神经递质
一个神经元的信息传递到下一个神经元时,必需释放化学物质做为中介。神经元支配效应器官(例如骨骼肌、平滑肌和心肌等)活动时,其轴突末梢也释放化学物质做中介,再将其信息传递到这些组织。神经元传递信息所借助的化学物质就称为神经递质。大多数神经元轴突末梢释放的递质是在神经末梢处合成的,也有在细胞体处合成后运送到轴突末梢去的。递质常常贮存在突触小泡内,以防止被细胞浆中的酶将其催化而破坏。但当递质释放并与下一个神经元膜上的受体结合发挥作用后,却迅速分解或流走。这一点也很重要,它决定了神经调节的灵活性,保证神经元的作用可迅速出现并立即终止。一个神经元兴奋时,其轴突的各个分支末梢所释放的递质是相同的,但不同的神经元,其轴突末梢释放的递质则可以不同。有的释放乙酰胆碱,有的释放去甲肾上腺素,有的释放多巴胺,有的释放内源性阿片样物质等。生理上依照神经末梢释放的递质不同,将神经元分为乙酰胆碱能神经元、去甲肾上腺素能神经元……等。目前已知的递质已超过20种,但在中枢神经系统中,哪些神经元释放哪种递质目前知道的仍是少数。对周围神经系统的神经末梢释放的递质了解较多,主要有两种(乙酰胆碱和去甲肾上腺素)。各种递质的神经元在中枢神经系统中都不是均匀分布的,而是相对集中在某些区域,并完成一定的机能。例如脑干的乙酰胆碱能神经元,其轴突末梢到达大脑皮层,它们兴奋时可出现觉醒。
神经递质neurotransmitter
传递神经冲动的一种化学物质。由神经末梢释放,作用于次一级神经元或效应器(即心脏、平滑肌等)的受体,完成神经冲动的传递过程。传出神经递质主要有乙酰胆碱、去甲肾上腺素等。中枢神经递质包括乙酰胆碱、去甲肾上腺素、多巴胺、5-羟色胺、γ-氨基丁酸、甘氨酸、谷氨酸、组胺、脑啡肽等。
神经递质neurotransmitter
又叫化学递质,系神经细胞之间或神经细胞与效应细胞之间起传递作用的化学物质。此类递质分中枢递质和外周递质:中枢神经元间递质有单胺类、肽类、氨基酸类、乙酰胆碱等;外周递质有去甲肾上腺素、乙酰胆碱、肽类和嘌呤类。递质能与受体发生特异性结合而传递冲动,发生生理效应。递质的作用不同;对下个神经元起兴奋作用的叫兴奋性递质;对下个神经元起抑制作用的叫抑制性递质。递质产生多少及性质有否变化,可影响神经功能,并产生相应的临床症状;医疗上已应用不同递质治疗某些神经系统疾病。
神经递质
在高等动物神经末梢的突触处,信号的传递是通过突触前神经元释放某种化学传递物质,即所谓“神经递质”(简称“递质”)来实现的。当神经递质与突触后神经元膜上的受体结合,可使之出现兴奋或抑制效应,从而完成信号的传递作用。
作为一种神经递质至少应具备下列基本条件和作用:
❶应存在于突触前神经元中,并能在此神经元中合成;
❷必须贮存在神经末梢的囊泡中,不致被胞浆内酶所破坏;
❸当神经冲动通过时能迅即从突触前膜释放进入突触间隙;
❹释放的递质能作用于突触后膜上特异的受体,并激发突触后膜离子通透性和电位的变化,从而将冲动信息传递给突触后神经元;
❺发挥上述效应后,应能迅即被终止活性或灭活。灭活的方式可以是被特异的酶所降解或被突触前膜、后膜和其他组织“重摄取”,也可以进入血循环进而被酶所降解。这些也是各种神经递质所共有的特征和“生命周期”。
