连锁与交换linkage and crossing-over
位于同一染色体上的基因连系在一起遗传与非姊妹染色单体上的基因互换位置,出现重组型的现象。
1906年贝特森(W.Bateson)等用香豌豆(La-thyrus odoratus)做杂交试验,首次发现连锁遗传现象。紫花、长花粉粒(PPLL)和红花、圆花粉粒(PPll)两个香豌豆品种杂交,F2的分离比不是预期的9:3:3:1,而是亲本型比值高于期望值,重组型比值低于期望值;另一组试验是紫花、圆花粉粒(PPll)与红花、长花粉粒(ppLL)的品种杂交,也是亲本型比值高于期望值,重组型比值低于期望值。但是,第一组的杂交是两个显性基因位于一条染色体上,两个与之等位的隐性基因位于同源染色体上,F1基因型为++/ab,称为相引构型。第二组的杂交则是一对同源染色体上都分别载有不同的显性和隐性基因,F1的基因型是a+/+b,称为相斥构型。1911年美国遗传学家摩尔根(T.H.Morgan)等根据果蝇的杂交试验结果提出连锁与交换的概念,指出控制不同性状的非等位基因位于一条染色体上,减数分裂时一起进入配子,表现连锁遗传现象;非姊妹染色单体交换节段,改变基因原有的连锁关系,出现新的重组型。从而发现经典遗传学第三定律——连锁与交换定律。
1909年比利时细胞学家詹森斯(F.A.Janssens)根据螈蝾孢母细胞减数分裂的研究,提出非姊妹染色单体的交叉是交换的产物的交叉说。1931年斯特恩(C.Stern) 等提出基因连锁与交换的细胞学证据。
连锁 如果位于同一染色体上的基因一起进入配子,非姊妹染色单体不发生交换,测交后代只出现亲本型、没有重组型的现象称为完全连锁。完全连锁是比较少见的,由特定的基因型和基因所决定,例如雄果蝇、雌蚕皆不发生交换。1922年英国学者霍尔丹(J.B.S. Haldane)曾提出,异配性别的生物体一般不发生交换。有的突变基因能抑制基因交换,例如果蝇的隐性交换缺陷型C(3)G。这类缺陷型在高等生物如人类、果蝇、小麦和玉米; 低等生物如粗糙脉孢霉、曲霉、酵母菌以及λ噬菌体等都曾发现。如果非姊妹染色单体发生交换,F2及测交子代既有亲本型也有重组型,但重组型的比率低于独立遗传中预期值的现象称为不完全连锁。生物界中不完全连锁是极为普遍的。位于同一染色体上的各个基因相互间都存在着连锁关系,组成一个连锁群。每个物种的连锁群数和单倍的染色体数相等。