操纵子operon
原核生物中,由一群基因组成的遗传调控单位。主要包括一个操纵基因和位置邻近的若干个结构基因。按照法国遗传学家雅各布(F.Jacob)和莫诺(J.Monod)在20世纪60年代初提出的操纵子假说,紧密连锁的一组结构基因,共同受一个操纵基因的控制,在转录功能上,形成一个整体。
操纵子caozongzi
基因对酶合成控制的学说。按
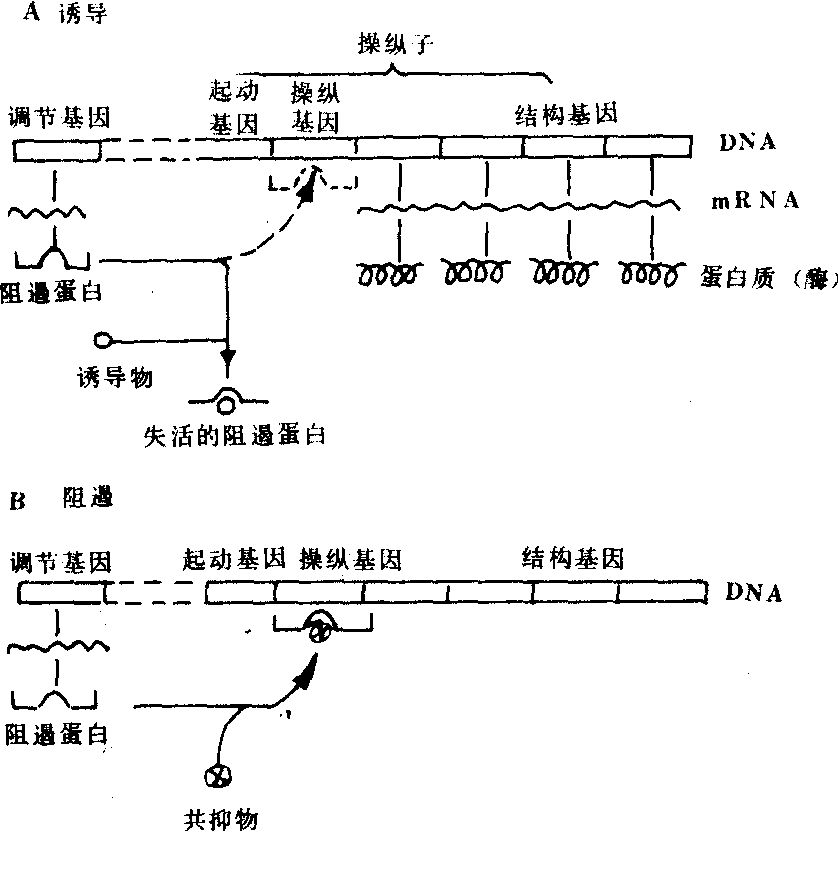
图588 基因对酶合成的调节
A.诱导:调节基因产生阻遏蛋白,后者阻遏操纵基因,从而使结构基因不能产生mRNA。当加入诱导物时,
阻遏蛋白失活,于是,操纵基因又使结构基因恢复功能。
B.阻遏:阻遏蛋白只有同共抑物结合后才有阻遏活性,
而共抑物是结构基因产生的一种酶的代谢产物。
操纵子operon
系DNA上由相关基因组成的功能单位。此功能单位包括1个或几个功能相关的成簇结构基因及操纵基因和启动基因。1个操纵子在转录中只形成1个大的单一的信使核糖核酸分子;在原核细胞中,操纵子基因所编码的蛋白质一般为催化同一代谢过程中功能相关的酶;操纵子可由于细胞生长环境的变化而改变转录信使核糖核酸的频率。
操纵子
由操纵基因以及紧接着的若干结构基因共同组成的一个超基因的功能单位。其中结构基因的转录由操纵基因所控制。
操纵子operon
是由操纵基因和一个或一个以上的结构基因组成的基因簇。调节和控制某一代谢过程中酶的合成,如大肠杆菌乳糖操纵子,调节和控制乳糖代谢过程中酶的合成。
操纵子
操纵子是指在脱氧核糖核酸 (DNA)中,由互相邻接的包括结构基因、操纵基因和起动部位等基因所组成的一种遗传调节的功能单位。这些基因的功能是由同一操纵基因与阻抑蛋白来联合控制的。
多年来已知细菌体内存在着一类称为 “操纵子” 的基因调节单位。这一调节单位控制着某种信使核糖核酸(mRNA)分子的基因转录,由此间接控制一种或一套蛋白质(如酶)分子的生物合成。据知操纵子由信息区,以及与之相邻的控制区共同构成; 信息区含有转录生成mRNA的信息,由其相应的控制区控制其是否转录,由之控制这些信息的表达(图1)。
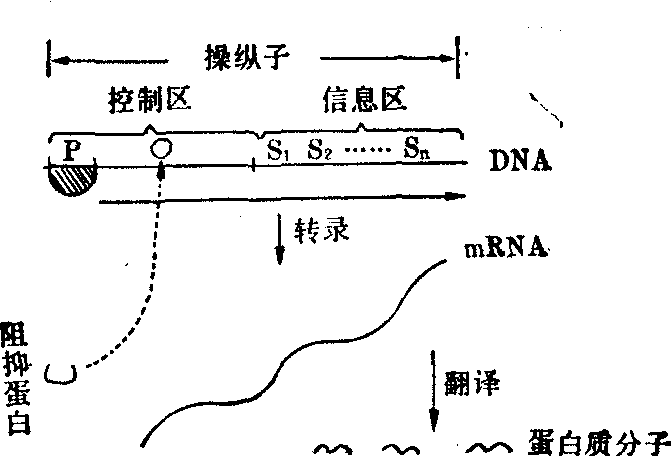
