体断口。双线期联合复合体解体,配对的两条同源染色体开始分离,只靠一些交叉点联系在一起,交叉是同源染色体的非姊妹染色单体间发生交换的表现。交叉随着时间推移向末端移动。终变期的染色体高度凝缩,交叉数目减少,只在同源染色体端部保留。此时核膜开始解体,核仁开始消失。进入中期Ⅰ之前有一个过渡性的前中期Ⅰ,其染色体高度凝缩,在染色体丝牵引下向赤道面汇集。中期Ⅰ时染色体端部排列在赤道面上,同源染色体的着丝粒分居赤道面两侧。后期Ⅰ二价体的两个同源染色体分离,分别移向两极,每极的染色体数比母细胞减少一半。有的生物没有末期Ⅰ,由后期Ⅰ直接进入前期Ⅱ或中期Ⅱ。有的生物在末期Ⅰ核膜重新出现,核仁再现,染色体展长但仍保持一定形态,随之进行胞质分裂形成两个子细胞,或者直接进入减数分裂Ⅱ。在减数分裂Ⅰ和减数分裂Ⅱ之间有一个很短的间期,但不进行DNA复制。减数分裂Ⅱ类似有丝分裂。在此过程中,每个染色体的两条染色单体分开,分别移向两极,核膜重新形成,染色体复原成染色质,核仁重现。随之进行细胞质分裂。减数分裂完成后,由一个二倍体原始生殖细胞产生出四个单倍体生殖细胞。减数分裂产生的单倍体性细胞,受精后又产生二倍体合子,从而保持物种的遗传性。同源染色体交换和非同源染色体重组导致配子的多样化,使后代产生变异和具有强大生活力。
减数分裂meiosis
系细胞的一种特殊分裂。主要发生在某些高等动物生殖细胞的形成过程中。雄、雌性的成熟生殖细胞均为各自正常细胞染色体的一半,即单倍体,受精结合后2个细胞的染色体相当于1个细胞的染色体数,即2倍体。人的卵原细胞与精原细胞阶段染色体均为46条(23对),为2倍体,但经过分裂后至成熟的卵子和精子时则染色体均为23条,即为单倍体,此分裂为减数分裂。
减数分裂
细胞分裂方式之一。染色体一次复制,紧接着的两次核分裂,生成的细胞染色体数减半,故名。可分为细线期、偶线期、粗线期、双线期和浓缩期5个时期。细线期:染色体呈细线状,全部以一端或两端与核膜连接,形成所谓的花束期。每条染色体实际上已分裂为两个染色单体。DNA复制发生在细线期前,但染色体的双重性要到粗线期才能看得清楚。偶线期:同源染色体联会,从染色体的某些部分开始而逐渐遍及整个染色体。此过程中出现联会丝复合体。联会结束时,出现相当于染色体数 一半的二价体。粗线期:除着丝粒相联外,染色体分开染色单体显然可见,出现n个由4个染色单体构成的四联体。并在其某些位置上,非姊妹单体间发生交换。双线期:除发生过交换的位置外,每一个四联体中由一个着丝粒所带着的两个染色单体与另一对染色单体相互分开,由于这一染色体运动,交叉点逐渐移向染色体的两端,称之为交叉端化。浓缩期:染色体进一步螺旋化而变为粗短,四联体分散在核膜附近,端化完成,核仁消失。第一次核分裂前核膜消失,四联体分布在赤道板上,两个着丝粒指向两极。两个着丝粒的进一步趋向两极使四联体分作两个二联体,第一次核分裂的结果产生两个初级精母细胞,每一个细胞包含n个二联体,它们为重新出现的核膜所包围。第二次核分裂在简短的间期后开始,核膜再度消失,二联体分布在赤道板上,接着着丝粒分裂,这样就使每 一个二联体的两个染色单体分开,分别进入两个子细胞中。通过两次核分裂和细胞分裂,形成暂时联在一起的4个单倍体细胞(即四分子)。
减数分裂
减数分裂又称成熟分裂,是所有真核细胞生物有性生殖中生殖细胞形成时染色体由二倍减少到单倍的普遍现象,是一种特殊形式的有丝分裂。通过减数分裂使成熟的精 (卵) 子所含的染色体为其母细胞两组染色体的一半,即仅有母细胞每一对染色体中的一条。精卵结合形成合子,即新一代的个体,由精子和卵子所带来的父方和母方的两组染色体重新组合,所以染色体数量与母细胞一样。这就保证了各个世代染色体数量的相对恒定性。因为染色体发生重组,所以也形成了亲代与子代以及子代个体之间的差异。
