植物生/发芽/长出/植物长/丛生/生长旺盛
☚ 繁衍 植物生 ☛
植物生长growth of plant
植物干重的增加和体积的不可逆增长。生长是植物体内各种生理过程的协调综合,也是植物朝着获得充分成熟体积前进的过程。在生长过程中,植物有序地同化来自环境的无机物质,将它们转化为生命的重要物质,并产生构成植物机体的极为复杂的结构。大多数高等植物的生长依赖分生组织的活动,因此生长局限于发生细胞分裂的分生组织以及附近细胞进行扩伸的区域。生长是产量形成的基础,控制产量必须控制生长。为了了解生长的本质,人们已从数学、解剖学、形态发生和生理学上对生长进行了广泛的研究。生长除决定于遗传因素外,还受环境条件的强烈影响。通过遗传和环境来调控植物生长,是一切农业增产措施的基础。
生长曲线和生长分析 在植物或植物任一器官的生长进程中测定其干重或体积的变化,可以得到一条生长曲线,典型的生长曲线呈S形(见生长周期)。根据生长曲线可分为三个时期,即指数生长期、线性生长期和生长衰减期。在指数生长期中绝对生长速率不断增大; 在线性生长期中绝对生长速率保持恒定; 在生长衰减期中生长逐渐减弱直至停止。由于植物生长过程的复杂和多变,实际的生长曲线与典型的S形曲线常有一定程度的偏离。有时某一个生长期可能消失或特别突出; 有时中间有一段时间生长趋于停顿而形成双S形曲线; 有时曲线平缓; 有时曲线陡峭; 这些变异很大程度上决定于植物的发育情况。
曾有许多植物生理学者试图用数学公式来概括植物的生长规律,20世纪的20~30年代,韦斯特(C.A.West,1920)和格列戈瑞 (F. G. Gregory,1917,1926) 等提出了相对生长速率、叶面积比和净同化率等三个主要参数,用于植物生长的数学分析。
相对生长速率(RGR,R)指增加单位干重所需植
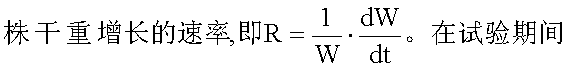
式中 W1为第一次取样时(t1)的植物干重,W2为第二次取样时(t2)的植物干重,ln为自然对数。
叶面积比(LAR,F)指叶面积(L)与整株植物干重之比。即F=L/W。在试验期间的平均叶面积比(
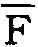 )可用下式表示:
)可用下式表示: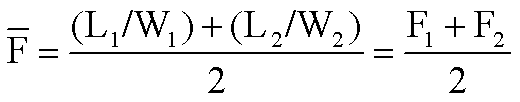 式中 L1和W1分别为第一次取样时的叶面积和整株干重,L2和W2分别为第二次取样时的叶面积和整株干重,F1和F2分别为第一和第二次取样时的叶面积比。
式中 L1和W1分别为第一次取样时的叶面积和整株干重,L2和W2分别为第二次取样时的叶面积和整株干重,F1和F2分别为第一和第二次取样时的叶面积比。净同化率(NAR,E)指增加单位叶面积所需植株干重增长的速率,亦称单位叶速率(UIR),即E=1/L·dW/dt。如在试验期间植物干重与叶面积成线性关系,则试验期内的平均净同化率(E)可用下式表示:
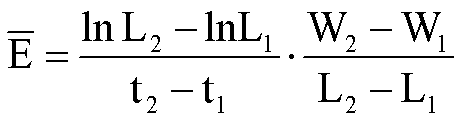 面积比和净同化率三者之间的关系为R=F×E。