例如水稻、小麦、陆地棉、人、黄牛、猪和鸡的连锁群数分别为12、21、26、23、30、19和39。位于性染色体上的基因所控制的性状还表现与性别连锁(见性连锁遗传)。可以利用各种试验设计,确定基因所在的染色体及其连锁位置(见基因定位)。把连锁群上的各个基因间的顺序和距离标志出来就绘成了连锁遗传图。至今已对果蝇、玉米、番茄、大麦、小麦、水稻、高粱、豌豆、大肠杆菌和T4噬菌体等绘制出较好的连锁遗传图。高等动、植物连锁图上各基因之间以交换值作为遗传距离,交换值应小于50%。
交换 染色体交换的结果产生了基因重组,交换数和重组数并不是1对1的关系,在两个连锁基因之间发生双线双交换不产生基因重组。重组型配子占总配子数的百分率称交换值,最大交换值为50%。因为单交换只涉及两条染色单体,另两条不发生交换,重组率为50%。双交换,若二基因间发生二次交换则有四种情况(图1): ❶双线双交换,产生的配子全属亲本型,交换值为0;
❷和
❸三线双交换,产生亲本型与重组型配子各一半,交换值为50%;
❹四线双交换,产生的配子全部为重组型配子,交换值为100%。如果交换所涉及的染色单体是独立的,各种交换频率相等,平均交换值等于50%。所有的孢母细胞并不都在所研究的连锁基因间发生交换,交换值一般小于50%。特定连锁基因间的交换率是相当稳定的,因此交换值可用于基因定位(见基因定位)。但交换率也受基因和环境的影响。前者如染色体畸变会降低交换率,再如黑麦自交系的交换率一般低于异花授粉的亲本; 后者如年龄、温度、营养、水分、物理射线和化学药品等也影响交换,雌果蝇在22℃时交换率最低。
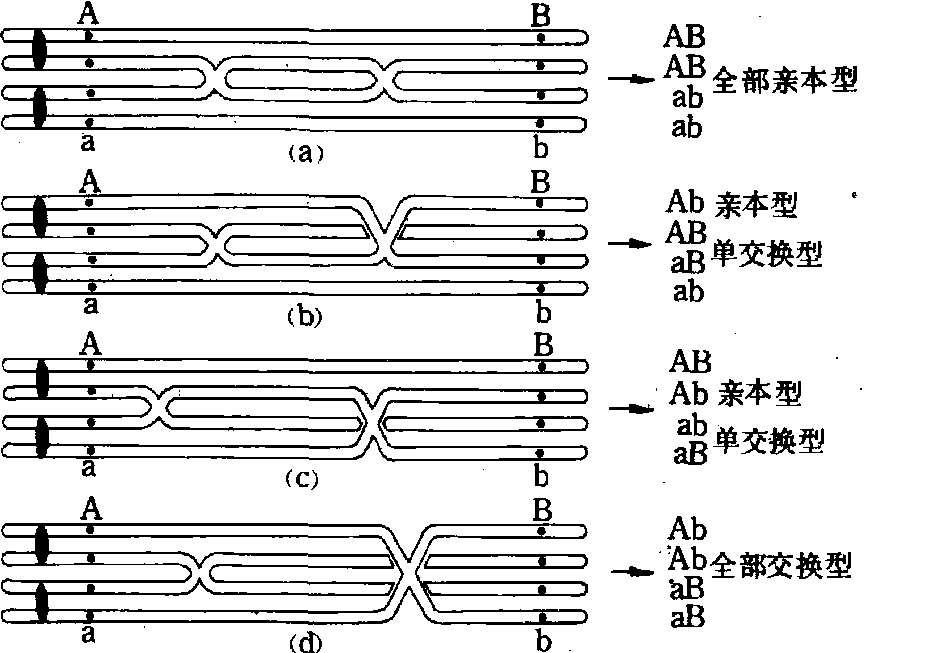
图 1 四种双交换模式图
染色体中一个区段发生的交换可能影响相邻区段的交换,称为干扰。1916年美国遗传学家马勒(H.J.Muller)首次在果蝇中发现干扰现象。干扰程度由实际双交换值与理论双交换值之比即符合系数测算,理论双交换值由两次单交换值之积求出,故干扰率=1-符合系数。干扰率小于1时为正干扰。自然界正干扰的现象极为普遍,而且基因间距离愈近干扰程度愈大。干扰率大于1称为负干扰。在噬菌体、细菌、真菌、酵母菌和家蚕中发现有负干扰现象。染色单体干扰会减少双线双交换的频率,有可能出现交换值大于50%的现象,在酵母菌、真菌和地钱中曾观察到。此外,还存在染色体间效应。1939年马瑟(K.Mather) 提出在一个细胞中交叉总数稳定的假说,即某一对染色体交叉增加时,另一对的染色体交叉就减少。