减数分裂发生在由初级精 (卵) 母细胞形成成熟的精(卵)子的过程中,虽然细胞分裂两次,即第一次减数分裂和第二次减数分裂,但染色体只复制一次,因而染色体由二倍数(2n),减为单倍数(n)。
减数分裂的过程
前减数分裂期 相当于一般体细胞的间期。在前减数分裂期的G1期,每条染色体解旋,舒展松散,形成染色质丝;S期为染色质丝的DNA复制;G2期,有少量RNA和蛋白质合成。在这个期间,精(卵)原细胞变成初级精(卵)母细胞,准备进入减数分裂期。
第一次减数分裂 可以分为前期、中期、后期及末期。与有丝分裂不同的是,前期特别长,变化复杂。
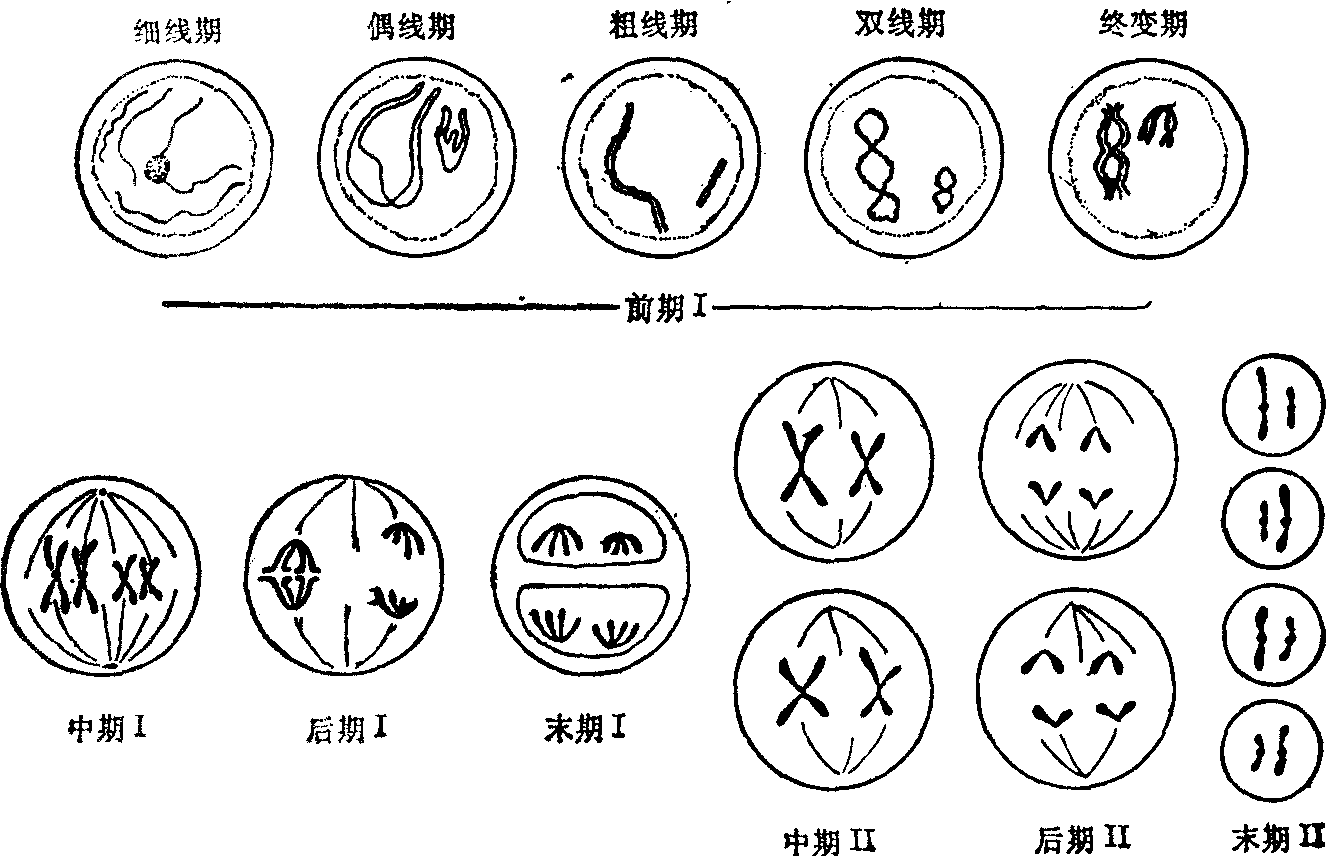
图1 减 数 分 裂
(1) 前期Ⅰ(Ⅰ指第一次减数分裂,以下同):又可分为细线期、偶线期、粗线期、双线期和终变期五个时期。
细线期: 染色体在前减数分裂期很难辨认。在细线期,每条染色质丝虽然含有两个DNA双螺旋,但在表面上只能看到一条舒展松散的细线。在这个时期,有些种类的染色体一端或两端靠近核膜,并集结在一处,形成所谓“花束”。
偶线期:偶线期的主要特点是来自父方的一条和来自母方的一条,在形态上和遗传上都基本相同的同源染色体相互配对。两者从一点或几点开始逐步在全部染色体的相同位点上配对接合,这一接合配对过程叫做联会。人类生殖细胞里的22对常染色体就是这样配对的,只有性染色体X和Y则是末端相接。同源染色体配对完成后,原来细线期细胞中的46条(2n)染色体表面上变成了23条(n),这种由二条同源染色体组成的复合结构称为二价体,具有两个着丝粒。在电子显微镜下,在同源染色体之间可以看到一个纵行的联会复合体,与两侧同源染色体平行。联会复合体两侧为电子密度较高的物质组成,称为侧体,中央有中体(图2)。其构成成分尚未确定。联会复合体的存在可能与染色体的分子配对和交换有关。近来亦有人用镀银染色法,在光学显微镜下,在人的生殖细胞的偶线期看到联会复合体。
粗线期: 本期特点是二价体螺旋化程度不断增加,染色体不断缩短变粗。二价体内每条同源染色体已各自分裂成两条子染色体,借一个着丝粒连在一起,称为姊妹染色单体。这样,每个二价体由四条染色单体构成,叫四联体。在粗线期末,同源染色体的非姊妹染色单体之间发生局部断裂,随后发生部分遗传物质的交换。
双线期: 这个时期的特点是四联体中的同源染色体之间出现纵裂,表明同源染色体开始分开,同时联会复合体也逐渐消失。同源染色体的某些部位彼此连接,这种连接点叫做交叉,是由于交换而形成的细胞学表现。交叉的形式多样。一般认为,有一个交叉就代表一次交换。终变期: 染色体螺旋化程度更高,所以更为粗短,形成各种形状。核仁和核膜消失,纺锤体开始形成。四联体分散于核膜附近。
(2) 中期Ⅰ:纺锤体形成,四联体的着丝粒排列在赤道面上,有两条牵引丝分别与两条同源染色体的着丝粒相连。
(3) 后期Ⅰ:由于牵引丝的作用,四联体中两条同源染色体分开,分别向细胞的两极移动。每一极只能获得每对同源染色体的一条,叫二联体,这就是实际上的减数分裂。至于四联体中的二联体移向哪一极,完全是随机的。(4) 末期Ⅰ: 核膜和核仁重新形成,细胞质分裂,形成两个子细胞。初级精母细胞分裂成两个次级精母细胞,而
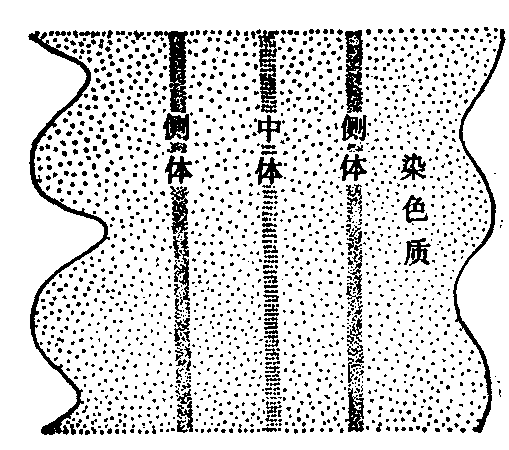
图2 联会复合体