早巳确知乙酰胆碱及去甲肾上腺素是植物神经及运动神经、肌肉接点的递质。关于中枢神经系统中的递质认识较晚。目前比较确定和尚在拟议中的递质有: 乙酰胆碱、单胺类(包括去甲肾上腺素、多巴胺、5羟色胺及组胺)、氨基酸类 (包括γ氨基丁酸、谷氨酸、天门冬氨酸及甘氨酸)以及某些肽类(P物质、脑啡肽及内啡肽等)。其中以乙酰胆碱、去甲肾上腺素、多巴胺、5羟色胺、γ氨基丁酸及甘氨酸等研究较深入,能较全面的满足上述基本条件外,其它也或多或少曾提出某些证据,但仍须进一步研究。
乙酰胆碱 早已发现乙酰胆碱(Ach)参予周围神经突触的信息传递,它是运动神经、交感神经节前纤维以及副交感神经节前及节后纤维突触的神经递质。现已证明它也是中枢神经系统中一种典型的递质,大脑的中隔可能是胆碱能神经元胞体所在的一个重要部位。
乙酰胆碱的生物合成是由胆碱乙酰化酶催化,从乙酰辅酶A转移乙酰基给胆碱,使之乙酰化而实现的。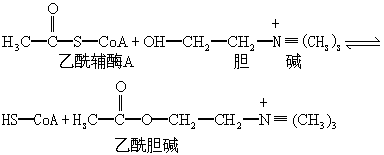
胆碱的供应是乙酰胆碱合成的限速因素。正常脑组织中游离胆碱仅20~200μmol/L,远比胆碱乙酰化酶与胆碱结合的Km值(4.5×10-4mol/L)低,即在正常情况下此酶远未能被作用物——胆碱所饱和。为了保持胆碱能神经元中乙酰胆碱正常水平,必须有外源性胆碱供应。但胆碱不易透过血脑屏障,须先经磷酸化,再转变为磷脂酰胆碱或其去脂肪酸产物(图1),才能通过血脑屏障,并在脑内水解生成胆碱,这一过程比较缓慢。在生理情况下胆碱的迅速补充可能还来自释放的乙酰胆碱,被突触间隙中乙酰胆碱酯酶水解,由神经末梢“重摄取”水解生成的胆碱。胆碱的重摄取和再利用对突触内乙酰胆碱的迅速合成有重要意义。
合成乙酰胆碱的乙酰基是来自葡萄糖氧化代谢,在线粒体内生成的乙酰辅酶A。另外游离乙酸在线粒体酶一乙酰辅酶A合成酶的催化和ATP参予下,可与辅酶A结合,也生成乙酰辅酶。线粒体内的乙酰辅酶A,还须合成为柠檬酸或乙酰肉毒碱,才能转运至胞浆,参予乙酰胆碱的合成。神经组织中乙酰胆碱的代谢见图1。
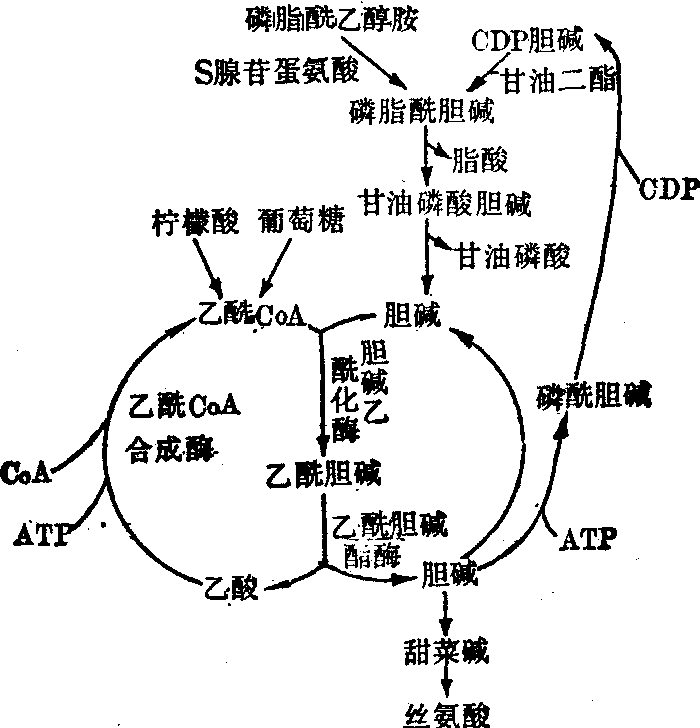
图1 神经组织中乙酰胆碱的代谢
胆碱乙酰化酶是一较稳定的碱性蛋白质,在中枢神经系统中主要分布在尾核、脊髓根中,从鼠脑提取纯化的酶分子量为65 000,主要存在于胞浆中,并可能与微粒体结合。它在神经末梢靠近突触前膜部位含量最高,这与乙酰胆碱在突触前神经元内迅速更新有关。
突触前神经元内合成的乙酰胆碱被囊泡摄取而贮存,每一囊泡约贮存104个乙酰胆碱分子,当神经冲动通过时,小泡所含有的乙酰胆碱从突触前膜全部释放排出,称为“量子式解放”。释放小泡的数量决定于突触前膜电位,换言之,乙酰胆碱的释放是一种电控分泌方式。这种释放还依赖于细胞外液的Ca2+,当突触前膜去极化时,Ca2+进入膜内,促进囊泡膜与突触前膜融合,从而使囊泡内容物排出。
乙酰胆碱的降解主要通过乙酰胆碱酯酶反应,这也是乙酰胆碱迅速灭活的主要方式。此酶普遍分布在靠近突触膜的附近,在神经细胞内结合在线粒体及微粒体的膜结构上。它的分子量约260 000,其活性中心包括两部分:
❶负离子部位,至少含一个谷氨酸的羧基,解离带负电荷,从而能将乙酰胆碱的季胺氮结合在酶分子上;
❷水解部位,包括组氨酸的咪唑基亲核基团及丝氨酸的羟基,能使乙酰胆碱从羰基碳原子处水解断裂。