交换的细胞学基础直到1931年才为斯特恩 (C.Stern)在果蝇的试验中以及克赖顿(H.B.Creighton)和麦克林托克(B.McClintock)在玉米的试验中所证实。斯特恩选到一个具有标记染色体和标记基因的雌蝇,其一条X染色体载有肉色眼基因(Car)和十六区A段重复(B)但却缺失了另一段。另一条X染色体载有显性眼色基因并连接了一段Y染色体,表型为红色棒眼。这种雌果蝇与肉色正常眼的雄蝇交配,对子代进行了交换的遗传学鉴定和染色体细胞学的观察,获得了交换的细胞学证据(图2)。
交换发生的机制 1931年美国细胞遗传学家贝林(J. Belling)假设同源染色体在复制成染色单体期间,由于互换复制模板而发生交换。1937年英国细胞学家达林顿(C.D.Darlington) 认为同源染色体联会时相互缠绕,染色体间的扭力使各自的一条染色单体断裂,两个断头重接时发生非姊妹染色单体的交换。T4和λ噬菌体的染色单体断裂和重接并非机械力所致,而是由核酸内切酶、DNA聚合酶和连接酶等一系列酶参与作用所完成。
细菌的重组通过转导、转化和不同菌种的接合实现 (见转导、转化、细菌接合)。1946年德尔布鲁克(M.Delbriick)等选用带速溶噬菌斑基因r和能感染大肠杆菌B和B/2菌株的突变基因h作标记,将h+r与hr+两种T2噬菌体杂交 (即对大肠杆菌B株双重感染),所得子代再接种在含有B及B/2株的培养基上,根据噬菌斑性状选得重组型h+r+和hr,即两种性状属野生型和两种性状均属突变型的后代,计算出的重组值用于绘制连锁图。电子显微镜的观察也显示了噬菌体杂合DNA配对及交换重组的图像。
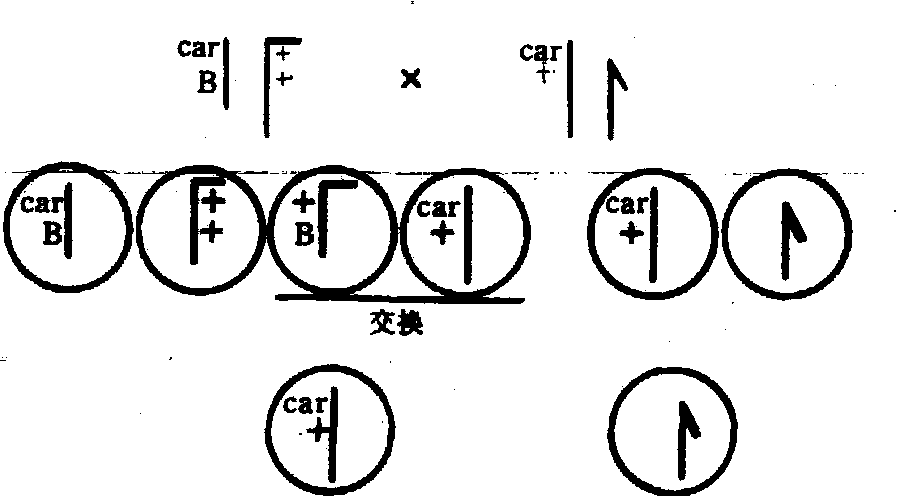
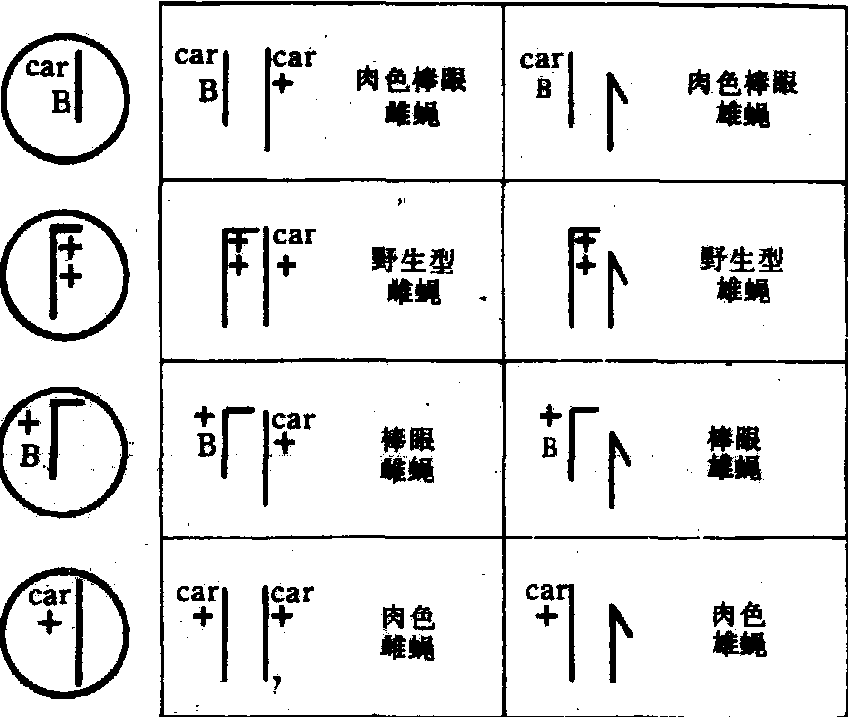
图 2 染色体交换的细胞学证据示意图
杂合体的体细胞染色体配对可能导致交换,出现性状嵌合现象。1936年美国遗传学家斯特恩(C.Stern)首先在果蝇中发现少数杂合雌蝇(sn+/+y)表现为有焦刚毛(sn)和黄体(y)孪生斑的嵌合体,是由于体细胞交换产生基因型为sn sn和yy的细胞的结果。1958年泰勒(J. H. Taylor) 应用同位素标记法首次报道细胞的有丝分裂过程中发生的姊妹染色单体互换现象。据研究,姊妹染色单体互换与DNA损伤有关,由于DNA修复功能缺陷导致染色体稳定性减弱而表现姊妹染色单体互换率升高。利用这种关系可用于测验某些先天性疾病以及检测环境中致癌、致畸和致变的诱变剂。