卵母细胞则由于细胞质不等分裂,形成一大一小的子细胞,大者称为次级卵母细胞,小者称为第一极体。第二次减数分裂 第二次分裂之前,是否有间期形成,依种类不同而异。第二次分裂与普通的有丝分裂一样,每个二联体的着丝粒分裂,二条姊妹染色单体分离成二条独立的染色体,分别进入两个子细胞中。初级精母细胞经过两次分裂形成四个精细胞,最后经过转变,变成四条精子。卵母细胞的第二次分裂停留在中期,受精之后,才完成第二次分裂,细胞质又进行一次不等分裂,形成一个成熟的卵子和一个第二极体。减数分裂的结果是,一个精母细胞形成四个精子,一个卵母细胞形成一个成熟的卵子;每个精子和卵子各含单组染色体,即每对同源染色体中只有一条。
减数分裂的性别差异 在人类,男女生殖细胞在减数分裂中,除染色体的变化与行动一致外,还有许多差别。在男性胎儿时期的细精管内,精原细胞即已存在,但一直等到青春期才开始进入精子发生时期。从青春期起数十年的育龄里,不断产生精子。每个初级精母细胞,经过减数分裂和转变,变成四个精子。与此相反,在早期女性胎儿卵巢里,卵原细胞已分化成初级卵母细胞,约在第四个半月到第五个半月期间,胎儿卵巢里的卵原细胞和初级卵母细胞增殖到最大限度,约有700万个,以后逐渐退化,到出生时,只剩下200万个,其中大约有400多个在生育年龄里被排出体外。初级卵母细胞在胎儿时期即已进入第一次减数分裂,在双线期末,初级卵母细胞的染色体重新解旋,变化成松散的核网状态,叫做核网期,此时,初级卵母细胞中止分裂,并合成大量胞质,细胞迅速长大,直到青春期。从青春期起,一般是每月排出一个卵母细胞。在排卵之前,才完成第一次减数分裂,形成次级卵母细胞和较小的第一极体。绝大部分的初级卵母细胞在整个生育年龄都保持核网状态。排出卵巢的次级卵母细胞,在输卵管内进行第二次分裂,到中期停止,此时如果受精,即可完成第二次减数分裂,形成一个成熟的卵子,排出第二极体。卵子受精时卵核与精核结合,形成合子的胞核,开始新一代的个体发育。
减数分裂中染色体行动规律 在正常减数分裂中,染色体的行动规律是同对的两条染色体分开,异对的则自由组合。如人体有23对染色体,精卵结合后的组合类型有223种。Carothers (1921)发现某些蝗虫的减数分裂时正常配对的染色体,在大小和形状上有能辨别出来的差异。她观察了同一个体里染色体的行动,据染色体上可辨认的特点,从细胞学上证明了,同对的两条染色体彼此分开,异对染色体之间则自由组合(图3),因为基因位于染色体上,所以从细胞学上证实了孟德尔式分离与自由组合定律。由于减数分裂和受精中染色体的行动和重组,于是保证了亲代与子代染色体数量上的相对恒定性,同时也造成了子代个体之间的差异,因而使同种生物的各个世代之间以及个体之间的形态特点基本相同而又不尽相同。
减数分裂中染色体的特殊现象是联会与交换。联会是交换的条件,但联会的机理至今尚未明确。交换的结果,表现为交叉。
同源染色体之间发生部分交换,从Stern (1931)的果蝇杂交实验中可得到证实。实验用雌果蝇的两条X染色体形状不同,很容易识别:一为L形,由Y染色体易位到X染色体上而形成的; 另一条X染色体断裂成两个片段。把这种雌果蝇同雄果蝇交配,子代果蝇除亲代雄果蝇X或Y染色体外,从母蝇传来的一条X染色体共有四种不同的形状: 两种与母蝇两条X染色体相同,即一种为L形X,另一种为由二个片段所成的X,反映同对的同源染色体彼此分离,这两种占子代的多数。另有两种X染色体:一种X染色体表现正常形状,另一种X既有一个较短的L形片段,又有一个小的片段,这两种X占子代的少数,说明母体里有少数卵母细胞的两条X染色体
