染色质chromatin
真核细胞间期核内DNA、组蛋白、非组蛋白以及少量RNA组成的纤维状复合物,是遗传物质的一种存在形式。1882年弗勒明(W.Flemming)提出的染色质概念是指细胞核内能被碱性染料着色的物质,以及分裂期间构成染色体的物质,是形态学术语。染色质和染色体的组成成分相同,但构像各异,两者相间出现在细胞周期中的不同阶段。
化学组成 染色质中的DNA、组蛋白、非组蛋白、RNA大约为1:1:0.1~1.0:0.05,此外还有微量的多胺类化合物。DNA构成染色质的骨架成分,其空间构像决定染色质的功能,而核苷酸序列决定遗传信息的本质。真核细胞中许多DNA序列重复存在,依照序列相同的片段在基因组中出现的次数可分为高度重复序列(又称丰富基因)、中度重复序列和单一DNA序列,其功能各不相同。绝大部分DNA与组蛋白相结合存在,大约只有3~20%的DNA呈游离状态。组蛋白是富含赖氨酸、组氨酸等碱性氨基酸的碱性蛋白质,可分为H1、H2A、H2B、H3和H45类。除精子中组蛋白被鱼精蛋白取代、鱼类和鸟类细胞中H1被H5取代等极少数例外,所有真核细胞中都存在上述5类组蛋白。编码组蛋白的基因属丰富基因,在细胞内为多拷贝,其表达在S期与DNA复制偶联进行。目前认为组蛋白有稳定染色质结构和使基因关闭的作用。非组蛋白是富含天门冬氨酸、谷氨酸等酸性氨基酸的蛋白质,种类繁多,且有生物种属和器官的专一性,也可因细胞的生理状况而变化。非组蛋白的确切功能尚不清楚,一般认为它有解除组蛋白对基因的束缚作用,使基因得以表达。某些RNA分子可与DNA一定部位相结合,构成染色质的成分,它们在细胞分裂时结合到DNA上,细胞处于间期时又回到核液中,推测这些RNA可能与染色质的高次结构形成有关,对基因的转录活性有控制作用。染色质中的多胺类化合物主要包括精液酸、精胞胺、腐胺和尸胺等,它们是鸟氨酸或精氨酸的脱羧产物,结合于DNA双螺旋的小沟内,能稳定或改变DNA的构像。
基本结构单位 1974年奥廉恩(D.E.Olins) 等将鸡红细胞等材料经低渗处理破裂后,用四氧化锇固定,醋酸铀染色,电镜观察发现,9nm左右的球状颗粒沿一条直径为2~3nm的纤维有规律地排列,呈念珠状结构,认为此球状颗粒是染色质的基本结构单位,并命名为ν-体。稍后,钱博恩(Chambon)也证实了这种颗粒状结构的存在,并主张称核小体。此名词沿用至今。1980年英国的弗拉格(A.Flug)小组得到了球状颗粒的结晶,经X衍射和电镜观察后,提出核小体为11×11×5.7nm的结构,140个碱基对绕其上1.75周,旋距2.8nm。现一般认为核小体由核心颗粒和桥两部分组成,核心颗粒由H3、H4、H2A、H2B各两个分子形成一个八聚体,140个碱基对的DNA在八聚体上绕1.75周。