面积比和净同化率三者之间的关系为R=F×E。相对生长速率可以作为植物生长能力的指标。从生长曲线来看,相对生长速率在指数生长期最高并保持恒定,之后逐步下降。叶面积比实质上代表植物光合组织与呼吸组织之比,在植物生长早期比值最大。净同化率可以作为光合效率的指标,但不能代表实际的光合作用,因为净同化率是单位叶面积对植株干重净增的贡献,数值因呼吸消耗量的大小而变化。叶面积比随植物年龄增长而下降,即使叶光合活性无变化,净同化率亦将随年龄增长而下降。光线、温度、水分、二氧化碳和无机养分等是影响光合、呼吸和器官生长的环境因子,都能影响相对生长速率、叶面积比和净同化率,因此这些参数可用来分析植物生长对环境条件的反应。生长分析参数值在不同物种间始终存在差异。以相对生长速率来说,低等植物通常高于高等植物;在高等植物中,C4植物高于C3植物,草本植物高于木本植物; 在木本植物中,落叶树高于常绿树,阔叶树高于针叶树。净同化率亦存在类似倾向,但差异小,有时规律还不明显(见表)。沃森 (D.J.Watson)1952年曾指出,决定植物相对生长速率主要是叶面积比而不是净同化率。
细胞水平生长 细胞生长是植物体生长的基础。细胞水平生长过程可明显地分为三个阶段,即细胞分裂、细胞增大和细胞分化。
细胞分裂 受精卵靠细胞分裂生成多细胞个体。细胞分裂本身不造成体积的显著增长,例如在植物顶端分生组织中,分生细胞通常在分裂间隔期内增大,但小量的增长只能接近母细胞的大小。细胞分裂面是决定器官形状的因素之一,在形态建成上有重要作用。如果细胞分裂主要在一个面上大量进行,就形成茎或根之类的伸长器官; 如果在两个面上进行,就形成叶之类的平面器官; 如果在三个或更多的面上进行,就形成果实之类庞大而大致等径的器官。决定器官形状的另一个因素是有差别的细胞增大。一个器官的大小决定于其中细胞的数目 (即器官发育过程中发生的细胞分裂次数)和细胞的体积。植物细胞分裂由吲哚乙酸、细胞分裂素和赤霉素等植物激素促进,由马来酰肼之类的生长抑制剂所抑制。
几种植物RGR和NAR的比较
| 物 种 | RGR (毫克/克·周) | NAR (克/米2·周) |
| 草本 | ||
| 玉米 绿苋 大麦 落叶木本 | 2 310 2 590 920 | 152 147 68 |
| 野苹果 假挪威槭 | — — | 69 33 |
| 欧洲白蜡 常绿木本 | 300 | 28 |
| 酸橙 可可 挪威云杉 | 140 140 58 | 18 13 20 |
细胞增大 从分生细胞发育成薄壁细胞,在体积上有惊人的增加。例如皮层薄壁细胞或叶片栅栏细胞比产生它们的立方形分生细胞宽5倍,长20倍,体积增加500倍。由于细胞长度较宽度增加大得多,细胞增大常指细胞的伸长。有些大致等径的细胞(如髓薄壁细胞)是三维的同时增长。植物细胞增大的先决条件是细胞壁的可塑化,这需要有适当浓度的生长素存在。细胞吸水膨胀后将细胞壁推出导致细胞增大。由于细胞扩大,压力势减小,水势因之降低而使更多的水分进入细胞。大量水分的进入,使液泡扩大和合并成一个,占据细胞的大部分体积,原生质成为靠着细胞壁的薄薄一层。细胞增大还包括大量纤维素、果胶化合物和其他细胞壁物质的掺入,使细胞壁显著增厚。原生质虽只占一薄层,但实际体积和在分生细胞中相比亦大为增加。所以细胞增大包含了相当量的同化作用。叶绿体等细胞器的发育和分化也在这个时期进行。