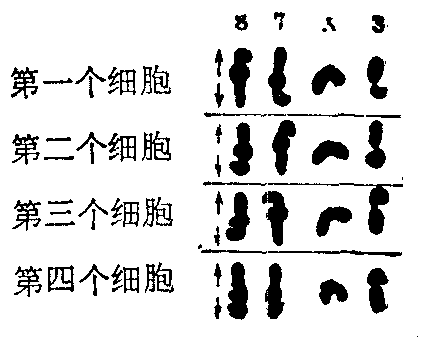
图3 昆虫减数分裂中期末异对染色体之间的自由组合方式(仿Carothers)
图示四个细胞中接近后期的染色体3号、7号、8号和X染色体的排列情况,每号染色体的两条同源染色体用上下两段代表,形式彼此不同,X染色体只有一条。根据四对染色体在赤道板上的排列,可以肯定四对染色体之间是自由组合的,箭头示后期同源染色体分离的方向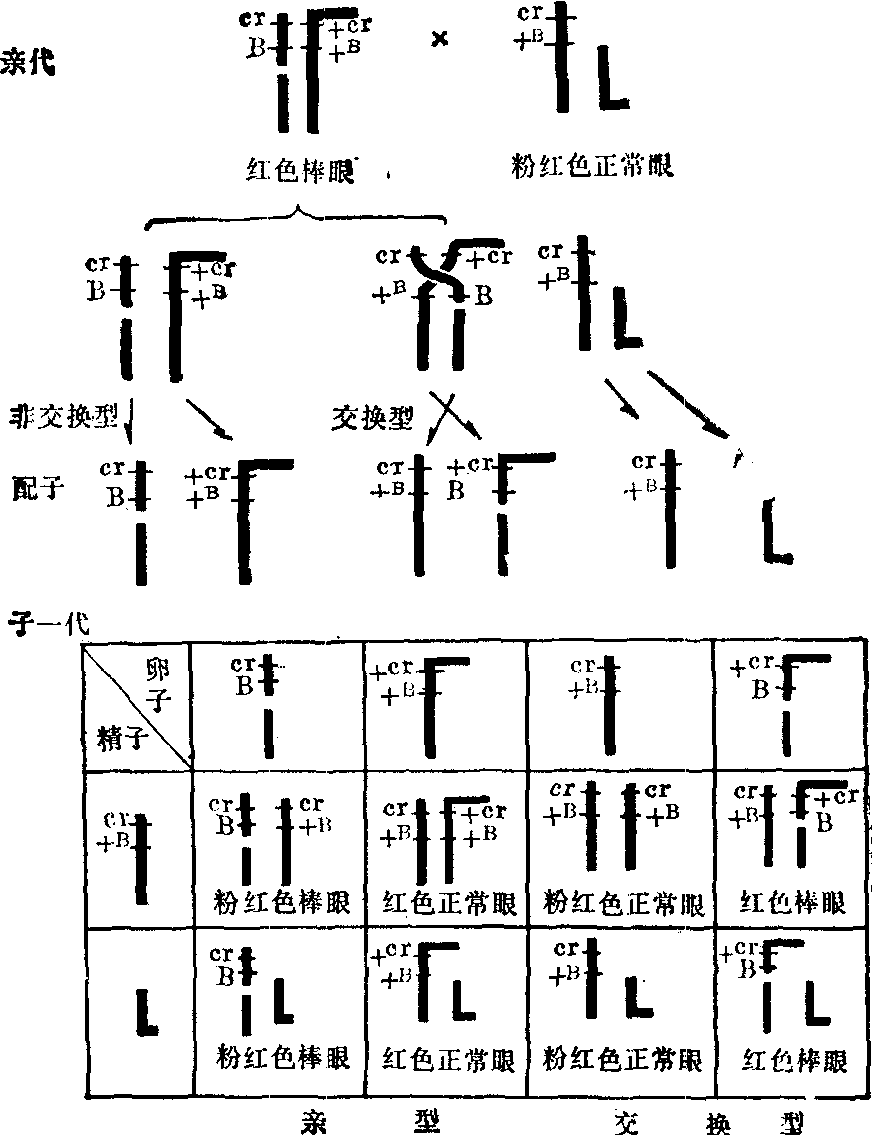
图4 果蝇杂交实验(自Stern)

在减数分裂过程中,如果染色体行动不规律,例如某对染色体不分开,就会造成配子中多一条或少一条染色体,在人类就将形成染色体病。
减数分裂
有性生殖是生物长期进化历史中较无性生殖更为进步的一种繁殖方式。雌雄配子的融合把不同遗传背景父母双方的遗传物质混在一起,其结果既稳定了遗传性,又添加了诸多新变异,这便大大增强了生物对千变万化的环境的适应能力。然而,如果没有一种机制使精卵细胞染色体数减少一半,则精卵融合将使染色体数倍增下去。事实上,染色体数为2n的体细胞通过一种特殊的有丝分裂,即减数分裂方式产生出染色体数为n的精卵,精卵融后又恢复2n。减数分裂是DNA复制一次继而细胞分裂两次,结果染色体数减少一半。
减数分裂前间期 生殖细胞如精原细胞在进行减数分裂之前首先要经过一次或几次不同一般的细胞间期,称之为减数分裂前间期。什么原因决定这种有丝分裂向减数分裂的转变尚不十分清楚,但这种转变和减数分裂前的间期活动有密切关系。减数分裂前间期也分为G1、S、G2期,但不同的是S期特别长,如蝾螈的S期由原来的12h增长到10天,此种现象在其他动、植物种类中也很普遍。此种延长并非由于复制叉的运动减慢,而是由于每单位长度DNA复制单位的启动数量减少所致。有的种类如小麦减数分裂之前的3次间期中S期延长,由3.8h延长到12—15h,这种反应可能是由于对细胞外因子如激素在细胞内浓度增加所致。此外还有一些精细的变化,如许多动物的性染色体的DNA复制总是在S期之末,而在减数分裂前间期的S期中变为早S期复制。另一令人瞩目的现象是在百合中,减数分裂前S期只合成全部染色体DNA的99.7%,其余的0.3%在偶线期合成。有些细胞阻断在减数分裂前的G2期,直到短时间冷休克处理,可启动减数分裂的发展。
减数分裂过程 由减数分裂前G2期细胞进入二次有序的减数分裂。第一次减数分裂可分为前期Ⅰ、中期Ⅰ、后期Ⅰ、末期Ⅰ。第二次减数分裂可分为前期Ⅱ、中期Ⅱ、后期Ⅱ、末期Ⅱ。两次分裂之间的分裂间期或长或短,无DNA复制。
第一次减数分裂有以下四期。
前期Ⅰ 前期Ⅰ变化最为复杂,呈现许多减数分裂的特征形态变化,时间长,可达几周、几月,甚至几年、几十年。前期Ⅰ又可根据染色质的变化分为以下各期。
(1) 细线期:或称凝集期。细线期染色体虽已复制,但染色体仍呈单条细线,其两端通过接触斑和核被膜相连,有的物种染色线集中在核的一端呈花束状向外放散,故又称花束期。染色线上有成串的膨大处,类似念珠,称之为染色粒。此期核体积增大,核仁也较大。