两个八聚体之间有60个碱基对的DNA相连,其上结合有H1,这部分称为桥。八聚体中组蛋白分子的结合方式,根据彼此之间相互作用的强弱有过不少模型。较多人认为H3、H4位于八聚体的中心,H2A、H2B结合其外。绕在八聚体上的DNA长度和桥部分DNA长度共200个碱基对,但亦非尽然,出现差异的部位是桥部分的DNA长度。关于DNA进行复制和转录时,核小体的结构状态如何,有过不少探索,目前尚无明确的看法(图1)。

图 1 核小体结构模式图
类型 细胞学根据染色性能不同将染色质分为常染色质和异染色质。常染色质是指间期核的染色质上和分裂期的染色体上被碱性染料染色较浅的区域,染色较深的区域称为异染色质。异染色质中又分为可变异染色质和结构异染色质。前者可以随细胞类型和发育阶段不同而变成常染色质,后者是指所有细胞类型和发育阶段深染的部分,主要存在于着丝点、核仁组织区等部位。染色体臂的不同部位也有不同量的结构异染色质插入其中。从物理特性和生化角度来看,常染色质分散程度高、纤维结构盘曲度大,可发生乙酰化和进行转录,在S期的早期进行复制,而异染色质则与此相反。
1976年赖斯(H.Ris)按功能将染色质分为活动性染色质和非活动性染色质。活动性染色质是指有转录功能的部分,大部分在常染色质区,螺旋化程度低,比较松散; 非活动性染色质螺旋化程度高,不能进行转录。两者还有一些特性上的差异(见表)。
活动性染色质和非活动性染色质特性的比较
| 特 性 | 活动性染色质 | 非活动性染色质 |
| 热变性温度 在蔗糖梯度中的沉降速度 随体DNA 活化RNA聚合酶的能力 与新生RNA的连接 与同源RNA的杂交能力 | 较 低 较 慢 无 有 有 有 | 较 高 较 快 非常丰富 无 无 无 |
染色体组装 一条染色体中的DNA长度约为中期染色体长度的1×104倍。因此,由染色质变成染色体必需经过复杂的组装过程:DNA双螺旋分子直径约为2nm,它与组蛋白结合形成核小体时,成为11nm左右的纤维状结构,这是染色质的一级结构。从一级结构变成30nm左右的纤维状二级结构可能有两种情况,一是由每六个核小体盘旋一周形成外径30nm、内径10nm、旋距11nm中空螺旋管;另一种则可能是相邻核小体交错排列成两列,形成粗约30nm双列核小体结构。从一级结构到二级结构的变化过程中H1可能起一定作用。H1分子的中央为球状结构,两端有处于伸展状态的带氨基末端和羧基末端的两个臂,一旦H1的中央球状部分与核小体的特定部位结合,则伸展状态的两臂中的一臂在桥连接DNA和核小体八聚体结合部位附近盖住连接的DNA,另一臂与相邻的另一个八聚体连接使核小体之间彼此相连,通过H1使核小体集合起来形成一个规则的重复结构。