乙酰胆碱释放后作用于突触后膜上的受体。受体主要有二种类型,一类可被毒蕈碱所激动,称为毒蕈碱样受体或M型受体; 另一类易为烟碱所激动,称为烟碱样受体或N型受体。N型受体分布在脊椎动物的神经、肌肉接点、植物神经节及小脑等部位; M型受体分布在中枢神经系统的大脑皮质及纹状体。在脑的某些部位尚发现同时兼有两种受体者。从电鱼的电器官及脊椎动物骨胳肌抽提纯化的N型受体蛋白质是一分子量为250 000~300 000的寡聚体,可能由5或6个亚基构成。亚基的分子量为40 000,其N末端为丝氨酸。乙酰胆碱受体蛋白质还含有中性糖3~5%。至于M型受体的本质,目前了解不多。儿茶酚胺 儿茶酚胺是指含有儿茶酚核的胺类化合物的总称,在神经化学中仅指多巴胺(即二羟苯乙胺)、去甲肾上腺素及肾上腺素三者,现认为多巴胺(DA) 及去甲肾上腺素(NE) 是作为哺乳动物的神经递质而发挥生物活性,而肾上腺素则为一激素,在两栖类动物,肾上腺素也是一种递质。
去甲肾上腺素不仅是交感神经末梢的递质,而且还是一重要的中枢神经递质。在脑干及下丘脑含量最多。去甲肾上腺素能神经元的胞体主要集中在桥脑及延脑的蓝斑核。多巴胺则主要是作为中枢神经递质,在哺乳动物脑中80%分布在基底神经核。
儿茶酚胺是从L酪氨酸通过一系列酶促反应合成的,其合成的主要途径如图2。
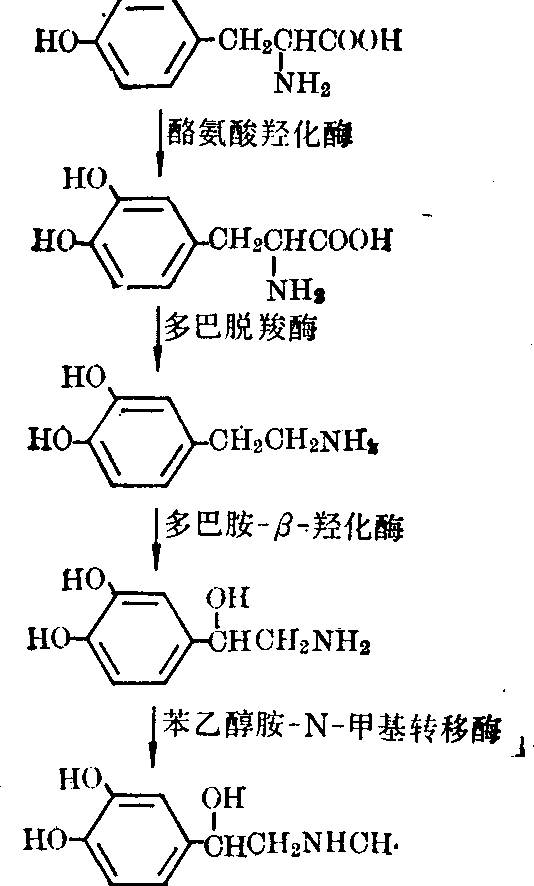
图2 儿茶酚胺的合成
结构式自上而下,依次为: 酪氨酸、多巴、多巴胺、去甲肾上腺素、肾上腺素酪氨酸是合成儿茶酚胺的直接前身,血液中酪氨酸通过特殊运载系统转运进入脑内,其他芳香族氨基酸对酪氨酸转运有竞争抑制。因此食物中氨基酸成分可间接影响脑内儿茶酚胺及其他单胺含量。在哺乳动物的中枢神经系统中,不仅有去甲肾上腺素能神经元,也有多巴胺能神经元。前者除含有合成多巴胺的酶体系外,还含多巴胺β羟化酶,使多巴胺羟化转变为去甲肾上腺素,而在多巴胺能神经元中则无此酶。这二类神经元中都没有苯乙醇胺N甲基转移酶,故都不能合成肾上腺素,因此脑内无或很少肾上腺素。多巴胺β羟化酶分布在神经末梢的囊泡中,其他酶都是在胞浆中。
酪氨酸羟化酶: 催化酪氨酸,在苯环的间位上羟化生成多巴(DOPA),此酶反应需要分子氧、Fe2+及四氢叶酸或二氢生物蝶呤或6甲基四氢蝶啶作为辅助因子。此酶的特异性很高,几乎只能使L酪氨酸羟化,与L酪氨酸结合的Km值为0.4×10-5mol/L。虽然在生理情况下它经常处于被作用物饱和的状态,但催化的反应是儿茶酚胺合成反应系列中最慢的一步反应,所以它仍是合成过程中的限速反应。
多巴脱羧酶: 催化多巴α脱羧,生成多巴胺,特异性不高,几乎可作用于所有天然的芳香族L氨基酸,包括酪氨酸、色氨酸、苯丙氨酸以及组氨酸、5羟色氨酸等,故又称为“L芳香族氨基酸脱羧酶”。脑中此酶活性很强,分子量为109000,需磷酸吡哆醛作辅酶,与多巴结合的Km值为4×10-4mol/L,作用物上有未被取代的氨基,是酶的特异性所要求的,这可能是与酶或辅酶结合的部位。
多巴胺β羟化酶(DβH): 催化的反应是去甲肾上腺素生物合成最后一步反应,作用于多巴胺使之在侧链的β碳上羟化,与多巴胺结合的Km为5×10-3mol/L,还能催化某些苯乙胺(如酪胺)生成相应的β羟化产物。此酶为一含铜的蛋白质,分子量为29000,每摩尔酶蛋白中含2Cu2+,同时还需分子氧及抗坏血酸作辅助因子。在这里Cu2+可能参予反应中的电子传递,而抗坏血酸则系电子供体。
关于儿茶酚胺作用的受体,根据对拟肾上腺素药物的反应不同,分为α受体及β受体,α受体对肾上腺素作用最强,β受体对异丙肾上腺素作用最强。最近又进一步把儿茶酚胺受体分为α、β1、β2及DM(多巴胺)四种受体,主要根据对去甲肾上腺素、肾上腺素、异丙肾上腺素及多巴胺的反应差异而分类的。