理论与实际意义 连锁与交换规律的发现奠定了遗传物质的染色体基础,开拓了遗传学发展的道路。目前已深入探讨染色体交换的分子机制,不同生物固有的连锁群保证物种的稳定性; 交换形成多样化的重组型,为生物进化提供了无尽的源泉。杂交育种的目的在于利用基因重组综合双亲的优良性状,选育新品种。预测目标性状间是否连锁以及根据交换率确定育种群体的大小,对提高杂交育种效率有十分重要的价值。此外,在人类遗传病的预测,产前诊断及环境保护等方面都有实用意义。
连锁与交换
父方或母方的某几种性状联合参加杂交,不论是显性或隐性,在子代中这些性状一般表现互相联合的趋势,这是因为他们的基因都在同一条染色体上。这种联合遗传的现象称为连锁。同对两组连锁性状(基因)之间,常常发生一定百分比的交换,这种现象称为交换,其机理在于减数分裂中,两个基因组所在的两条同源染色体之间,发生了有规律的片段交换。
摩尔根于1910年在野生型红眼果蝇中,发现一只白眼突变型雄果蝇,并通过杂交证实了“白眼”基因在遗传中的行动同一条X染色体的行动完全平行,于是断定“白眼” 基因就在这条X染色体上 (Y染色体没有它的等基因),因而随这条X染色体传给后代。以后,摩尔根等陆续发现果蝇的许多基因,如“黄体”、“朱眼”、“细翅”、“棒眼”等基因,均与“白眼”基因一样同在X染色体上。它们都随X染色体的行动联合传给后代。这种特殊的遗传方式最初称为性连锁遗传,以后称X连锁遗传。
连锁 以果蝇为例说明连锁现象,并从细胞学上说明其遗传机理。野生型果蝇呈灰身、长翅。突变型果蝇有黑身和残翅。杂交实验证明,灰身与黑身是一对相对性
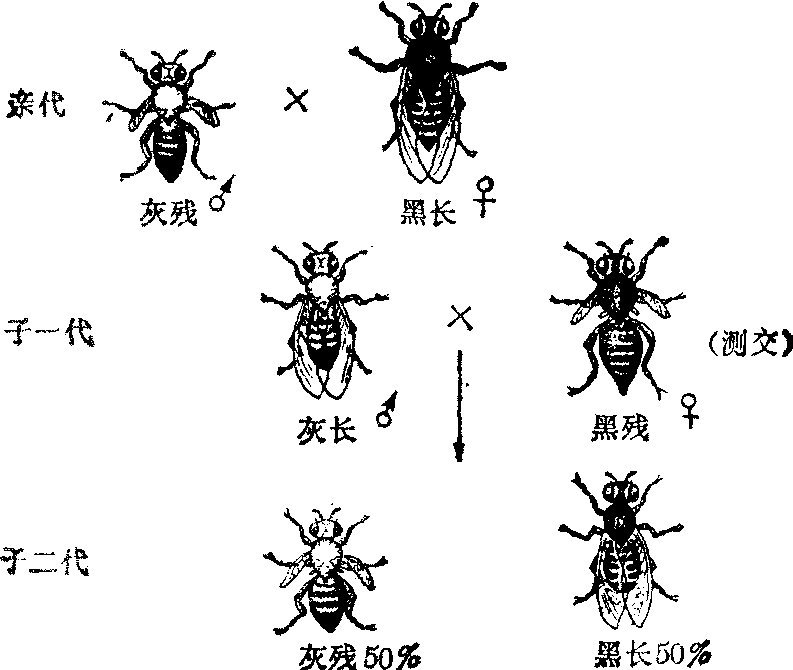
图1 果蝇连锁遗传
状,灰身是显性(基因B),黑身是隐性(基因b);长翅与残翅是另一对相对性状,长翅是显性(基因V),残翅是隐性(基因v)。以灰身残翅的雄蝇同黑身长翅的雌蝇杂交,子一代杂种完全是灰身长翅。再让子一代杂种雄蝇与黑身残翅雌蝇杂交。这种用纯合隐性型与待测定的显性表型杂交,以测验后者的基因型的方法称为测交。子二代个体和亲型完全相同: 一半是灰身残翅,另一半是黑身长翅。这种结果与孟德尔两对性状自由组合律(见“孟德尔定律”)完全不同。如果按自由组合律来预测(表1),子二代应该出现灰身残翅、黑身长翅、灰身长翅、黑身残翅四种类型,各占25%。