(2) 偶线期: 又称配对期。此期在光镜下有时可见染色体已经成双。一对同源染色体一条来自父方,一条来自母方。在此之前,一对同源染色体在核中随机分布,有时有相当的距离。到偶线期时,同源染色体与核被膜相连的端部移位到一起,两条同源染色体侧面紧密相贴进行配对,此现象称之为联会。同源染色体之间的识别,以及使两条染色体移位到一起的原因尚不十分清楚。此种配对是专一性的,一旦两条染色体相接触,便会像拉链一样迅速扩展,直到两条染色体的侧面全部锁合。联会也可同时发生在分散的几个点上,拉链锁合向端部发展。这时期有残余的0.3%的DNA合成,称之为Z-DNA,有人认为这种Z-DNA合成可能和配对机制有关。联会部位形成一种特殊结构,沿同源染色体纵轴分布,宽0.15—0.2μm,光镜可显示这一空间,称之为联会丝复合体(SC),在电镜下可显示其复杂的结构。这一结构参与了配对、重组机制。在细线期或偶线期加入DNA合成抑制剂抑制Z-DNA的合成,则同源染色体不能配对,SC的组装受到抑制。由于一条染色体是由2条染色单体所组成,故每一配对的结构中共有4条紧密结合在一起的染色单体,称之为四分体。由染色体水平来考虑称之为二价体,因每对是由2条同源染色体组成。
(3) 粗线期: 又称重组期。这一阶段开始于同源染色体配对完成之后,染色体明显变粗变短,结合紧密,同源染色体之间发生DNA的片段交换,产生新的等位基因的组合。此时期在SC的梯状结构中间出现重组小节,为球形、椭圆形或棒状,直径约90nm,内含蛋白质,结构尚不清楚。它是一种含有多种酶的“重组机器”,它横跨100nm的SC宽度将父母的单体DNA局部区域结合在一起,通过它发生活跃的重组过程。在此时期可检测出有DNA和组蛋白的合成。DNA合成发生在许多染色体上,有的种类是在编码rDNA的染色体上,称之为rDNA的扩增。合成过剩的rDNA拷贝形成附加的核仁,以后在前期Ⅰ中进行活跃的rRNA的合成。这时期合成一些减数分裂特异的组蛋白,可能在重组机制中起作用。DNA的合成主要用于DNA链的修补、连接,称之为P-DNA。前两阶段时间短,通常只有几小时;相反,重组阶段可以几天、几月,甚至更久。到此阶段同源染色体紧密配对,一直保持与核被膜的接触。粗线期核仁大,含有大量RNA。
(4) 双线期: 又称合成期。重组阶段一结束同源染色体便分开,但有几点相连,第一次看到四分体。这时SC消失,染色体与核被膜脱离接触。同源染色体之间的接触点称之为交叉,这是从形态学角度提出粗线期阶段同源染色体之间发生了交换的证据。此种交叉的数量和部位在同一种类的不同细胞中也有所不同,每一四分体上至少有一个交叉。
在合成期RNA合成活跃,贮存于卵中,有些种类RNA合成限于此时期。爬行类、鸟类、两栖类卵母细胞形成灯刷染色体,上有许多伸出的襻,mRNA和rRNA合成活跃,核仁明显变大、增多,可达上千个。两栖类卵此期长可达一年。人类胚胎卵巢中之卵母细胞5个月可达这一阶段,诞生后经儿童期、性成熟期,直到排卵之前,此期可维持50年之久。通常每月有一次卵成熟可打破此种阻抑进入减数分裂的其余阶段。许多昆虫此时染色体产生去凝集,很像间期核。
(5) 终变期:或称再凝集期。合成期结束,染色体变成紧密凝集状态,这一过渡标志着减数分裂前期的最后阶段出现。大多数种类核仁消失,四分体较均匀地分布在核中,随着交叉向染色体臂的端部移行,称之为端化。最后四分体只靠端部交叉使其结合在一起,姐妹染色单体由着丝粒连接在一起。终变期的完成标志着减数分裂前Ⅰ的结束,其结果发生了染色体的重组,合成了为配子所需要的,或胚胎早期发育所需要的全部或大部RNA、蛋白质及糖类,染色体凝集成短棒状。
当前期即将结束时,像有丝分裂一样,中心体(如有的话)已经加倍,向两极迁移,形成纺锤体。雄性细胞的纺锤体与有丝分裂的十分类似。卵母细胞中心粒在发育过程中消失,纺锤体的形成无中心粒参与,纺锤体形态与植物细胞的类似。