图1 操纵子的模式图
O:操纵基因 P:转录的起动部位 S:结构基因n:结构基因数
阻抑蛋白: 可特异地阻抑DNA结构上相应操纵子的转录 诱导物(图中未表示): 可使阻抑蛋白失去阻抑作用,mRNA得以生成。
控制区 控制区有一专供核糖核酸聚合酶系附的转录起动部位。与起动部位相邻,有一控制部位,这一控制部位即称为“操纵基因”。操纵基因可与特异的阻抑蛋白结合。这类阻抑蛋白若与操纵基因结合,则可阻碍核糖核酸聚合酶,于此顺着DNA进行转录。此时这一操纵子即被阻抑。
特异的诱导物,例如某些酶的诱导物,可与其相应的阻抑蛋白结合,从而使该阻抑蛋白与操纵基因分离,从DNA上脱落,此称之为脱阻抑。脱阻抑状态的操纵子即有可能被转录。除脱阻抑外,有时还需某种蛋白质(或该类蛋白质与cAMP等小分子结合在一起的复合物) 附着于起动部位,使催化核糖核酸合成的核糖核酸聚合酶易于起动。
阻抑蛋白起着阻抑作用,它的作用是一种负调节。而上述蛋白质或蛋白质复合物起着促进转录的作用,它们的作用是一种正调节。
信息区
信息区由一个或多个基因组成,由于信息区所含的基因起着规定蛋白质结构的作用,所以这些基因称为结构基因。
由操纵子转录生成的mRNA,即可按操纵子所含结构基因数的不同而被翻译成为一种或数种多肽链或蛋白质。
在操纵子之外,还存在着为上述阻抑蛋白编码的基因,这类基因称为调节基因。调节基因如因突变而丧失功能,则阻抑蛋白缺失,其相应的操纵子过分表达,因此而合成的蛋白质可因之过剩。
操纵子学说自1961年由法国的Jacob和Monod根据对大肠杆菌中与乳糖代谢有关的操纵子——乳糖操纵子,与阿拉伯糖代谢有关的操纵子——阿拉伯糖操纵子,以及某种沙门杆菌的组氨酸操纵子等已经逐渐弄清。
大肠杆菌乳糖操纵子是第一个被彻底研究过的操纵子模型。现以这一操纵子为例具体说明之,该操纵子在大肠杆菌中调节乳糖代谢。乳糖操纵子含有三个结构基因,各自含有合成β-半乳糖苷酶、透性酶以及半乳糖苷乙酰化酶的信息。除半乳糖苷乙酰化酶的生理功能还不清楚外,β-半乳糖苷酶催化乳糖水解为半乳糖与葡萄糖,透性酶可使半乳糖易于透入菌体。