细胞分化 细胞分化常指纤维、导管分子、筛管分子、木栓细胞、厚角细胞和保卫细胞等一些特化细胞的发育。分化过程包括次生细胞壁的沉淀,细胞形状的变化,角质、木栓等物质的合成和沉积,筛管分子中核和液泡膜的丧失,导管分子和纤维中原生质的丧失,导管分子壁端的溶解,以及细胞器种类、数目和大小的改变。在发生这些变化之前,细胞一般也要经过一个类薄壁期,这是原始分生细胞增大的结果。一旦开始分化,体积不再增长,所以通常不将细胞分化归入生长范畴。但是薄壁组织的细胞增大过程中也存在大量分化,使薄壁组织包含了在形态、构造和生理上有显著差异的各类细胞,在细胞分化过程中也发生着干重的增加,因此分化很难与生长截然分开。
营养生长和生殖生长 种子萌发后,先进行营养生长,形成根、茎、叶等营养器官。而后才开始生殖生长,进行花芽分化,形成花、果实、种子等生殖器官。茎和根的延长生长靠顶端分生组织的细胞分裂活动而实现。茎尖的顶端分生组织使茎轴延长,并形成新叶,产生腋芽。腋芽生长形成侧枝。根在延长过程中亦不断发生分枝,形成侧根。种子植物的侧根多起源于根毛区的中柱鞘,一定部位的中柱鞘细胞恢复分裂能力成为一新生长点,经过细胞分裂、伸长和分化,形成根的原始体,最后突破皮层和表皮形成侧根。裸子植物和双子叶植物的茎和根能依靠侧生分生组织形成层的活动而不断增粗。在禾本科植物茎秆的生长过程中,顶端分生组织的一些有分生能力的细胞能保留在节间基部,成为居间分生组织。居间分生组织活动引起的居间生长,是禾本科植物在拔节期间茎秆快速伸长的主要原因。叶起源于茎尖生长锥周围的叶原基。叶原基先进行先端生长,使叶迅速伸长; 到一定时间,先端生长停止,转为基部的居间生长,居间生长可维持较长时间。
植物经过一定时间的营养生长后,茎部顶端分生组织开始形成花或花序,出现生殖生长。种子植物的生殖生长过程主要包括花原体的发端、花器的成熟、花药中花粉粒的发育、子房各胚珠中胚囊的发育、授粉、花粉粒或花粉管中生殖核形成两个精子、花粉管从柱头长入胚珠、受精(一精子与卵并合)和三核并合(一精子与并合核并合)、受精卵发育成胚、胚乳核发育为胚乳、胚珠发育成种子以及子房及相邻组织发育为果实。这些生殖生长过程可分为两个不同阶段,即开花阶段和结实阶段。开花标志植物进入了一个新的发育阶段,有复杂的质变内涵; 结实则与营养生长性质相似。
营养生长与生殖生长间有复杂的相互关系。❶生殖生长需要以营养生长为基础。植物开花前必须经历的营养生长时期称为花前成熟期或幼年期,草本植物一般为数周到数月,木本植物长的可达30~40年。
❷营养生长和生殖生长相互制约。一次结实植物生殖器官的出现就是营养生长的全面结束; 多次结实植物虽然营养生长可与生殖生长并存,但在生殖生长期间营养生长明显减弱。摘除花芽可以维持某些植物的营养生长; 栽培烟草时进行打顶可促进烟叶生长。过分旺盛的营养生长亦能影响生殖生长,徒长的植株通常结实不良。一些抑制营养生长的措施,如修剪或施用生长抑制剂,能促进某些植物的成花。这种相互制约关系可用养分分配理论来解释,活跃生长的枝梢和发育期中的果实都能成为强有力的养分“壑”(Sink),控制植物体内光合产物的流向。植物激素对养分壑的建立有重要作用。
生长和遗传潜势 任何植物的生长都决定于遗传潜势和适于遗传潜势表达的环境。由于这两方面的差异,使不同植物的器官和组织的生长千差万别。遗传潜势决定于植物的基因型,基因是脱氧核糖核酸(DNA)分子中特定核苷酸序列提供的密码信息。活化态的基因能将DNA密码转录为具互补密码的信使核糖核酸(mRNA),mRNA密码在转移核糖核酸和核蛋白体的协同下,转译为有特定氨基酸序列的蛋白质分子。遗传潜势的运转决定了植物体所能合成的酶 (及其他蛋白质),这些酶决定了植物体所能产生的特定生化反应,即决定了植物的生长类型。植物的每个体细胞通常都含有相同的基因组(即相同的遗传潜势),但并非所有基因都处于活化态。在植物的不同部位和不同发育阶段,细胞内具有不同的活化态基因,产生不同的mRNA,因此细胞具有不同的酶系统和不同的生化能力。如果细胞内已具备了特定的mRNA,还可以在转译阶段抑制酶的合成。已存在于细胞内的酶,还可以被代谢反馈机制所钝化。