儿茶酚胺的降解或灭活可通过:
❶线粒体膜上的单胺氧化酶(MAO)催化,使儿茶酚胺氧化脱氨生成相应的醛;
❷分布在神经元外或其他组织中的儿茶酚胺O甲基转移酶(COMT)催化,转移甲基至儿茶酚胺的间位羟基上,生成相应的3甲氧化合物。放射性核素实验证明大约40~60%的儿茶酚胺是经这两种酶促反应的代谢而灭活,其余则是通过神经末梢的重摄取而灭活。这种重摄取能对抗浓度梯度,是主动转运过程,需消耗能量和依赖Na+的存在。
5羟色胺 5羟色胺(5HT)是一种具强生物活性的吲哚衍生物,能使平滑肌兴奋、血管收缩,并参予血凝固。在自然界分布很广,许多水果及蔬菜,如菠萝、香蕉中含有大量5羟色胺。在哺乳类动物的松果体、肠嗜铬细胞及血小板中含5羟色胺量很高。1953年才证明5羟色胺也存在于脑中,已有证据表明5羟色胺也是中枢神经系统中传递冲动信息的递质,并可能参予针刺镇痛过程。5羟色胺能神经元的胞体主要位于中脑、桥脑不同部位的中缝核群中,从这些核团发出的轴索几乎到达脑的所有部位,包括脑干网状结构、下丘脑、纹状体、边缘前脑及新皮质等,下行纤维投射到脊髓各级水平的灰质中。
5羟色胺不能通过血脑屏障。脑中5羟色胺全部是在神经元内合成。首先是L色氨酸羟化生成5羟色氨酸,然后再脱羧即生成5羟色胺。其生物合成反应如图3。
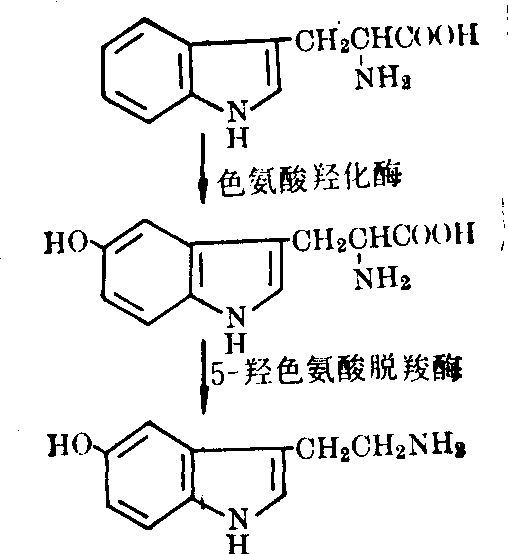
图3 5羟色胺的生物合成
色氨酸主要来源于食物,食物中色氨酸含量明显影响脑中5羟色胺的含量。由于食物的影响,血浆中色氨酸含量有昼夜周期性变化,也反映在脑内5羟色胺含量出现类似的周期性变化。最近提出神经胶质细胞中的cAMP可能参予调节5羟色胺能神经元对色氨酸的摄取及利用。
色氨酸羟化酶: 催化使L色氨酸在第5位碳原子上,加氧羟化生成5羟色氨酸,故又称为色氨酸5单加氧酶。在脑中只有5羟色胺能神经元胞浆中含有此酶。它需要分子氧、还原型蝶啶(四氢蝶啶) 作辅助因子,最适pH为7.2,与色氨酸结合的Km值为3×10-4mol/L,而脑内不同部位色氨酸浓度仅有2.5×10-5mol/L,因此正常脑内色氨酸羟化酶并没有能被作用物饱和,所以增加色氨酸的供给可加速5羟色胺的合成。另外脑内含氧水平也未能使酶饱和,当吸入纯氧,脑内5羟色胺的更新加速。由于上述原因,色氨酸羟化酶反应是5羟色胺生物合成的限速反应。
L5羟色氨酸脱羧酶: 可催化5羟色氨酸脱羧生成5羟色胺,不少实验证明它与芳香族氨基酸脱羧酶(或多巴脱羧酶)可能是同一种酶,但也有人怀疑。它与5羟色氨酸结合的Km值为5×10-6mol/L,活性很高,脑内5羟色氨酸一旦生成,即迅速被脱羧,故脑中5羟色氨酸含量很少。
在中枢神经系统中5羟色胺降解的有效途径是通过单胺氧化酶的脱氨作用,生成的5羟吲哚乙醛能迅即被醛脱氢酶氧化成5羟吲哚醋酸(5HIAA),后者可认为是5羟色胺代谢的终末产物。5羟色胺在体内还可能存在其他降解途径,它的摄取及贮存类似儿茶酚胺。
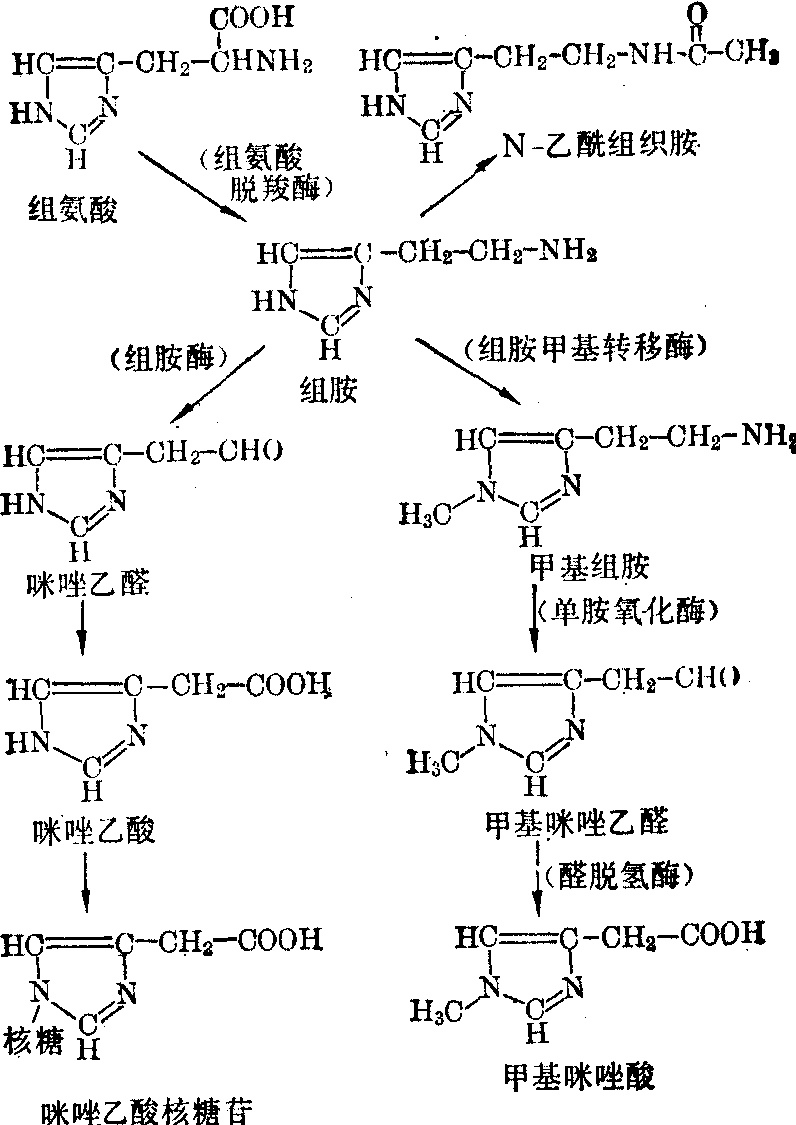
图4 组胺的代谢
组胺 早已知组胺存在于神经组织中,以周围神经及交感神经节后纤维含量最丰富,在中枢神经系统中也含有组胺,哺乳类动物的全脑中约含0.5nmol/L,在不同脑区的分布类似其他生物胺,以下丘脑含量最多,中脑次之,皮质及白质含量最少。在神经元内它主要集中在神经末梢。因观察到.组胺对皮质神经元有抑制效应,周围神经可以释放组胺以及外源性组胺能促进神经节的冲动传导等功能,因此认为它可能也是一种神经递质。