从二级结构到三级结构乃至中期染色体的形成过程有不同的解释。一种认为30nm左右的螺线管进一步螺旋化,形成直径约400nm的圆筒状的超螺旋体,超螺旋体进一步折叠和螺旋化变成中期染色体;另一种看法是30nm的纤维成分通过折叠形成直径300nm的纤维,再经螺旋成为直径700nm的纤维结构,由此进一步凝缩成直径为1 400nm的中期染色体。实际上由染色质组装成染色体的过程可能不只是一种方式,但最后都压缩到DNA长度的万分之一(图2)。
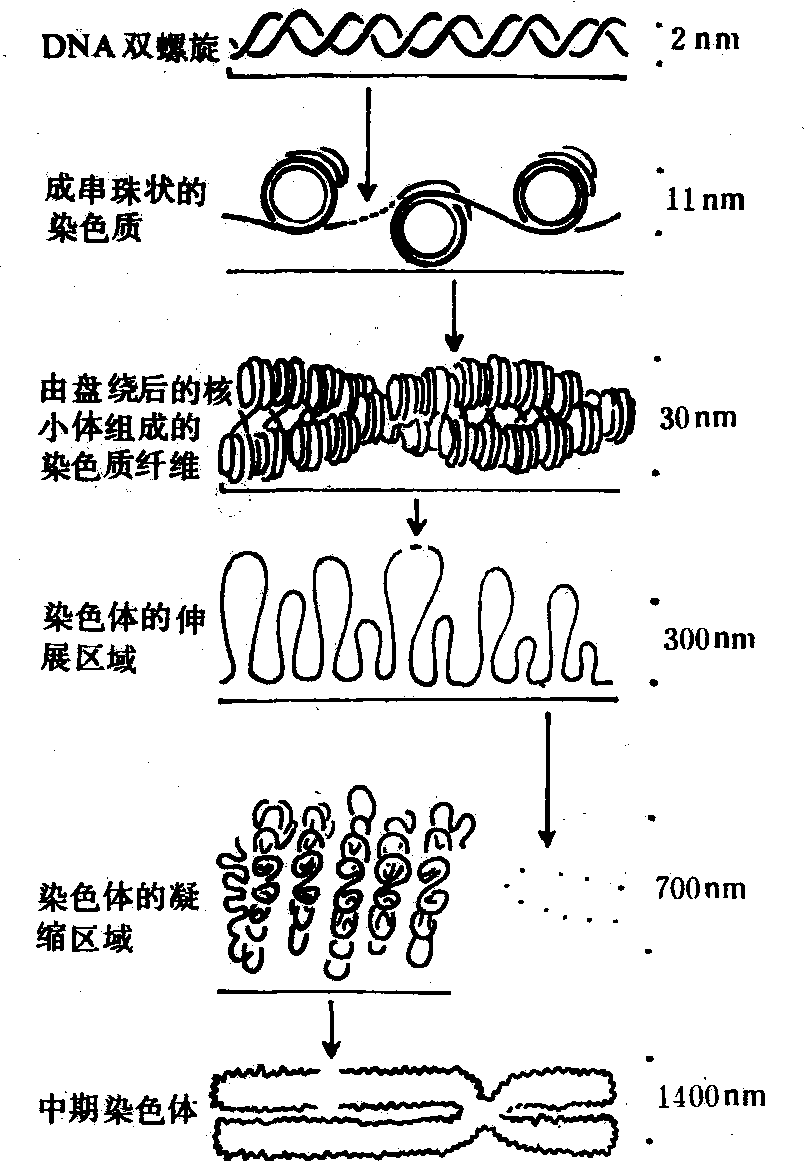
图 2 染色体组装示意图
非组蛋白骨架 非组蛋白在染色质中除了可能作为一种调节基因活性的物质外,其中有一部分还可能参与染色质的高次结构,与中期染色体的形成有关。实验表明,用各种方法除去海拉(Hela)细胞中期染色体的组蛋白和非组蛋白的大部,其DNA与蛋白质为6:1,仍然保持中期染色体的形态。电泳分析,其中含有30种非组蛋白。主要有3种,分子量分别为5.3×104、5.1×104、3.5×104,表明非组蛋白在中期染色体中使DNA保持高度折叠的构型。后来有人用电镜进一步观察这种没有组蛋白的染色体,也证明它是由一种骨架成分构成,具有中期染色体的外部形态特征,但周围被宽阔的DNA晕轮包围。晕轮是由许多长为10~30μm的DNA环构成的,每个环的基部都固定在骨架内。这些结果表明,中期染色体的基本形态是由非组蛋白骨架决定,非组蛋白在染色质的高次结构中有重要作用。
染色质chromatin
真核细胞间期核内由DNA、组蛋白、非组蛋白及少量RNA组成的细丝状复合物。易被碱性染料着色,故名。DNA链构成染色质的基本骨架,带有由不同核苷酸碱基序列编码的遗传信息;组蛋白是含有碱性氨基酸的碱性蛋白质,可分为H1,H2a ,H2b,H3和H45种,有稳定染色质结构和抑制基因表达的功能;非组蛋白为酸性蛋白,种类繁多,具有物种和组织特异性,有专一性控制DNA表达和参与形成染色质高层次结构的作用。染色质细丝的基本结构由DNA链盘绕组蛋白形成核小体,核小体间有较短的DNA链相连,许多核小体重复排列形成直径10 nm串珠状的染色质丝,并可进一步螺旋化使DNA链进一步卷曲、折叠。根据螺旋化程度可分为异染色质和常染色质两类。
染色质ransezhi
间期细胞核中可被碱性染料染成深色的物质。其中,呈伸展状态的区段称为常染色质;呈卷曲凝缩状态的称为异染色质。