但实验结果与预期结果并不一致。摩尔根把灰身和残翅、黑身和长翅分别联合遗传的现象称为连锁。这是因为“灰身”基因和“残翅”基因同在亲体传来的一条染色体上; “黑身”基因和“长翅”基因则在另一个亲体传来的一条同源染色体上(图2)。在子一代杂种生殖细胞成熟过程中,上述两条同源染色体彼此分离,黑长组合和灰残组合也随之分离,分别进入一个成熟的生殖细胞。因而子一代雄蝇只产生两种精子: 一种带有“黑身”基因和“长翅”基因的染色体; 另一种则带有“灰身”基因和“残翅”基因的染色体。与子一代杂种杂交的黑身残翅的雌蝇为隐性纯合体,其两条同源染色体上,都带有“黑身”基因和“残翅”基因; 减数分裂后,只产生一种带有“黑身”基因和“残翅”基因染色体的卵子。在测交中,子一代雄蝇的“灰身”、“残翅”精子和雌蝇的“黑身”、“残翅”卵子结合,将发育成灰身残翅的子二代; 子一代雄蝇的“黑身”、“长翅”精子和雌蝇的“黑身”、“残翅”卵
表1 按孟德尔自由组合律的预测结果
| 精子 卵 | BV | Bv | bV | bv |
| bv | BbVv 灰身长翅 | Bbvv 灰身残翅 | bbVv 黑身长翅 | bbvv 黑身残翅 |
| 比例 | 25% | 25% | 25% | 25% |
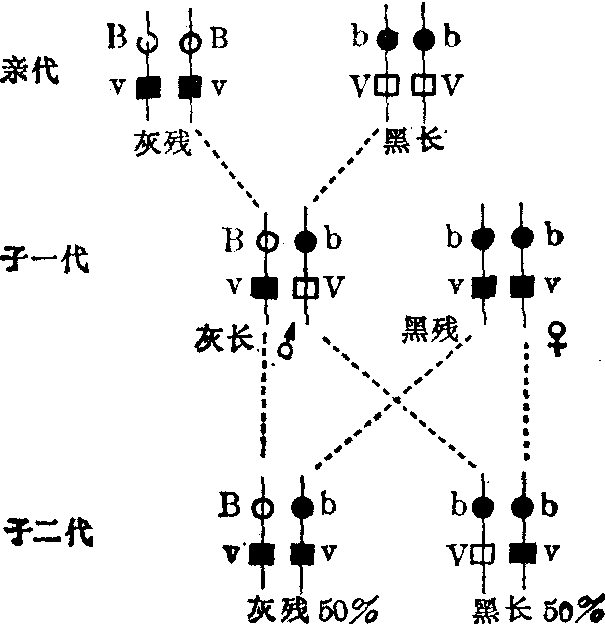
图2 果蝇基因连锁
○灰身基因显性,用B表示 ●黑身基因,隐性,用b表示
□长翅基因,显性,用V表示 ■残翅基因,隐性,用v表示
交换 用黑身残翅的纯合体雄蝇和上述实验的子一代杂合体雌蝇进行测交,子二代有四种类型; 其中两型与亲型相同: 一半为灰身残翅,另一半为黑身长翅; 两类的数量共占子二代的83%;另外两个类型为新的组合:半数为灰身长翅,半数为黑身残翅; 共占子二代17%。这两种新的组合称为重组型或交换型。子二代虽出现四型,但比例并不象自由组合那样各占25%。
摩尔根认为交换的染色体机理是,在子一代多数雌蝇生殖细胞第一次减数分裂中,载有“灰身”基因和“残翅”基因的一条染色体与载有“黑身”基因和“长翅”基因的另一条同源染色体,彼此分离,所以两组基因仍有83%保持原有连锁关系,这是子二代中与亲代相同的亲型组合;其余17%的是重组型或交换型,是由于在两条同源染色体上两个基因对之间发生了交换,使一条染色体上的“灰身”基因与另一条同源染色体上的“长翅”基因相连锁,前一染色体上的“黑身”基因与后一染色体上的“残翅”基因相连锁。因而,子二代中出现了灰身长翅和黑身残翅这两种重组型,这种情况称为不完全连锁(图3,图4)。