脑中组胺是由组氨酸脱羧形成,而组氨酸则系由血液供给。脑内组氨酸脱羧酶的分布与芳香族氨基酸脱羧酶很相似,可能它们又是同一种酶。但肥大细胞及其他组织尚含有另外一种、最适pH不同的 “特异性”组氨酸脱羧酶。脑中是否也有,尚无证明。组胺在脑内可转变为甲基组胺及甲基咪唑乙酸,其代谢途径如图4。
γ氨基丁酸 在哺乳动物只有中枢神经系统中才发现有γ氨基丁酸 (GABA)。它在脑中分布广,黑质及苍白球含量最多(达9.7mmol/L浓度); 皮质含量较低(约2mmol/L浓度),其他部位介于其间。因对龙蝦牵张感受器有抑制作用,曾称为Ⅰ因子,现已知也是哺乳类动物的中枢神经系统的一种抑制性递质。
γ氨基丁酸是谷氨酸经α脱羧生成。L谷氨酸脱羧酶(GAD)参予催化此反应。给动物注射14C葡萄糖,大部分在脑内转变为谷氨酸及γ氨基丁酸。脑内γ氨基丁酸代谢途径已基本清楚 (图5)。γ氨基丁酸可经γ氨基丁酸转氨酶(GABA_T) 催化,将其氨基转移给α酮戊二酸,生成琥珀酸半醛及谷氨酸。此反应可逆,但在体内实际上并不存在逆向反应。因琥珀酸半醛能迅速被一种脱氢酶(琥珀酸半醛脱氢酶,SSADH) 氧化生成琥珀酸,并进入三羧循环被氧化。这样,γ氨基丁酸的代谢就组成脑内三羧循环的一条重要支路,即GABA旁路(图5),在脑的氧化代谢中有重要意义。
L谷氨酸脱羧酶Ⅰ(GAD_Ⅰ):在哺乳动物也只在中枢神经系统中才有,主要存在于灰质,多分布在突触前神经元末稍中。动物组织中至少有两类谷氨酸脱羧酶。一类分布在脑内,其辅酶为磷酸吡哆醛,即GAD_Ⅰ; 另一类则分布在肾及某些非神经组织中,并不需磷酸吡哆醛,但阴离子能促进其活性,称为GAD-Ⅱ。
γ氨基丁酸转氨酶(GABA_T): 广泛存在于各种组织中,在神经元的核周围、树突以及胶质及内皮细胞中。因此设想GABA是在突触后神经元内及细胞间质部位进行降解或灭活。脑中此酶最适pH为8.2,也需磷酸吡哆醛作辅酶,而且硫氢基可能是酶活性必需的基团。琥珀酸半醛脱氢酶 (SSADH): 能特异地作用于琥珀酸半醛,使之脱氢生成琥珀酸。人脑中此酶的最适pH为9.2,与作用物结合的Km值为5.3×10-6mol/L,所以脑内此酶活性很高,催化的反应需NAD+作为辅酶。
γ氨基丁酸和其他递质一样,是聚集在突触小泡内,它的释放需要Ca2+,释放的γ氨基丁酸可通过γ氨基酸转氨酶降解灭活,但在哺乳动物,突触前末稍的再摄取可能仍是主要的灭活途径。
谷氨酸及天门冬氨酸 L谷氨酸及L天门冬氨酸大量分布在哺乳动物的中枢神经系统中,它们都可在脑内合成 (图5)。因向脊椎动物的脑中离子泳注入谷氨酸或天门冬氨酸,能使神经细胞去极化和增加电发放,故认为它们可能作为中枢神经系统中的兴奋性递质,但仍有争议。它们在脑内的代谢途径见“氨基酸的一般代谢”条及本条中γ氨基丁酸的代谢途径(图5)。
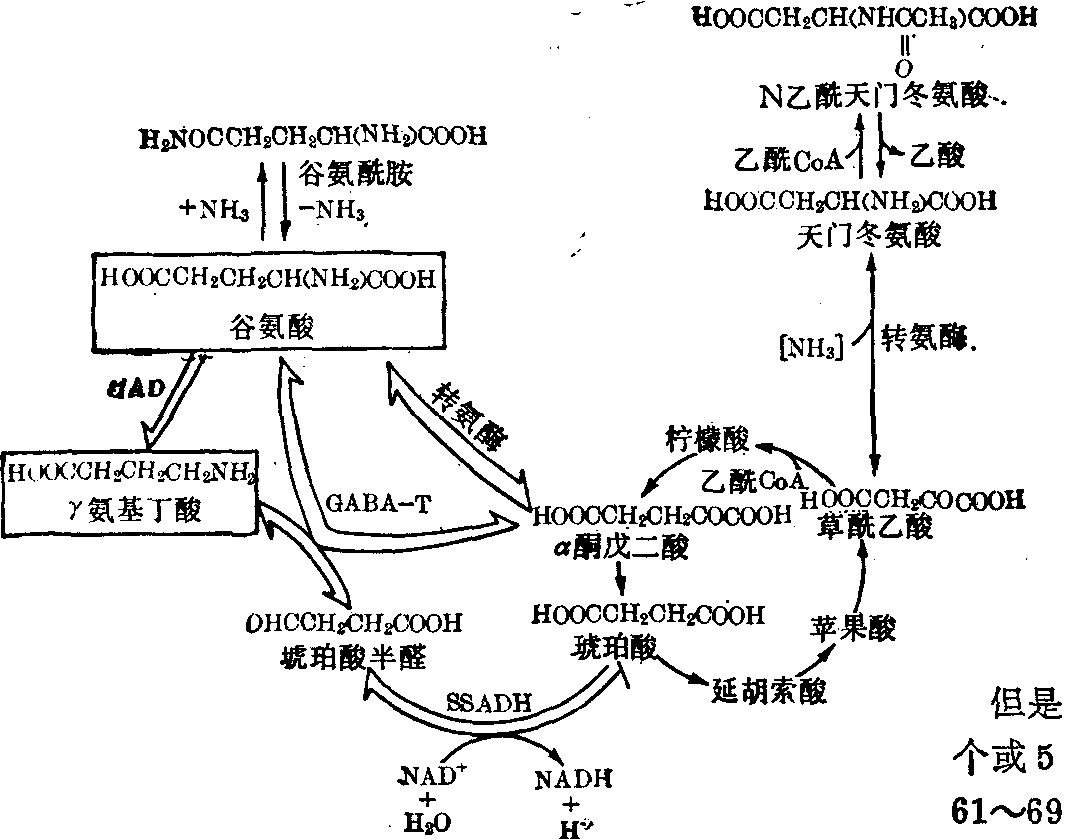
图5 神经系统中γ氨基丁酸、谷氨酸及天门冬氨酸的
主要代谢途径。双线箭头表示γ氨基丁酸代谢的
主要反应(GABA旁路)
甘氨酸 甘氨酸是哺乳类动物脊髓中的一种抑制性递质,主要依据是:
❶甘氨酸在脊髓中含量很高,并集中在灰质中(达4.47μmol/g组织),同时在脊髓前角比在前根纤维中含量高,可能甘氨酸是分布在抑制性中间神经元内;
❷用离子泳技术可证明甘氨酸能降低脊髓运动神经元及中间神经元的电发放及兴奋性;