中期Ⅰ 核被膜的破裂为前期Ⅰ向中期Ⅰ转化的标志,纺锤体侵入核区,分散于核中的四分体开始向纺锤体中部移动,不同于有丝分裂的是,四分体上有4个着丝点,一侧纺锤体只和同侧的2个着丝点相连(图1)。此前可称之为早中期Ⅰ; 最后染色体排列在赤道面上,称中期Ⅰ。
后期Ⅰ 同源染色体的分离和向极运动的开始,标志着后期的起点,此时纺锤丝分别拉一对同源染色体向本极移动。后期完成时,染色体趋向两极,每极接受单倍体数量的染色体。由于每条染色体仍含有2条染色单体,因而每极仍是2×DNA含量。不同的同源染色体对分向两极时相互间是独立的,因而父方、母方来源的染色体随机组合,有利于减数分裂产物的基因组变异。如人有23对染色体,父母双方染色体有223的组合方式,由于还有父母染色单体的重组混合,因而除了同卵孪生之外几乎不可能得到遗传上相同的后代。
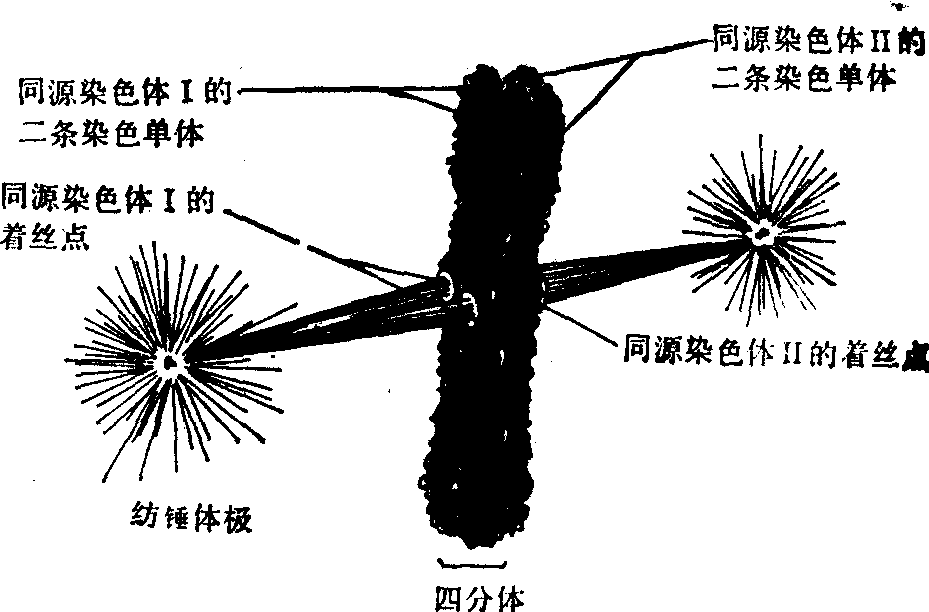
图1 减数分裂中期纺锤体与着丝点相连的图解
末期Ⅰ及间期 在自然界中,末期Ⅰ和间期的类型有二:一种是没有明显可见的染色体去凝集,另一类型是完全逆转到间期核的状态。后者染色体去凝集,核被膜形成围绕两极的染色质。然而大多数种类,末期Ⅰ和间期是在第一次及第二次减数分裂期之间的短暂停顿,在所知的生物中未见有DNA合成。
第二次减数分裂和有丝分裂过程基本相同,可分为前、中、后、末期。如在末期Ⅰ及间期中染色质已经去凝集,则前期Ⅱ染色质重新凝集。中期Ⅱ染色体形成致密的短棒状,排列在赤道板上,每条染色体上的着丝点分别和两个不同极的纺锤丝相连。后期两组染色体拉向两极,其结果每极为1×DNA含量,相当于进入减数分裂的原G2期细胞DNA含量的1/4。末期Ⅱ重新组成核被膜,染色质去凝集,形成四个单倍体细胞(图2)。
所形成的四个核在不同动、植物种类其最终的命运也有所不同。雄性动物形成的四个核为胞质所包围,进一步分化成有功能的雄性配子——精子。雌性动物减数分裂所形成的四个配子中,由于卵的胞质不均匀分裂,胞质集中于一个大的卵细胞中,发育成有功能的配子,在卵的一极形成胞质极少的小细胞,无功能,称之为极体(图3)。高等植物发育类型与动物相似,花药中减数分裂产生的四个产物,形成花粉的“精子”核。在雌蕊子房中经减数分裂,仅有一个发育成卵子核。
大多数动物和少数植物雌性细胞性染色体为XX,雄性细胞为XY或XO。X、Y染色体配对时有部分结合和重组;后期Ⅰ时X、Y染色体分别分向两极;后期Ⅱ时X、Y染色体分别分到四个子细胞中去。XO型中,后期Ⅰ时X分到一个子细胞中,另一细胞为O,后期Ⅱ时四个子细胞性染色体的分配为X、X、O、O。
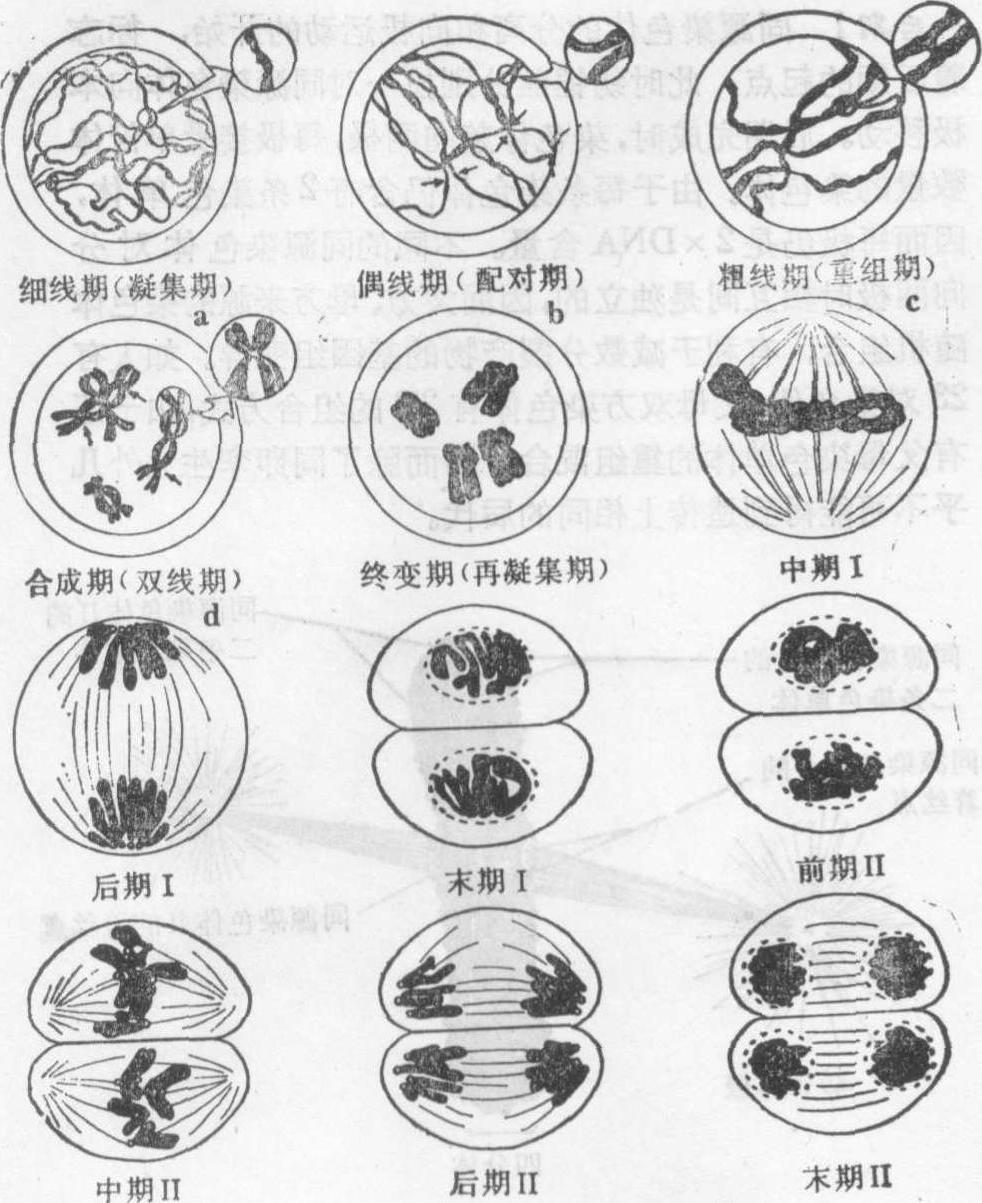
图2 减数分裂图解
a、b、c阶段两单体间裂隙实际看不到,引出的圆圈内结构为局部放大;b箭头所指处配对在发展;d可见四分体
减数分裂的时间和部位 真核生物的生活周期中,减数分裂发生的时间和部位有三种主要类型:
❶配子的或终末的减数分裂: 减数分裂发生在将形成配子之前。一般的动物,还有许多原生动物,少数低等植物属此类型。
❷孢子的或中间的减数分裂: 此类型发生于高等植物和某些低等植物和菌类中,每个世代交替于单倍体及二倍体之间。受精产生二倍体孢子世代,减数分裂发生于孢子世代的某些点上,产生的是孢子而不是配子,通过有丝分裂出芽生长,形成单倍体的配子体世代。成熟时,同一配子世代通过细胞有丝分裂,分化形成卵子和精子,精卵融合又回复到二倍体世代。
❸合子的或起始的减数分裂:受精之后立即发生减数分裂,生成精、卵(未分化者又称为正或负配子),融合后产生二倍体合子,合子立即进入减数分裂,产生四个单倍体细胞,经一次或几次有丝分裂,产生单倍体世代,由此产生配子(图4)。