大肠杆菌的上述酶活性在通常情况下不高,给以乳糖或某些类似乳糖的诱导物时,上述三种酶的合成速率增高,活性可由此增加10~100倍。但当培养基中存在葡萄糖时,此种诱导现象并不出现。直至葡萄糖耗竭,诱导现象才出现。这是因为大肠杆菌乳糖操纵子的表达,不仅需有诱导物,以解除阻抑蛋白的阻抑,还需要某种蛋白质(代谢降解物基因活化蛋白(CAP)与cAMP的结合物附着于起动部位,使核糖核酸聚合酶催化的转录作用易于起动(图2)。
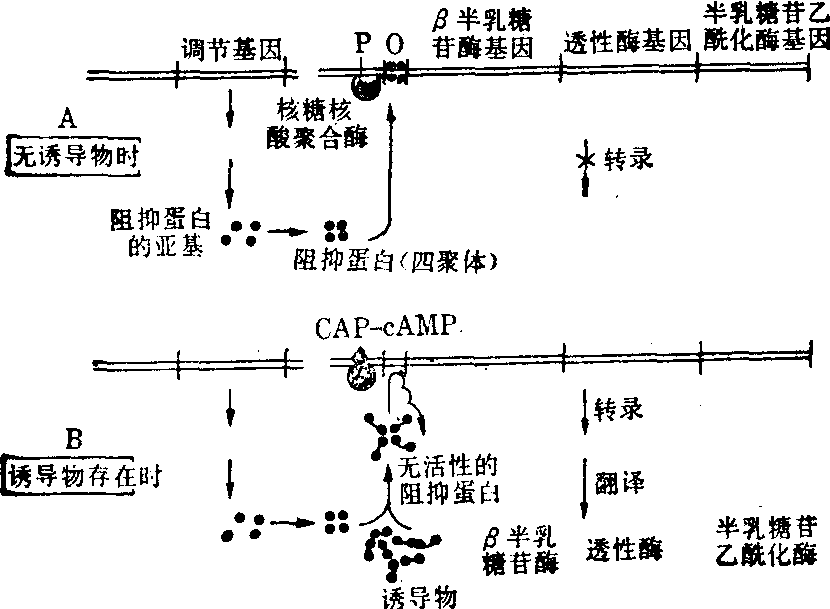
图2 乳糖操纵子的阻抑与脱阻抑
O——操纵基因 P——转录的起动部位
在无诱导物存在时 (图2A),阻抑蛋白与操纵基因结合,妨碍核糖核酸聚合酶的起动,从而阻止这一操纵子的转录,mRNA因而难以合成。在诱导物存在时(图2B),阻抑蛋白因与其结合,不能附着于操纵基因,如此时存在前述cAMP与CAP的结合物,则RNA聚合酶即可转录上述三种结构基因,生成mRNA。此mRNA如被翻译,即产生β-半乳糖苷酶、透性酶与半乳糖苷乙酰化酶。乳糖的分解代谢由此即可得到促进。已知乳糖操纵子的阻抑蛋白由四个分子量为40 000的同样亚基缔合而成。
其他操纵子的调节机制与乳糖操纵子者大同小异,例如大肠杆菌的阿拉伯糖操纵子,可因其操纵基因与特异的阻抑蛋白结合而被阻抑,但在阿拉伯糖操纵子中,可与其起动部位结合而起正调节作用者,并非cAMPCAP,而是阿拉伯糖与该操纵子特异阻抑蛋白的结合物。
在细菌等原核生物中,操纵子遗传调节单位的存在已经确立。但在哺乳动物等真核生物中是否也存在着类似的调节单位尚得进一步研究。
操纵子
操纵子是DNA分子内由一定的核苷酸序列组成的一个基因表达的协同单位,包括几个结构基因及基因表达的共同的调控序列。结构基因也称顺反子,是为多肽链编码的核苷酸序列,从起始密码子ATG始至终止密码子TAA、TGA或TAG止,其间有各种三联体密码子,决定多肽链的氨基酸排列顺序,每个顺反子之间有时也存在少许不翻译的序列,调节基因编码阻遏物,也列入操纵子的结构基因中。
基因调控序列主管基因转录的调控,其中有转录起始、转录通行、转录终止或停顿的各种调节位点。一些位点的核苷酸序列常存在着二元对称性,形成链内的二级结构。
