细胞质基质cytoplasmic matrix
细胞质内除去细胞器和内含物以外的胶状物质。即生化学家用分级离心法得到的上清液部分,又称细胞溶质。
最初,细胞质基质仅指光镜下能看到的所有细胞器以外的透明质部分。以后在电镜下发现,细胞质基质中还含有许多细胞器,如由微管、微丝、居间丝相互交错成网的细胞骨架,它对细胞运动和形态建成均有重要作用。20世纪70年代,美国细胞学家波特(K.R.Porter)在高压电镜下,观察到细胞质基质中还存在更为纤细的微梁网络系统(见图)。
细胞质基质的主要组成成分是蛋白质,占20~25%,包括糖酵解、脂肪酸合成、核苷酸代谢及氨基酸活化所需要的酶类。还有脂类、糖类、无机盐及转移核糖核酸(tRNA)、信使核糖核酸(mRNA) 和蛋白质合成的有关因子。细胞质基质呈复杂的胶体性质,可因环境条件的变化,在溶胶态与凝胶态间互为转换。
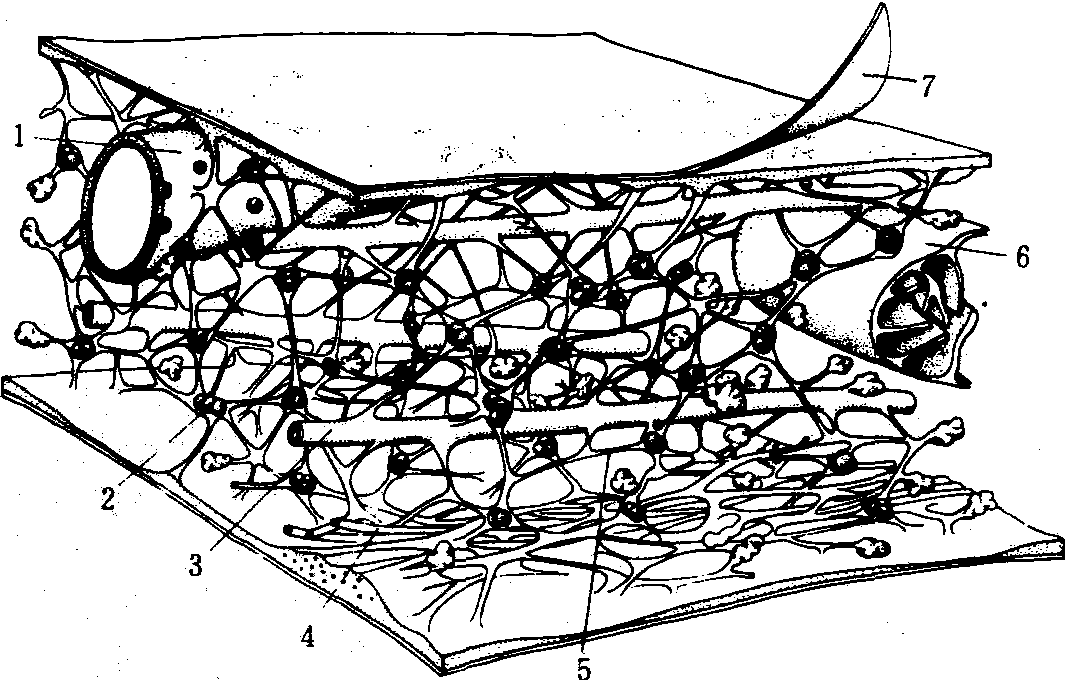
细胞微梁网络模式图
1. 内质网; 2. 核糖体; 3. 微管; 4. 微丝;5. 微梁网络; 6. 线粒体; 7. 质膜
细胞质基质充满细胞,因此是内膜系统和细胞器的内环境,同时这些膜性结构和细胞器的基本组成成分都依靠细胞质基质提供,所以又是它们形态发生的源泉。
根据波特(K. R. Porter)等的观察,微梁网络在细胞内可随细胞生理功能和细胞环境的改变而变化。如低温(4℃)时培养细胞呈球状,与此相应的内部骨架系统发生有序拆卸,首先是微管,然后微丝,最后是微梁网络(虽然不完全)解体; 37℃下5分钟后又重新构建而恢复原状。微梁网络在细胞内可能有组织细胞质基质内各种代谢酶的作用,如糖酵解途径中有关的酶,并非随机地与微梁网络相结合,而是有序地排列,使一个酶将其反应产物就近传给代谢途径中的下一个酶作为底物,不与膜结合的酶也有可能在空间协作起来。在蛋白质合成中,游离核糖体位于微梁网络的交叉处,蛋白质一旦被制造出来,就结合于微梁网络,然后以非随机方式提供微管、微丝或其他细胞结构的受控装配。另外细胞内色素颗粒的集散,可能也与微梁网络的变化有关。但至今从细胞质成分中尚未分离到组成微梁网络所特有的蛋白质,因此对它的存在尚有争议。
细胞质基质cytoplasmic matrix
细胞质中除细胞器和内含物以外的胶态基底物质。包括:
❶组成细胞骨架的3种主要纤丝——微丝、微管和中间纤维;
❷由纤丝桥所组成的相互交联的丝状结构——微梁系统;
❸糖、氨基酸、蛋白质、无机盐、水等有机物和无机物,并由这些物质组成一个有序的、各种成分相互协作的组织结构。为细胞内分子合成、能量转换、细胞分化和代谢、细胞器行使功能等提供所需的环境,参与细胞许多动力活动过程,如细胞分裂、变形运动和一些细胞的病变过程等。