染色质由脱氧核糖核蛋白组成,它是DNA、组蛋白、非组蛋白和少量RNA的复合物。不同染色质中,RNA和非组蛋白的含量不同,但是DNA与组蛋白的含量比例均保持为1:1。电镜下观察,间期核中染色质呈串珠状细丝,每条染色质丝的直径约为10毫微米,它是由核小体连接而成的。核小体是以组蛋白为核心的DNA分子的缠绕部分,许多核小体之间的连接线则是DNA分子的延伸部分。每条染色质丝可进一步卷曲成直径为30毫微米的螺旋管,称为染色质纤维,螺旋管的每一圈由6~8个核小体围成。在细胞有丝分裂的前期,染色质纤维卷曲成染色体,到分裂中期时高度凝缩成有一定结构形态的中期染色体。到分裂末期时染色体逐渐解螺旋化,复原成间期状态的染色质丝。可见,染色质和染色体是细胞间期中不同阶段的两种存在形态。
染色质chromatin
系DNA和组蛋白的复合体。与染色体实为同一种物质。只是染色质出现在细胞的分裂间期,在光学显微镜下观察为大小不等的颗粒,不均匀地分布在细胞核中。在电子显微镜下观察为念珠状细丝,有的呈松散的缠绕状态,有的呈伸展状态。染色质在细胞分裂期逐渐螺旋卷曲成染色体。其功能见染色体条。
染色质
真核细胞的核中,由DNA、碱性蛋白(组蛋白)、非碱性蛋白以及少量RNA构成的复合体。细胞间期时,基本形态是由染色质丝组成的网状组织;在分裂期固缩成染色体。
染色质chromatin
指细胞核内容易被碱性染料染色的化学物质。其主要化学成分是DNA、组蛋白、非组蛋白和少量RNA。其细微结构是由组蛋白形成的八聚体,外缠绕13/4圈DNA分子链所形成的核小体,许多核小体连接的念珠状结构即染色质。它是染色体的前体物质。当细胞分裂时,染色质螺旋化、折叠,形成染色体。
染色质
染色质是间期细胞核的重要组成成分,主要由DNA和组蛋白构成,易被碱性染料染色,间期核的染色质在光学显微镜下呈丝状、颗粒状或斑块状,在电子显微镜下则可辨别出异染色质和常染色质。异染色质电子密度较高,呈斑块状,通常分布于内核膜的内侧。常染色质电子密度低,位于异染色质之间的浅亮区域。染色质在细胞有丝分裂时期,浓缩成具有一定形态结构的染色体,染色质与染色体的组成成分相同,只是由于细胞所处生活周期和功能状态不同,而呈现的不同形态结构。
染色质的化学组成 染色质的化学成分主要是DNA和组蛋白,另外还有非组蛋白和少量RNA,组蛋白与DNA的含量大致相等,非组蛋白的含量变化很大。
染色质中的DNA是遗传的主要物质基础。每种生物细胞核中的DNA含量都是稳定的,例如大白鼠的体细胞核内含有6.5×10-2gDNA,人体的体细胞核内含有5.6×10-12gDNA。DNA是由两条互相缠绕着多核苷酸链组成的双螺旋分子。多核苷酸则由多个单核苷酸排列而成,每个单核苷酸又由磷酸、脱氧核糖和四种碱基(腺嘌呤、鸟嘌呤、胸腺嘧啶及胞嘧啶)组成。遗传信息蕴藏在DNA分子的许多碱基排列的顺序之中,当碱基顺序发生变化时,遗传信息也随之改变。