❸甘氨酸可导致神经细胞膜的超极化及通透性改变。
中枢神经系统中甘氨酸的合成是通过丝氨酸羟甲基移换酶或甘氨酸转氨酶,分别催化下列反应而生成:

脑啡肽和内啡肽 脑啡肽是在1974~1975年由Hughes从猪脑抽提液中分离得到的二种具吗啡 (或鸦片) 受体激动剂作用的活性肽,并确定为二个相似的五肽,其氨基酸顺序如下所示:
H-酪-甘-甘-苯-蛋-OH 蛋脑啡肽
H-酪-甘-甘-苯-亮-OH 亮脑啡肽
二者仅羧基末端不同,一为蛋氨酸称为蛋脑啡肽,另一为亮氨酸,称为亮脑啡肽。亮脑啡肽的活性较弱,只有蛋脑啡肽活性的1/2~1/3。在小鼠、家兔、猪及豚鼠的全脑中,蛋肽啡肽与亮脑啡肽含量的比值约为3.5,在不同脑区的分布与鸦片受体的分布大致平行,但两种脑啡肽的比值则各异。在消化道中也含脑啡肽活性物质,并且是集中在纵肌肠肌丛中。
由于蛋脑啡肽的氨基酸顺序与一种脑下垂体多肽——β脂肪酸释放激素(β-liptropin; β-LPH) 的第61~65位氨基酸顺序相同,因此认为脑啡肽可能是由β脂肪酸释放激素水解断裂生成。人的β脂肪酸释放激素是由91个
氨基酸组成蛋白质,其氨基酸顺序如下:

β脂肪酸释放激素从第61位酪氨酸残基起包括5个以上残基的肽段都有鸦片样活性,如β-LPH,61~79,61~89,61~91等肽段。这类具鸦片样活性的多肽统称为内源性鸦片样肽或内啡肽。
关于脑内鸦片样物质,命名较混乱,有人不主张用内啡肽表示这一组化学物质的总称,而建议使用 “内源性吗啡样物质”一词。因为还可能存在有非肽类鸦片样物质,而Endophins应包括所有这类物质,因此改译为“内啡素”。但本文仍沿用“内啡肽”一词。
分子最大的内啡肽,βLPH61~91,又称为β内啡肽或C肽段,它的镇痛作用最强,并可延续达2小时之久; 而βLPH 61~65 (即蛋脑啡肽)对腺苷酸环化酶的抑制作用较强,而镇痛作用较弱,仅持续15~20 min;其他的活性肽段即βLPH61~65 ……91间各种不同长度的肽段的活性则较为一致。究竟那一肽段是真正生理上有意义的成分,很难确定。不过有人认为短时间作用的肽段(即脑啡肽)可能是神经递质,而长时间作用的肽段可能是神经通路的调节物,二者都有生理意义。
脑啡肽的五肽顺序是内源性吗啡样物质所必需的最小结构,断裂任何一个末端氨基酸残基,活性仅保留1%。酪氨酸、苯丙氨酸的芳香族侧链及疏水的羧基末端都是活性所必需的,尤其是酪氨酸的羟基对活性更是绝对必需。内啡肽如βLPH61~91,镇痛作用时间长,可能只是因分子较大,不易被蛋白酶水解。脑啡肽的空间构型与鸦片结构相似 (即脑啡肽的酪氨酸相当于鸦片分子中酪胺部分),特别是oripavine更为接近,也许这就是二者都能与鸦片受体结合和具有同样生物活性的原因。
P物质 P物质是在1931年从马脑中提取出的一种有降压作用,并能兴奋平滑肌的活性肽; 1970年从牛的下丘脑分离的P物质能使家兔流涎,故又称为催涎肽,并确定为一种十一肽,其氨基酸顺序如下:
精-脯-赖-脯-谷胺-谷胺-苯-苯-甘-亮-蛋-NH2
P物质对初生大鼠的脊椎运动神经元有强烈的兴奋作用,比L谷氨酸的活性强1000~9000倍,其活性能被β(4氯苯基)γ氨基丁酸所拮抗,在中枢神经系统中它还具镇痛作用。P物质高度集中分布在脊髓的背根和背角的背侧部分,并证明是在脊髓神经节中合成,然后运输至背根,贮存在初级传入神经元的轴索末梢中。P物质可能是初级传入神经纤维的兴奋性递质。P物质在不同脑区的分布也不均匀,在人脑中以黑质、下丘脑及松果体含量最高,同一脑区的不同核团P物质含量也有较大的差异。