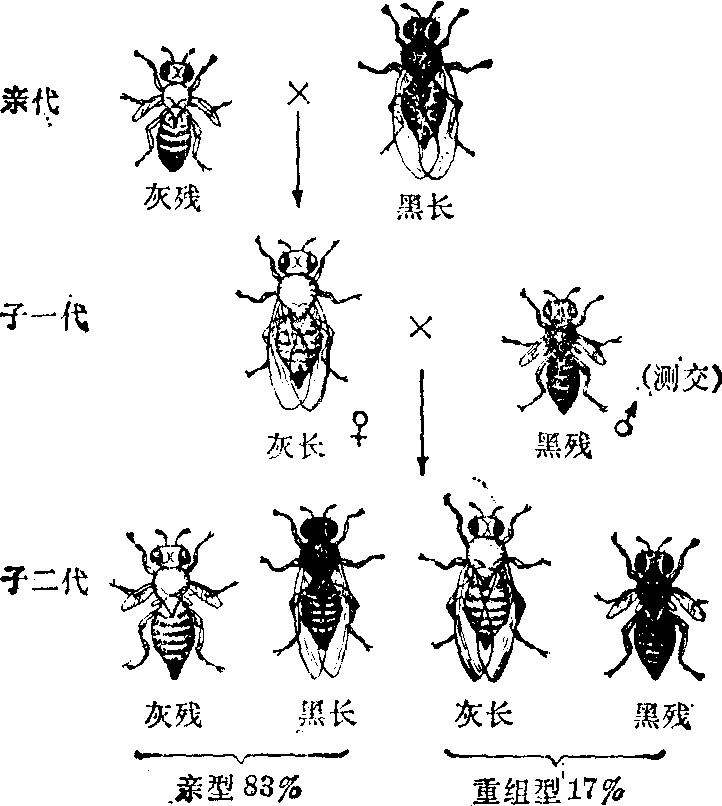
图3 果蝇连锁与交换
这种交换发生在雌蝇第一次成熟分裂中,在双线期,同源染色体之间形成一定数量的交叉,这就是发生过交换的细胞学表现。在雄蝇精母细胞减数分裂中看不到同源染色体之间的交叉现象,说明在雄蝇中连锁基因之间不发生交换,因此在交换实验中必须用杂合体雌蝇作为母方。
同源染色体上异对基因之间的交换从另一个细胞遗传学实验中得到充分证实。Stern(1931)培养出一种雌蝇,它的两条X染色体形状不同,容易识别,所含的相对基因也不相同。第一条X染色体断裂成两段; 一段上载有“粉红眼”基因(隐性)和“棒眼”基因(显性); 第二条X染色体和Y染色体一部分相连,形状象L。这一条X染色体上载有第一条X染色体的两个基因的等位基因——“红眼”基因(显性)和“圆眼”基因(隐性)(见“减数分裂”之图4)。Stern让红色棒眼的雌蝇同粉红色圆眼的雄蝇交配,产生四种类型的子蝇,经过细胞遗传学检验发现,多数子蝇中,一半为棒形粉红眼,具有一条断裂X染色体; 另一半为圆形红眼,具有一条L形的X染色体。这充分证明:“粉红眼”基因和“棒眼”基因在断裂X染色体上呈连锁关系; “红眼”基因和“圆眼”基因则在另—L形X染色体上呈连锁关系。此外,还有少数子蝇表现新的基因组合: 一半为圆形粉红眼,具有一条正常形状的X染色体;另一半为棒形红眼,具有一条L形断裂的X染色体。这说明本来是一条断裂X染色体上连锁着“粉红眼”基因和 “棒眼”基因,另一条L形X染色体上连锁着“红眼”基因和“圆眼”基因; 两条X染色体发生交换后,使“棒眼”基因交换到L形染色体上,“圆眼”基因交换到断裂的X染色体上,结果形成“粉红眼”基因和“圆眼”基因连锁; “红眼”基因和“棒眼”基因连锁。Stern证实: 两对基因之间的交换和它们所在的两条染色体相应部位之间的交换的一致性。
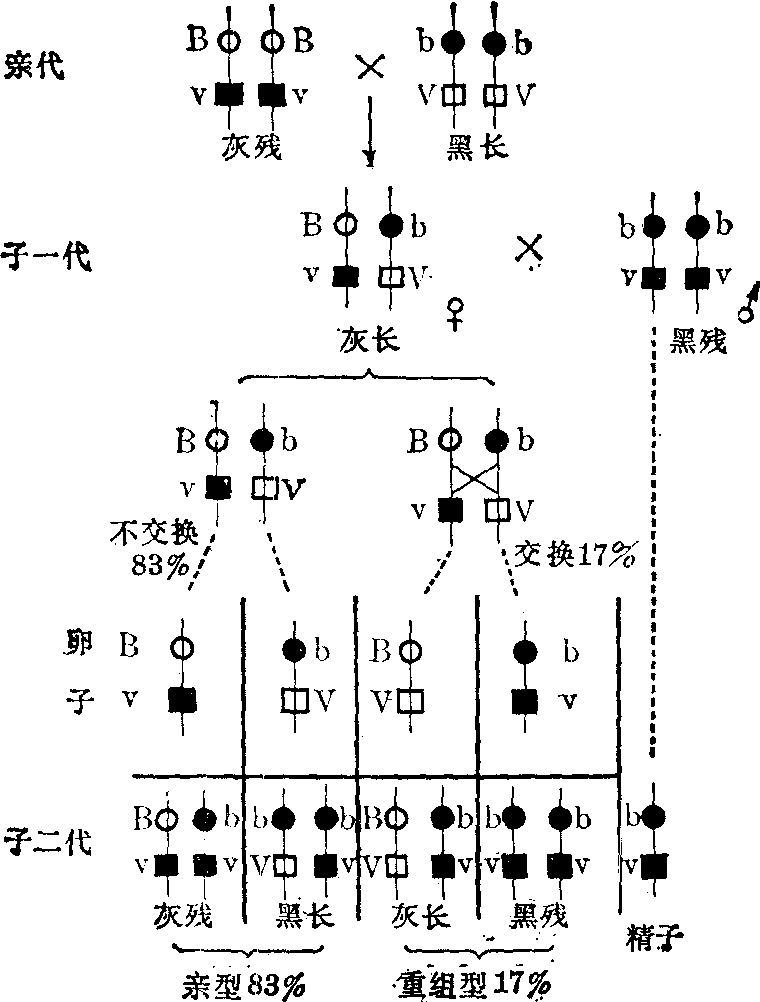
图4 果蝇基因连锁与交换
细胞遗传学研究发现,同源染色体之间可以发生不等交换,造成部分基因的重复和缺失。例如果蝇中,由野生型圆眼突变为棒眼; 棒眼(突变型)又可以还原成正常眼(野生型),就是染色体的不等交换所致。