有核细胞的染色质中含有五种组蛋白,即H1、H2A、H2B、H3和H4。这五种组蛋白又可根据其赖氨酸和精氨酸的相对含量而分为三类,即富含赖氨酸的H1,稍富含赖氨酸的H2A和H2B以及富含精氨酸的H3和H4。组蛋白为碱性蛋白,能以离子键与DNA紧密结合。组蛋白具有自动聚集的趋势。一般情况下,H4和H3、H2△和H2B相集合成对,并构成核体的核心颗粒。H1则不与任何其他组蛋白相集合。各种生物细胞核中的组蛋白类型与含量没有明显差别。一般认为组蛋白与DNA功能的调节有密切关系,它能抑制DNA的转录活动。
染色质非组蛋白,又称酸性蛋白。因为,在大多数非组蛋白,其酸性氨基酸(谷氨酸、门冬氨酸)含量比碱性氨基酸(精氨酸、赖氨酸和组氨酸)高。染色质非组蛋白含量变化很大,种类也很多。非组蛋白中有许多为酶蛋白,如与DNA合成有关的DNA聚合酶及DNA连接酶,与RNA合成有关的RNA聚合酶,以及与蛋白质加工,降解有关的蛋白质酶等。除了酶蛋白以外,在非组蛋白中还有一些结构蛋白和少量基因调控蛋白。基因调控蛋白在控制基因表达上起着重要作用,认为它能活化基因组中被组蛋白掩蔽的区域,而使DNA转录RNA。
染色质的微细结构 近年来应用电子显微镜铺展技术,核酸酶消化和X线衍射等方法,对染色质的微细结构进行了研究。1973年有人应用电子显微镜直接观察到,处于细胞分裂间期的染色质是由彼此相隔一定距离的珠状颗粒组成的串珠状结构。珠状颗粒被称为核小体,是染色质有序的重复亚单位。核小体由核心颗粒和颗粒间的连接丝构成。核心颗粒为八聚体组蛋白,其表面盘绕着DNA。因之,染色质中DNA与组蛋白的排列关系不是以前所认为的组蛋白覆盖在DNA分子之上,而是DNA包绕着组蛋白。
近几年来的研究工作发现,核小体是11nm×11nm×5.7nm大小的扁圆形球体,其核心颗粒包括由各两个分子的H2A、H2B、H3和H4组成的八聚体组蛋白核心和包绕在核心外围,链长为140个碱基对的DNA;而其连接丝则包括组蛋白H1和链长60个碱基对的DNA组成八聚体的四种组蛋白近似等量,彼此之间具有很大的亲和性。四种组蛋白的排列方式,可能是H3和H4各两个分子组成四聚体,两个H2A和H2B分子附着其中。核小体DNA包括核心颗粒DNA和连接丝DNA,最常见的长度为200个碱基对。其中140个碱基对在八聚体表面左向超微螺旋1.75圈,间距为2.8nm,旋转直径为11nm;其余的60个碱基对组成连接丝。各种细胞中的核体连接丝的DNA长度并不相同,介于20—100个碱基对之间。这样便造成核小体的DNA长度也不一样,一般具有160—240个碱基对。目前认为组蛋白和DNA可由组蛋白多肽链上碱性氨基酸所带正电荷和DNA分子磷酸上所带电荷之间的静电吸引力而互相结合(图1、2)。