在真核生物中,细胞在特定时期活化特定基因的控制机理尚不清楚。由于大部分DNA与组蛋白形成可逆性复合物,组蛋白能通过改变染色体的构型来调控基因的活性,与组蛋白复合的DNA处于超卷曲状态,无法与RNA聚合酶接触,因此失去了转录的能力。有许多效应物通过组蛋白对核蛋白的作用能影响基因的活性,激素就是一种效应物。基因的抑制和活化是激素在生物体内发挥影响的途径之一。发芽种子胚中产生的赤霉素诱导胚乳中α-淀粉酶的合成,显然这种植物激素能活化特定的基因,使编码这种酶的mRNA得以转录产生。环境因子通过影响激素的合成,亦能在基因的活化或抑制中发挥作用。
生长和环境因素 任何代谢过程的进行都要求一定的环境条件,不满足这些条件,即使细胞中含有代谢所需的全部酶类,代谢亦无法进行。代谢的速率常被供应量最低的环境因子(限制因子)所决定。某些环境因子(特别是光和温度) 还有显著的形态建成效应。环境因子包括生物因子、物理因子和化学因子。
生物因子 两个不同物种的个体间的任何生态互作,能影响两者的生活过程和改变它们的正常生长型式。例如在寄生情况下,寄生物有时能杀死寄主或抑制寄主的生长,有时能引起寄主不正常的生长进而形成瘤瘿。在共生情况下,则共生成分的生长均受到促进。生物体也可能通过改变生态环境来影响另一生物体。如林木影响下木的生长; 某些高等植物生产抗生物质来抑制其他高等植物的生长。
物理因子 包括能量的各种不同表现形式。如热、光、声、重力和动能等。温度影响与代谢有关的所有生物物理和生物化学反应的速率。温度对植物生长有时间效应,即生长最适温随着持续时间的延长而下降。这主要由于酶在高温下的钝化现象,温度超过40℃,酶开始钝化; 到60℃,大多数酶就变性。植物生长的最适温,黑夜与白昼有所不同,这种现象称为温周期。一般植物在夜温低于昼温的条件下生长最好。生长最适温还随植物的发育期而变化。这可看作是年或季节温周期。温度对生长还有诱导效应,即短暂的温度处理能引起持久的效应,如低温对某些植物芽和种子的破眠作用。
光强、光质和光周期三者对植物生长都有深远影响。光合作用和光形态建成是两个最重要的受光控制的生理过程。光合作用中通过光引起的光化学反应,将光能转变为化学能,还原无机碳化物为有机碳化物,构成植物生长的基础。光形态建成是低强度光对植物形态和结构的调节效应。从广义来说,凡受光调节和控制的一切非光合效应,如光周期、向光性、趋光性和脱黄化作用等,都属于光形态建成范围。从狭义来说,仅指光与种子萌芽、茎叶生长、开花成熟、细胞器形成等方面的关系。
超声波能杀死微生物,强度大时对高等植物生长亦有不利效应。声波(包括音乐)对植物生长的影响尚无定论。重力影响生长可从植物的向地性得到证实。有些植物(如糖槭)叶的大小和形状,以及枝条上下侧的次生生长量亦受重力的影响。影响植物生长的动能类型是大气流和水流。大气流主要通过它对光合和蒸腾的效应而影响植物生长; 水流主要通过改变水生植物表面氧、二氧化碳和无机盐的浓度而影响水生植物的生长。