Sturtevant根据杂交结果断定,棒眼果蝇还原成圆眼果蝇,是因为载有棒眼的两条X染色体之间发生了不等交换,以致一条X染色体失去它的棒眼基因,使子代雄蝇成为正常的圆眼; 而另一条X染色体则载有两个棒眼基因,使子代雄蝇复眼缩得更细小(称双棒眼)。
Bridges(1936) 在果蝇染色体的研究中证实了Sturte-vant上述推断的正确性。Bridges发现在普通圆眼果蝇的唾液腺X染色体的有关片段上约有6条横带。在棒眼果蝇的X染色体上,这一片段重复了一次 (图5,图6)。
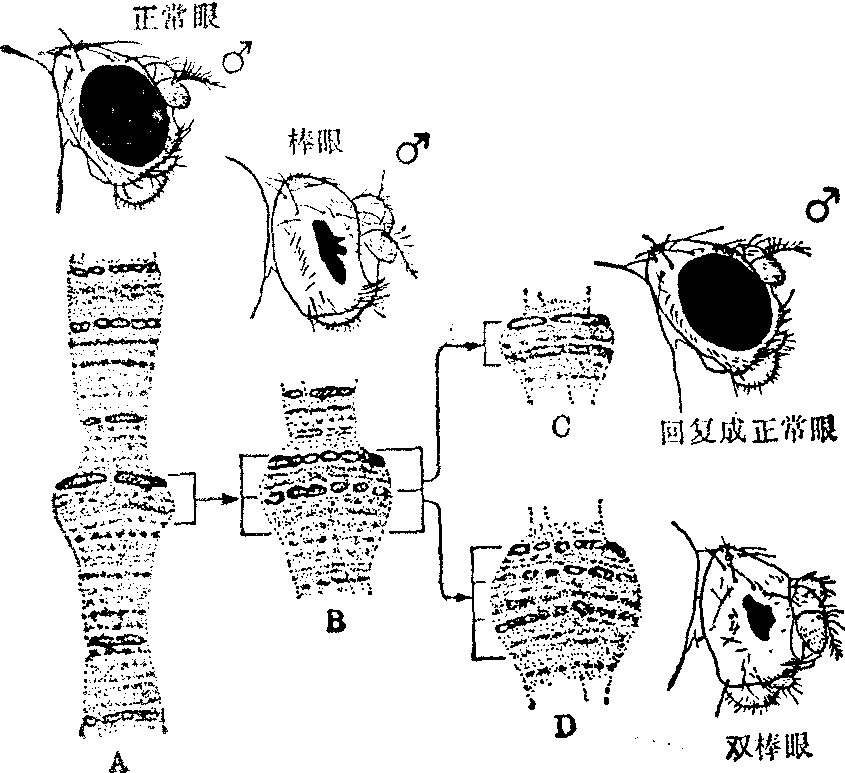
图5 棒眼基因还原为圆眼基因
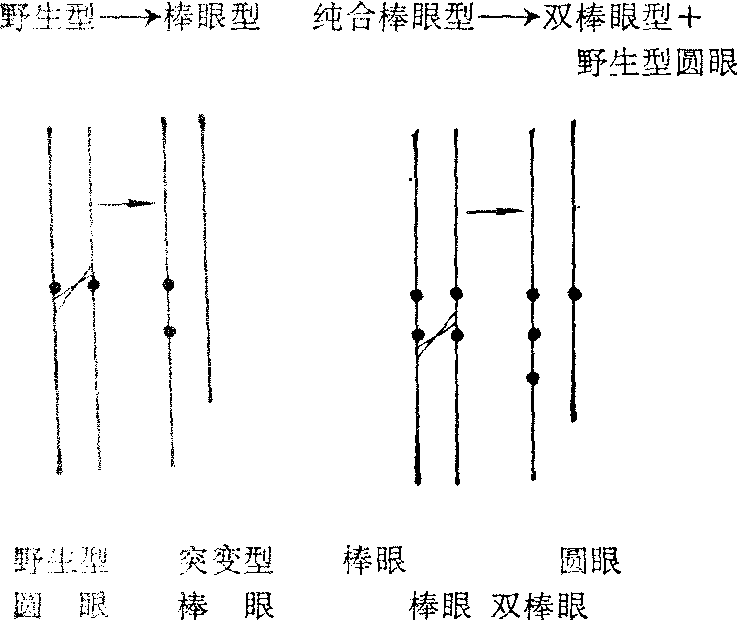
图6 减数分裂中不等交换
在双棒眼果蝇的X染色体上,这一片段则重复了两次。由棒眼型还原所形成的正常型果蝇,它的X染色体只有一个片段,其带纹与普通圆眼果蝇X染色体上“圆眼”基因区的带纹相同。由此证明在棒眼果蝇的两条X染色体之间,确实发生了不等交换。
基因的不等交换也可能发生在人体细胞中。例如人类的一种异常血红蛋白分子病——Hb Lepore,临床表现为β-海洋性贫血病症状。患者的血红蛋白分子中出现一种异常的δ-β链,即δ链氨基端的一个节段同β链羧基端的一个节段连接而成的多肽链。从δ-β多肽链结构的特点推测,它的结构基因也是由δ基因的一部分和β基因的一部分连接而成的δ-β融合基因。根据Hb基因位点关系已知δ基因与β基因同是一条染色体的连锁基因,这样的两条同源染色体之间在减数分裂时发生不等交换,交换发生在一条染色体的β基因位点和另一条同源染色体的δ基因位点之间,因而δ基因与β基因交换了部分片段,结果产生一个δ-β融合基因,并产生另一个与之对应的β-δ融合基因 (见“血红蛋白病与血浆蛋白病”)。由δ-β融合基因决定蛋白质合成,产生异常的δ-β多肽链。
连锁群 到1928年止,培养的果蝇中已先后出现了400多种基因突变,影响蝇体内外各种特性。根据杂交实验结果,这些基因分成4个连锁群:第1连锁群包括全部性连锁基因,说明他们都在X染色体上;第2、第3连锁群的基因最多,与之相应的两对染色体也最长; 第4连锁群只有少数几个基因,第4对染色体也最短。由此可知,基因连锁群数同染色体对数相等; 各连锁群内基因的多少又与有关染色体的长短相对应。其他生物凡是经过遗传学上充分研究的,其连锁群数均等于染色体对数即单倍染色体数; 未经过充分研究的,也有低于染色体对数的。人类有23对染色体,每对染色体上都有其特定基因,分属于包括Y连锁基因在内的24个连锁群。随着研究的深入,有关连锁数据的增多,可以预料,人类各个连锁群的连锁基因必将与年俱增。