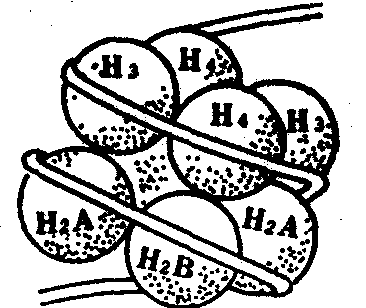
图1 四种组蛋白构成八聚体
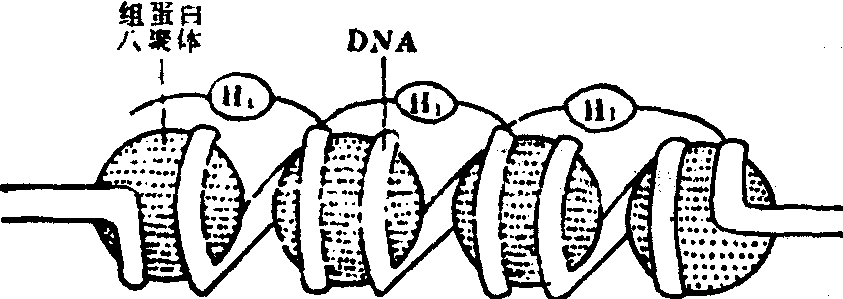
图2 核小体结构示意图
1976年在电子显微镜下看到间期核中有直径30nm的染色质纤维,称为染色质粗纤维。染色质粗纤维是由核小体进一步螺旋化而形成的外径为30nm、 内径为10nm的中空管状结构,称为螺线管。螺线管的螺旋间距为30nm,每一螺旋含有6个核小体。组蛋白H1对螺线管结构的形成有一定作用。H1不仅能和DNA有力地结合而交联核小体,而且能使核小体螺旋化,导致高级螺旋结构——螺线管的形成。
在细胞分裂前期中,螺线管可再次盘绕成为更高一级水平的染色质结构,称为超级螺线管。超级螺线管直径为40nm,超级螺线管再经卷曲、折叠便形成染色体。
在染色质由其初级结构——核小体经螺线管、超级螺线管而卷曲、折叠成染色体的过程中,通常长达数厘米的一条DNA,由于螺旋化,而被压缩近万倍。例如,人体细胞中的1.4—7.3 cm长的染色质DNA可被紧紧地包装在长2—10μm的染色体中。一般认为DNA的长度被压缩的具体倍率是:在核小体内为7倍,在螺线管内为6倍,在超级螺线管内为40倍,最后在染色体内为5倍,总共被压缩8400倍(7×6×40×5)。显然,这种结构方式有利于将大量的遗传信息储存于有限的空间之内(图3)。
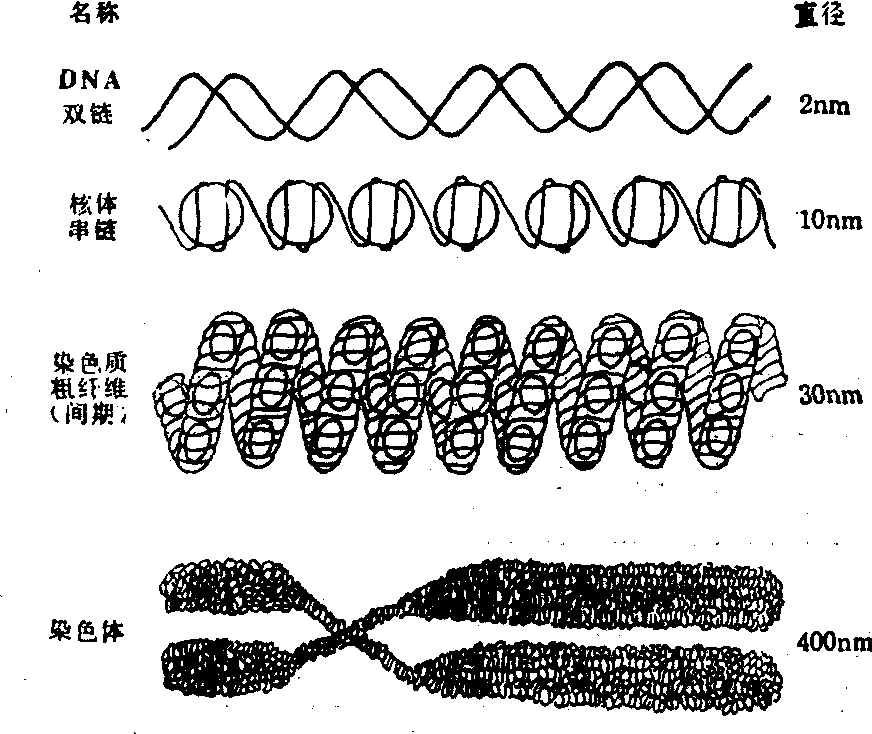
图3 染色质各级结构示意图
染色质的功能 染色质的主要功能是复制DNA和转录RNA。处于功能状态的染色质,螺旋化程度低,结构松弛,即所谓常染色质,或称伸展染色质。功能不活跃的染色质,即异染色质,由于其螺旋化程度高,结构紧缩,也称浓缩染色质。间期核内的上述两种不同形状表现的染色质,可随细胞的功能状态而相互转变。例如,小淋巴细胞的核富于异染色质,但在植物性血细胞凝集素的作用下,大部分异染色质可转变为常染色质,并大量地转录RNA。
真核细胞染色质DNA所含碱基对中,只有1—3%参加,RNA的转录,其余的则负责调节功能,或做为染色质的结构成分。DNA以其双链中的一条为模板进行转录。DNA转录rRNA、mRNA和tRNA时,需分别有RNA聚合酶Ⅰ,RNA聚合酶Ⅱ和RNA聚合酶Ⅲ的参与。染色质组蛋白对DNA的转录具有调节作用。当组蛋白与DNA结合时,能掩盖RNA合成的起始位点,阻止RNA聚合酶与起始位点接触,从而抑制了转录。然而,非组蛋白与组蛋白相反,却能解除组蛋白对DNA的抑制作用。关于非组蛋白对DNA的转录的调节机制,目前虽尚未完全阐明,但通常认为可能是非组蛋白由于磷酸化而带负电荷,能在DNA识别位点附近与带正电荷的组蛋白结合,而使DNA一组蛋白复合物构象发生改变,此时组蛋白一非组蛋白复合物发生部分转移,致使DNA裸露,显露出许多RNA聚合酶结合位点,RNA聚合酶与结合位点接触,而引起DNA转录RNA。