P物质耐热、抗酸,但可被多种蛋白酶水解而灭活,其羧基末端与活性有密切关系,随羧基末端氨基酸断裂,其活性明显降低或消失。如从氨基末端断裂下某些氨基酸后的十肽、八肽,仍有同样活性,但五肽、四肽或三肽则几乎无活性。
神经递质
神经递质是一种传递神经冲动的化学物质,由神经元末梢所释放,经突触传递至另一神经元或效应器细胞膜上受体,使之发生各种生理效应。其作用非常广泛,可通过效应器调节脏器功能,如调整心率、血压,腺体分泌,肌肉张力,胃肠蠕动和分泌功能,中枢神经的兴奋或抑制等。据释放神经部位可分外周神经递质和中枢神经递质两大类。
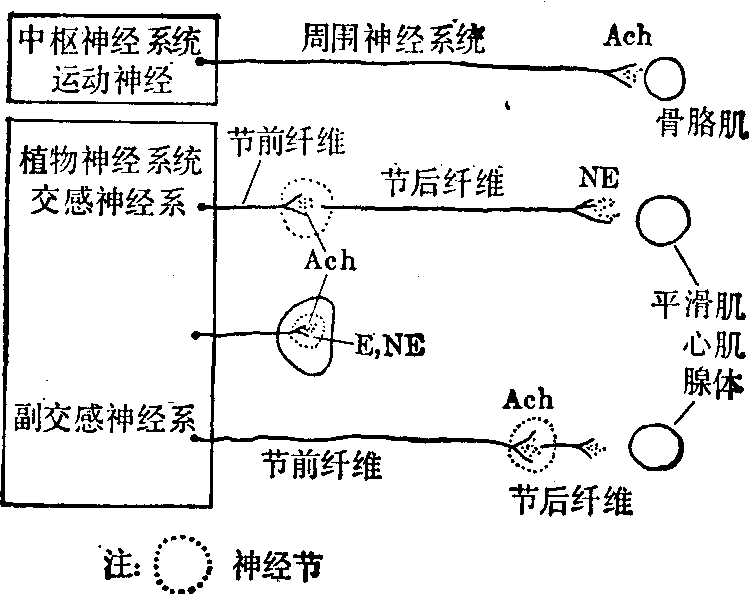
胆碱能神经及肾上腺素能神经分布
神经递质分类表
| 一、外周递质: | 1.季铵类物质(乙酰胆碱)(Ach) 2.儿茶酚胺类:去甲肾上腺素(NE) 多巴胺(DA) |
| 二、中枢递质: | 1.乙酰胆碱(Ach) 2.单胺类: ❶儿茶酚胺类——去甲肾上腺素 (NE) 多巴胺(DA) |
❷5羟色胺(5-HT) ❸组胺(H) 3.氨基酸类: ❶γ-氨基丁酸(GABA) ❷甘氨酸(Gly) ❸谷氨酸(Glu) ❹门冬氨酸(Asp) 4.其他肽类:ATP(核苷酸)、前列腺素(PG) |
外周神经递质 简称外周递质。根据释放的递质不同,可将周围神经中传出神经纤维分成胆碱能神经和肾上腺素能神经两类。胆碱能神经兴奋时其末梢释放乙酰胆碱,它包括:
❶植物神经节前纤维;
❷全部副交感神经节后纤维;
❸小部分交感神经节后维纤(手、足心以外所有汗腺分泌神经、骨胳肌中血管舒张神经等);
❹运动神经。肾上腺素能神经兴奋时,其末梢释放去甲肾上腺素(NE),除大部分汗腺的分泌神经和骨胳肌的血管舒张神经外,交感神经的节后纤维都属肾上腺素能神经。肾上腺髓质受内脏大神经(末梢分泌乙酰胆碱支配,其中嗜铬细胞组织在胚胎中与交感神经都来源于外胚层神经嵴,故也分泌NE与肾上腺素(E)。
心肌、平滑肌(包括胃肠等内脏、血管、虹膜和睫状体肌)、许多腺体都受胆碱能和肾上腺素能神经双重支配,其中之一常占优势而起生理调节作用。
多巴胺为去甲肾上腺素的前体,也是具有生物活性的终末代谢产物,外周组织如肾实质、胰、肾、脑及肠系膜血管与冠状动脉等都有多巴胺能受体,受多巴胺神经元支配。兴奋时DA引起尿钠排出增加,胰分泌增多,肾、脑、肠系膜血管与冠状动脉舒张,血流增多。DA与Ach具有拮抗作用。
中枢神经递质 简称中枢递质。有乙酰胆碱、去甲肾上腺素、多巴胺、5-羟色胺,此四者于下丘脑、边缘系统等处浓度较高,为调节下丘脑释放激素的主要递质,此外尚有几种氨基酸,包括γ-氨基丁酸及组胺等被认为系递质者,详见表。
神经递质neurotransmitter
亦称“神经介质”。神经元之间传递信息的化学物质。在一个神经元与另一个神经元接触的部位(突触),神经冲动的传导叫突触传导。其传导原理是:来自突触前神经元的神经冲动到达突触后,突触前膜内的小泡就会释放出某种化学物质到突触间隙,并对突触后膜发生作用。当这种作用够大时,即可引起突触后神经元发生兴奋或抑制的反应。这两种不同的反应决定于化学递质的性质。例如, 乙酰胆碱能使突触后膜发生去极化,这种电位变化达到一定阈值时,突触后神经元始段就会发生动作电位,并沿着神经纤维传布出去。这是一种兴奋的反应。如果化学递质是多巴胺,就会使突触后膜的电位超过静息电位的数值,加大膜内外的电位差,发生超极化。在超极化状态下,不易去极化, 因而没有动作电位向前传递。这就是抑制性的反应。已确定的和可能的神经递质有:乙酰胆碱、5-羟色胺、多巴胺、去甲肾上腺素、肾上腺素、γ-氨基丁酸、谷氨酸、甲脑啡肽、亮脑啡肽、β内啡肽, 以及前列腺素、P物质等。