同源染色体的联会 同源染色体的配对是减数分裂的核心问题。由于有了联会配对,才可能有同源染色体之间的遗传物质的重组和同源染色体的准确分开。而同源染色体之间是如何识别,紧密接触,交换染色质片段等许多问题并未完全阐明。
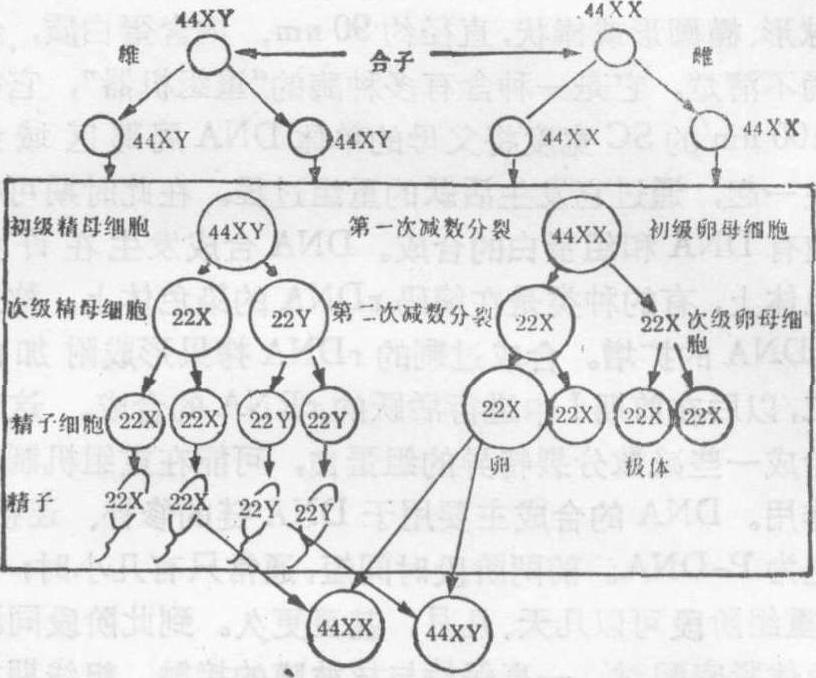
图3 人类精子和卵子发生图解
(a)配子或 (b)孢子的或 (c)合子的
终末 中间的 或起始的
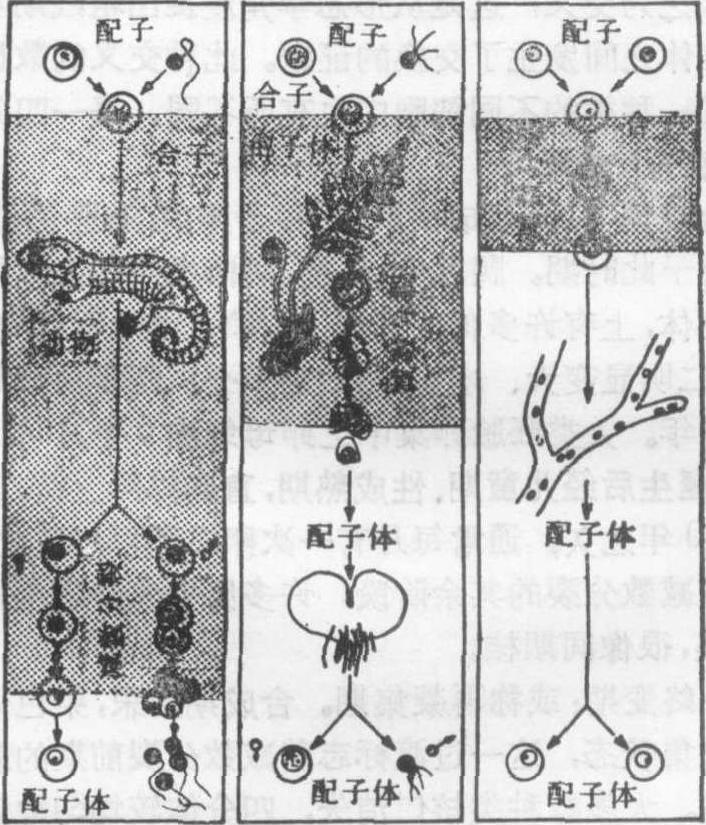
图4 真核细胞中减数分裂发生的时间部位的三种类型阴影处为生活周期中的二倍体时间
SC的结构 SC是应用电镜观察蝲蛄精母细胞二价染色体时发现的,以后在动物的卵母细胞和植物的减数分裂中都得到证明。SC位于两条同源染色体之间,沿纵轴方向延伸。两侧为侧生组分,宽为20—40nm,电子密度高。两侧生组分间为中间区,宽约100nm,中间区在电镜下比较明亮。中间区的中央为中央组分,比较暗,宽约30nm。侧生组分之外为染色质,不同生物或者浓密或者疏松。侧生组分和中央组分之间有横向排列的纤维,大致成直角相连,称之为L-C纤维,长约60—70nm,L-C纤维之间距离20—30 nm。而使SC结构像一架梯子,呈扁平带状。SC主要由碱性蛋白和RNA组成,含有微量的DNA(图5)。

图5 蝗虫粗线期的SC框内为SC与染色质纤维之间的关系图解
SC的形成与去组装 当细线期同源染色体尚未配对时,通过横切面的细致分析,可见有一条侧生组分的轴心位于2条姐妹染色单体之间的缝隙中,而且和染色质密切联系。此时若用Bernhard EDTA染色,侧生组分呈负结果,并可一直持续到配对时,说明侧生组分在充分配对和SC完全形成之前没有RNA与之结合,因而也可以说RNA分子在同源染色体联会中不参与识别过程。减数分裂前的G2期或早前期Ⅰ,对减数分裂信号刺激作出反应,SC的单体在胞质中合成,通过核被膜进入核内,在核内达到高浓度,受到和染色体结合的一些因子的作用,促进亚单位组成侧生组分轴心。两条侧生组分轴心逐渐靠拢,直到距离达100 nm左右时,二者大量横向垂直伸出的纤维,互相交错形成中央组分。继而单链Z-DNA序列或RNA、蛋白质序列识别分子组合到SC之中,进一步促进同源染色体的紧密整列,这时SC便可提供有组织的网络,稳定联会,提供分子配对和与重组活动有关的因子及酶。有人认为侧生组分轴心蛋白亚单位是根据染色质上每一基因位点的严格指导,组成一种结构构型,与另一条相应装配起来的构型相似的轴心可相互识别而结合。联会多由核被膜一端开始,像拉链一样使两条染色体联会到一起。联会后在粗线期进行DNA重组,同源染色体之间交换染色质断片,故在粗线期、双线期有交叉出现。到双线期时同源染色体之间分开,因核条件的改变,推动平衡向单体库方向移动,SC去组装,大多数有机体亚单位分散到核质及胞质中去。有少数种类亚单位局部重组为多聚复合体,以此形式暂时贮存,直到崩解(图6)。