染色质DNA除转录RNA之外,还能复制自身。DNA复制自身时,以其一条多核苷酸链为模板合成新链,称为半保留复制。真核细胞进行DNA复制时可有多个起始点,如中国地鼠细胞中的整个基因组共有50 000个起始点。复制从起始点开始,向两个方向进行,直到终点。现在已在电子显微镜下观察到DNA复制时所必需的DNA聚合酶。DNA聚合酶为直径6.5nm的粗糙圆形体,间隔地附着DNA链上。在DNA酶参与下所合成的DNA并不连续,而是一些片段,这些片段再由DNA连接酶连接在一起。
染色质
chromatin
- 完成任务后,向有关方面报告结果是什么意思
- 完成体是什么意思
- 完成其事是什么意思
- 完成前人未竟之业是什么意思
- 完成前行为是什么意思
- 完成变态昆虫的幼体是什么意思
- 完成句子是什么意思
- 完成句子测验是什么意思
- 完成台本是什么意思
- 完成合同法是什么意思
- 完成后行为是什么意思
- 完成学业是什么意思
- 完成 实现是什么意思
- 完成家业是什么意思
- 完成工作的经济合同是什么意思
- 完成工时是什么意思
- 完成差事后回报是什么意思
- 完成很容易是什么意思
- 完成态是什么意思
- 完成情况是什么意思
- 完成情况(完成质量)是什么意思
- 完成所交给的任务是什么意思
- 完成投资是什么意思
- 完成某事是什么意思
- 完成某项事业是什么意思
- 完成某项工作的期限是什么意思
- 完成百分比法是什么意思
- 完成的事实构成是什么意思
- 完成行为是什么意思
- 完成貌是什么意思
- 完成配音的影片是什么意思
- 完房是什么意思
- 完扇记是什么意思
- 完护是什么意思
- 完捐是什么意思
- 完操是什么意思
- 完整是什么意思
- 完整不可分的样子是什么意思
- 完整体同一认定是什么意思
- 完整准确地宣传马克思主义是什么意思
- 完整制是什么意思
- 完整制与分离制是什么意思
- 完整句是什么意思
- 完整地准确地理解毛泽东思想是什么意思
- 完整地奉还是什么意思
- 完整式是什么意思
- 完整性是什么意思
- 完整情报是什么意思
- 完整教学法是什么意思
- 完整无缺是什么意思
- 完整无缺损的东西是什么意思
- 完整无缺的样子是什么意思
- 完整无缺;支离破碎是什么意思
- 完整 残缺是什么意思
- 完整法是什么意思
- 完整的东西出现裂缝是什么意思
- 完整的东西破成零片零块是什么意思
- 完整的书是什么意思
- 完整的事物是什么意思
- 完整的文章是什么意思