神经递质neurotransmitter
简称“递质”。神经细胞间信息或冲动传递时释放的具有生物活性和药理活性的化学物质。分成两种类型: (1) 外周递质,由神经肌接头、外周神经节和支配内脏组织的自主神经末梢释放的递质;(2) 中枢递质,由中枢神经系统所释放的递质。具有充分条件的递质为乙酰胆碱、多巴胺、去甲肾上腺素、5-羟色胺。一些氨基酸行使着递质作用但未完全具备递质的条件。各种递质不仅与动物和人的感觉、知觉、情绪、学习、记忆等心理活动有关,而且与中枢神经系统调节和控制的各种机能活动有关,如睡眠、觉醒、饮水和摄食等活动。
神经递质
简称“递质”。神经细胞间信息或冲动传递时释放的具有生物活性和药理活性的化学物质。分成两种类型: (1) 外周递质,由神经肌接头、外周神经节和支配内脏组织的植物性神经末梢释放的递质;(2) 中枢递质,由中枢神经系统所释放的递质。现在具有充分条件的递质为乙酰胆碱、多巴胺、去甲肾上腺素、5-羟色胺。一些氨基酸行使着递质作用但未完全具备递质的条件。各种递质不仅与动物和人的感觉、知觉、情绪、学习、记忆等心理活动有关,而且与中枢神经系统调节和控制的各种机能活动有关,如睡眠、觉醒、饮水和摄食等活动。
- 字体劲瘦有骨力是什么意思
- 字体很小,笔画纤细是什么意思
- 字体或笔墨雄健挺秀是什么意思
- 字体瘦细而劲健是什么意思
- 字体的笔瘦硬挺拔是什么意思
- 字体的风格是什么意思
- 字体笔势之妙是什么意思
- 字体结构是什么意思
- 字体设计是什么意思
- 字体辨正是什么意思
- 字体飘逸,笔势飞动是什么意思
- 字儿是什么意思
- 字儿话是什么意思
- 字元是什么意思
- 字典是什么意思
- 字典、词典、古代字书是什么意思
- 字典序系统是什么意思
- 字典式目录是什么意思
- 字典排列法是什么意思
- 字典校录是什么意思
- 字典校录外编是什么意思
- 字典正文是什么意思
- 字典眉题是什么意思
- 字典纸是什么意思
- 字典编写法是什么意思
- 字典考证是什么意思
- 字典辞书语体是什么意思
- 字典部首音解是什么意思
- 字典(词典)是什么意思
- 字养是什么意思
- 字写得不好,弯弯曲曲,像蚯蚓和蛇爬行的痕迹是什么意思
- 字写得精彩飞逸,像盘旋往复的喜鹊和鸾鸟一样,潇洒灵动是什么意思
- 字划是什么意思
- 字别辞典是什么意思
- 字势是什么意思
- 字匀是什么意思
- 字匠是什么意思
- 字匯是什么意思
- 字印是什么意思
- 字卷是什么意思
- 字厘南北是什么意思
- 字原是什么意思
- 字原偏旁小说是什么意思
- 字县是什么意思
- 字口是什么意思
- 字句是什么意思
- 字句、内容简明易懂是什么意思
- 字句增损是什么意思
- 字句的错误和脱漏是什么意思
- 字号是什么意思
- 字号儿是什么意思
- 字号店是什么意思
- 字号眼儿是什么意思
- 字号铺是什么意思
- 字同步是什么意思
- 字名是什么意思
- 字向是什么意思
- 字和语素、词的关系是什么意思
- 字唬是什么意思
- 字囝是什么意思