表2 几种生物的连锁群与染色体对数
| 生物类型 | 连锁群数 | 染色体对(单倍)数(n) |
| 黑腹果蝇 玉 米 大 麦 豌 豆 链孢霉 小 鼠 家 兔 家 蚕 | 4 10 7 7 7 19 11 22 | 4 10 7 7 7 20 22 28 |
基因定位与遗传图 见“基因定位”。
- 指典舆是什么意思
- 指典袍是什么意思
- 指兽发踪是什么意思
- 指冀是什么意思
- 指冠是什么意思
- 指冬瓜说葫芦是什么意思
- 指冬瓜骂葫芦是什么意思
- 指冬瓜骂葫芦,指和尚骂秃(子)是什么意思
- 指冬瓜,骂葫芦是什么意思
- 指冻无完甲,琴断有折弦。是什么意思
- 指准是什么意思
- 指凑是什么意思
- 指出是什么意思
- 指出事物的本质所在是什么意思
- 指出事物的根源是什么意思
- 指出人的姓名是什么意思
- 指出优点和缺点是什么意思
- 指出对方的罪行,加以谴责是什么意思
- 指出或标出是什么意思
- 指出来使人知道是什么意思
- 指出来给人看是什么意思
- 指出犯规者是什么意思
- 指出缺点、毛病是什么意思
- 指出让人注意是什么意思
- 指出证据是什么意思
- 指出谬误,加以改正是什么意思
- 指出过失,加以批评是什么意思
- 指出错误是什么意思
- 指出错误以改正是什么意思
- 指出错误以求改正是什么意思
- 指击是什么意思
- 指刀是什么意思
- 指分是什么意思
- 指切是什么意思
- 指切压手法是什么意思
- 指切押手法是什么意思
- 指切进针法是什么意思
- 指划不来是什么意思
- 指划动作是什么意思
- 指列是什么意思
- 指别词是什么意思
- 指到哪儿打到哪儿是什么意思
- 指到黄牛便是马是什么意思
- 指刺是什么意思
- 指办是什么意思
- 指动是什么意思
- 指北针是什么意思
- 指南是什么意思
- 指南、手册是什么意思
- 指南公是什么意思
- 指南后录是什么意思
- 指南宫是什么意思
- 指南录是什么意思
- 指南录后序是什么意思
- 指南总论是什么意思
- 指南打北是什么意思
- 指南摘要是什么意思
- 指南攻北是什么意思
- 指南方是什么意思
- 指南村志是什么意思