细胞质基质cytoplasmic matrix
亦称胞溶质。指细胞质内无定形的胶状物质,内含多种可溶性酶,如糖酵解酶以及与氨基酸合成和分解有关的酶等。
细胞质基质
细胞质基质为除去细胞器和包含物等可见结构之外的细胞质,呈均质透明,简称基质。基质为细胞的重要组成部分。各种细胞器的生理活动、细胞的变形运动、吞噬作用和细胞分裂等均与细胞质基质有密切关系。
细胞质基质由水、无机盐、碳水化合物、蛋白质(包括各种酶类)、脂类和核酸等构成。这些化学成分的比例大致为,水分75%、蛋白质20%、脂类3%、碳水化合物1%、无机盐1%。但各种成分的比例可随细胞类型的不同,而不一致。水是细胞质基质的主要成分,为许多溶质的生物溶媒和活动介质,与基质内的物质代谢过程直接相关。除水分之外,蛋白质也是细胞基质的重要成分。蛋白质中包括许多催化物质代谢的酶,如参加无氧酵解的酶系,脂肪酸激酶,参加蛋白质生物合成的各氨基酸激酶以及一些结构蛋白。结构蛋白在一种条件下可为球状蛋白,而在另一种条件下则能聚合成为纤维状蛋白,再构成某种微细结构。如管蛋白能聚合成为原丝,再构成微管;肌动蛋白能聚集为细丝,再组成微丝。脂类和核酸,主要与蛋白质相结合,以脂蛋白或核蛋白的形式存在。细胞质基质中的钾、钠、氯等无机离子对维持细胞内外的渗透压及酸碱度具有重要作用。
细胞质基质可被视为胶体系统,但由于其胶体特性比较复杂,不少现象又不能以一般的胶体性质予以说明,所以又被看成是一个高度异质性的胶体系统。细胞质基质具有粘滞性。这种粘滞性与基质的胶体性质有关。当基质的胶体呈凝胶状态时,粘滞性增高;反之,当胶体处于溶胶状态时,粘滞性便降低。 据认为凝胶⇄溶胶的变化,可能与基质中结构蛋白的聚合和解聚有关。细胞质基质的粘滞性可随细胞内外的条件因素而发生改变。如变形虫的细胞质基质在低渗溶液中粘滞性降低。而在超过30℃的条件下粘滞性便急剧增加。压力因素也可改变细胞基质的粘滞性,如海胆卵分裂时,在分裂沟会出现凝胶环,但当施以压力时,凝胶环则呈溶胶状态。细胞质基质的酸碱度一般略偏于酸性,pH约6.8。当向细胞质基质中加入酸性或碱性物质后,其pH值便会发生改变。但是,只要细胞质基质的生活力没有改变,便会依靠其缓冲能力,使酸碱很快地恢复到原来的数值。