化学因子 包括各种化学物质,如水分、大气和矿质等。植物生长对水分供应状况最为敏感,细胞分裂和细胞增大都受水分亏缺的抑制。特别是细胞增大时相对含水量稍低于饱和就不能进行。供水不足的植物,体积增长提早停止,结构分化较快发生。水分亏缺亦会影响光合作用,缺水对光合的抑制主要是通过水对气孔运动的效应而不是水作为光合反应物的作用。
大气成分中对植物生长影响最大的是氧、二氧化碳和水气。氧为一切需氧生物生长所必需,大气含氧量相当稳定,植物的地上部分通常无缺氧之虞,但土壤在过分板结或含水太多时,常因不能供应足够的氧气而使根部生长不良甚至坏死。大气中二氧化碳含量很低,常成为光合作用的限制因子。人为提高空气中的二氧化碳浓度,常能显著促进植物生长。在通气不良的土壤中,因根部呼吸引起的二氧化碳大量积聚,不利于根部生长。大气中水气含量变动很大。水气含量通过影响蒸腾作用而改变植物的水分状况,从而影响植物生长。由人类经济活动产生的许多大气污染物对植物生长有显著影响,使植物生长减弱,出现受害症状,甚至整株死亡。
土壤中植物必需的矿质元素,目前已知有氮、硫、磷、钾、钙、镁、铁、锰、锌、钼、铜、氯、硼等13种,这些元素有些属于原生质的基本组分; 有些是酶的组分或活化剂; 有些能调节原生质膜透性,参与细胞缓冲体系以及维持细胞的渗透势。植物缺乏这些元素便会引起生理失调,影响生长发育,并出现特定的缺素症状。
- 旧金山会议是什么意思
- 旧金山和约是什么意思
- 旧阮是什么意思
- 早产是什么意思
- 早产是什么意思
- 早产是什么意思
- 早产是什么意思
- 早产儿和小于胎龄儿是什么意思
- 早产儿的特点和护理是什么意思
- 早作是什么意思
- 早发是什么意思
- 早发白帝城(1)是什么意思
- 早发竹下(1)是什么意思
- 早川一荣是什么意思
- 早强剂是什么意思
- 早拆模板体系是什么意思
- 早搏是什么意思
- 早晚用收敛性化妆水是什么意思
- 早晴至报恩山寺是什么意思
- 早期(Ⅰ、Ⅱ期)乳腺癌的治疗及疗效是什么意思
- 早期妊娠人工流产法是什么意思
- 早期智人是什么意思
- 早期港口建设是什么意思
- 早期码头和货仓建设是什么意思
- 早期胚胎外形的主要变化是什么意思
- 早期航运事业是什么意思
- 早期茶馆有什么类型是什么意思
- 早期蒙古社会性质是什么意思
- 早期认识是什么意思
- 早期诊断是什么意思
- 早期防治老年眼病是什么意思
- 早泄是什么意思
- 早熟型消费是什么意思
- 早熟型消费是什么意思
- 早熟型消费是什么意思
- 早熟性是什么意思
- 早熟栽培的技术措施是什么意思
- 早熟栽培葡萄成熟的时期和采收标准是什么意思
- 早熟栽培葡萄的包装和运输是什么意思
- 早熟栽培葡萄采收后的分级是什么意思
- 早熟禾品种鉴定方法是什么意思
- 早熟禾属的分类和形态特征是什么意思
- 早熟禾属苯酚鉴定法是什么意思
- 早熟葡萄品种是什么意思
- 早熟鲜食葡萄未来的市场前景是什么意思
- 早玛瑙是什么意思
- 早玫瑰是什么意思
- 早生高墨是什么意思
- 早老症是什么意思
- 早艳是什么意思
- 早行是什么意思
- 早行是什么意思
- 早起见雪是什么意思
- 旬邑县是什么意思
- 旬阳县是什么意思
- 旭日牌牛黄清心片是什么意思
- 旱伞草是什么意思
- 旱伞草是什么意思
- 旱地栽培施肥技术是什么意思
- 旱草清是什么意思