重组发生的机制 细胞经过染色体重组,造成一些染色体上基因序列改变,是由于同源染色体之间染色体片断交换的结果。两同源染色体均向SC伸出襻环,经过L-C纤维直达中央组分,直到距离约1nm时分子可以互相识别。襻环处于连续运动之中,在中央组分中进行碱基序列的连续比较,一直到同源序列排在一起,这时在分子水平上发生DNA链的断裂、交换,最后达到DNA的重组。配对阶段所需时间,反应了包含在单体中的纤维进行序列比较所需的时间。这一过程触发了一系列的酶促活动,如DNA内切酶可将DNA链切断,DNA外切酶可将暴露在外的单链DNA切掉,DNA聚合酶可以填补间隙,解旋酶可使切断的DNA链展开,连接酶可连接DNA链的切口等。大量实验证明,百合中β型核酸酶在偶线期上升,粗线期达高峰,粗线期末消失。3H-TdR标记实验也可见在粗线期有掺入。偶线期末到粗线期连接酶活性上升。粗线期有P-DNA合成,不同于Z-DNA,是由非常短的断片组成,如抑制此种DNA的合成则染色体断裂,故P-DNA在断口修复中起作用(图7)。
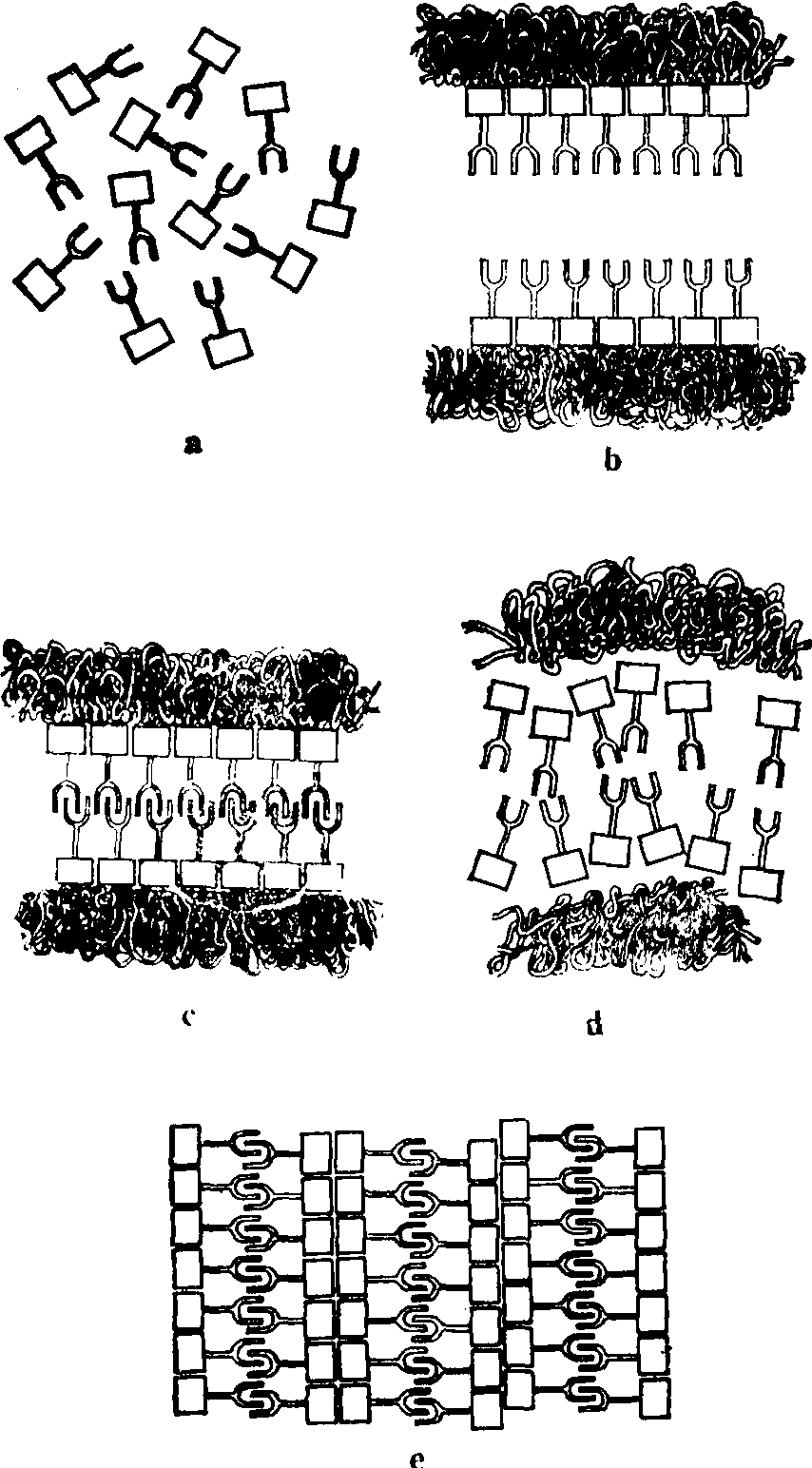
图6 SC和多聚复合体形成的可能顺序
a. SC单体的胞质内合成;b. 单体组装成同源染色体的侧生组分,尚未配对; c.配对形成:d. 粗线期结束时SC解离为单体; e.解离出的单体暂时组装成多聚复合体
由此可见,减数分裂不仅是使有性生殖的生物染色体数目保持稳定的机制,而且是使生物遗传基础发生变异的机制。在减数分裂过程中同源染色体配对,不同对同源染色体自由组合,非姐妹染色单体片段的交换重组,形成众多的不同染色体组成的配子,增加了变异性,扩大了后代的变异范围,增强对外界环境的适应性。
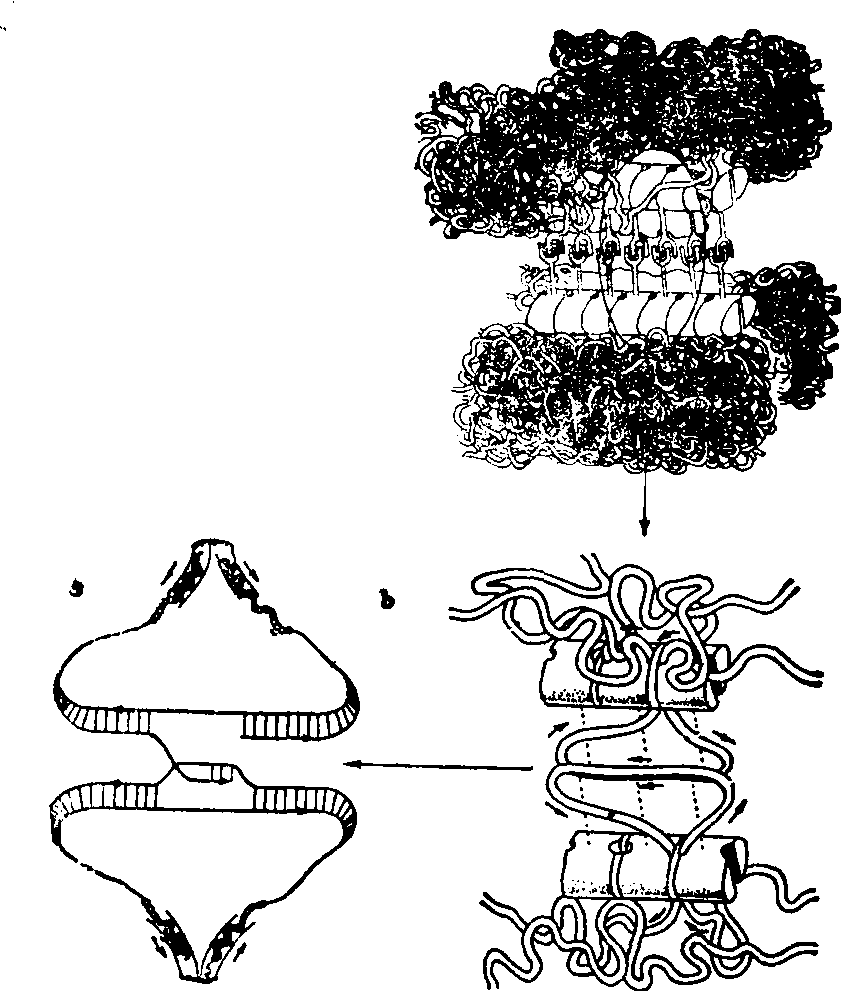
图7 在重组中SC的可能作用
a. 同源染色体初始的配对,SC已经组装; b. 初始配对后同源染色体伸出襻一直延伸到中央组分,襻在连续运动,两侧襻挨近到分子距离; c.在中央组分中发生DNA顺序的连续比较,一旦达到同源顺序的识别,则启动了发生重组的系列活动
减数分裂
meiosis
减数分裂
meiosis
减数分裂
meiosis
- 阿伯的是什么意思
- 阿伯西尼亚是什么意思
- 阿估茉是什么意思
- 阿伽是什么意思
- 阿伽门农是什么意思
- 阿伽门农墓是什么意思
- 阿伽陁丸是什么意思
- 阿佐和阿佑是什么意思
- 阿佐图斯是什么意思
- 阿体八升山是什么意思
- 阿何服内他适案是什么意思
- 阿佛佛是什么意思
- 阿佛加德罗是什么意思
- 阿佛加德罗定律的计算是什么意思
- 阿佛加德罗常数是什么意思
- 阿佛加德罗常数的测定是什么意思
- 阿佛洛狄特是什么意思
- 阿佛罗狄忒是什么意思
- 阿你真那是什么意思
- 阿佤是什么意思
- 阿佤人民唱新歌是什么意思
- 阿佤山是什么意思
- 阿佩是什么意思
- 阿佩氏综合征是什么意思
- 阿佩莱斯是什么意思
- 阿佼仔是什么意思
- 阿使是什么意思
- 阿使仔是什么意思
- 阿侄是什么意思
- 阿侄男女是什么意思
- 阿侖是什么意思
- 阿依丁库勒硝尔鲁克碱滩是什么意思
- 阿依古烈克是什么意思
- 阿依吉伦和伦吉善是什么意思
- 阿依吐尔沟是什么意思
- 阿依吐尔能乔阔峰是什么意思
- 阿依吐尔逊是什么意思
- 阿依吐拉是什么意思
- 阿依尔萨依沟是什么意思
- 阿依尕依库木山是什么意思
- 阿依尕尔特沟是什么意思
- 阿依巴依别勒山口是什么意思
- 阿依库乃斯达坂是什么意思
- 阿依库勒是什么意思
- 阿依拉尼什乔阔峰是什么意思
- 阿依木尼莎是什么意思
- 阿依格尔加勒戈壁是什么意思
- 阿依格尔加勒登喀巴克戈壁是什么意思
- 阿依浪苏山是什么意思
- 阿依特库勒乔阔峰是什么意思
- 阿依苏鲁女皇是什么意思
- 阿依莫孜是什么意思
- 阿依莫玉孜是什么意思
- 阿侬是什么意思
- 阿侬头是什么意思
- 阿侬袋是什么意思
- 阿侬语是什么意思
- 阿侯是什么意思
- 阿俄是什么意思
- 阿俄魁是什么意思