从细胞质基质的物理化学特性来看,它在分子水平上必定具备某种结构。但是由于对细胞质基质的微细结构研究得较少,目前尚提不出明确的看法。有人试以结构键的学说来说明细胞质基质的结构。认为细胞质基质中的长链状蛋白质分子的一些侧链,可通过各种键相互连接起来,形成分子网。一些亲水基团,水或疏水基团可依其性质的不同与蛋白质侧链末端的基团结合; 无机离子也可依其荷电性质的不同而被吸引到蛋白质侧链末端的酸基或碱基周围。这样,水、无机离子和脂类等便都被配列到适当的位置上,而使细胞质基质在分子水平上形成一定结构。 细胞质基质的凝胶⇌溶胶变化, 可能是由于细胞内外条件的改变,而使侧链的连接键发生了结合或拆开而造成的。连接键拆开的多,细胞质基质粘滞性降低,而显示溶胶状态;反之,连接键结合的多,粘滞性增高而呈现凝胶状态。在正常生理条件下,总是有些连接键结合,而另一些连接键拆开,从而使细胞质基质具备一定的弹性。
- 诗兴浑飞在渔乡橘里。是什么意思
- 诗兵是什么意思
- 诗具是什么意思
- 诗册是什么意思
- 诗写得出神入化是什么意思
- 诗准是什么意思
- 诗凭写兴忘工拙,酒取浇愁任浊清。是什么意思
- 诗出乎志者也,乐出乎诗者也是什么意思
- 诗出乎志者也,乐出乎诗者也。是什么意思
- 诗函是什么意思
- 诗切是什么意思
- 诗切不分卷是什么意思
- 诗刊是什么意思
- 诗刊放假是什么意思
- 诗创作是什么意思
- 诗创作社是什么意思
- 诗创造是什么意思
- 诗创造社是什么意思
- 诗到无人爱处工是什么意思
- 诗刻是什么意思
- 诗前哨是什么意思
- 诗剧是什么意思
- 诗剧(1)是什么意思
- 诗剧(2)是什么意思
- 诗力是什么意思
- 诗势是什么意思
- 诗勋是什么意思
- 诗化小说是什么意思
- 诗化散文是什么意思
- 诗化舞蹈的先驱是什么意思
- 诗匠是什么意思
- 诗十首是什么意思
- 诗南是什么意思
- 诗印是什么意思
- 诗卷是什么意思
- 诗卷光阴楼是什么意思
- 诗原是什么意思
- 诗友是什么意思
- 诗双声叠韵谱是什么意思
- 诗双声叠韵谱一卷是什么意思
- 诗叟是什么意思
- 诗叟未相识,竹儿争见君。是什么意思
- 诗古微是什么意思
- 诗古微17卷是什么意思
- 诗古微十七卷是什么意思
- 诗古徽是什么意思
- 诗古训补遗是什么意思
- 诗古音是什么意思
- 诗古韵表二十二部集说是什么意思
- 诗古韵表二十二部集说二卷是什么意思
- 诗古韵表廿二部集说是什么意思
- 诗句是什么意思
- 诗句中最精练传神的一个字是什么意思
- 诗句名句是什么意思
- 诗句名句 / 诗词名句1是什么意思
- 诗句名句 / 诗词名句10是什么意思
- 诗句名句 / 诗词名句11是什么意思
- 诗句名句 / 诗词名句12是什么意思
- 诗句名句 / 诗词名句13是什么意思
- 诗句名句 / 诗词名句14是什么意思