可诱导操纵子 许多负责糖分解代谢的酶类都由这类操纵子合成,如乳糖操纵子、半乳糖操纵子等。在不需要从所负责分解代谢的糖类获取碳源时,该操纵子的阻遏物与操纵基因结合,转录进行得很少,当诱导物存在时可与阻遏物结合,使之从操纵基因上脱落,转录通道畅通,转录水平升高。
可阻遏操纵子 目前已知负责氨基酸合成的操纵子如色氨酸操纵子、组氨酸操纵子等,都属于这一类。它们所负责合成的终产物氨基酸是辅阻遏物,对转录进行负反馈作用。这个氨基酸也是衰减作用的调节物,当细菌生长在该氨基酸过剩的环境中时,由于氨基酸与阻遏物蛋白一起构成阻遏物,结合在操纵基因上而阻碍转录,同时衰减子起衰减作用。当处于该氨基酸饥饿状态时,操纵基因上没有阻遏物占位,同时出现反衰减作用而使转录得以进行。
操纵子的基因转录调控序列负责转录的起始、延伸、终止等的调控,根据各部位所起的作用不同可分为:
❶操纵基因,是调节因子-阻遏物的结合部位,二者结合转录即被阻断或速度大大减慢,这是转录的负调节。
❷启动子,是RNA聚合酶结合和启动RNA合成的部位。
1975年Pribnow比较了几种原核生物的RNA聚合酶保护片段的序列,发现有一个七核苷酸的同源区5′-TAT-AAATG,其中心点大约在mRNA转录起始点上游约10bp (碱基对)的位置,通常称为-10区,也称TATA盒或pribnow盒,其中TA,TAA及第7位T具有很强的保守性。后来Schall等人发现用纯化的上述片段再与RNA聚合酶作用时则该酶不能选择正确的引发点,说明还有TATA盒以外的序列对酶识别启动子是必需的。对噬菌体λ和SV40病毒DNA的研究,说明启动子区应从mRNA转录的起始点向上游延伸到35bp以远,大肠杆菌的一些启动子需从转录起始点向上游延伸到60 bp以远,比较了大肠杆菌46个启动子的结构,除发现-10位的保守序列外,-35位左右的5′-TTGACA也有很强的保守性,虽不同来源结构不尽相同,但TTG的保守性很强。
此外,有些调节因子如降解物基因活性蛋白(CAP)的结合部位也在启动子内。CAP 是由两个亚单位构成的环磷酸腺苷 (cAMP)受体蛋白,分子量为45000,与cAMP一起可增强RNA聚合酶与启动子的结合而加快转录速度,所以启动子的结构一般应包括:
❶RNA聚合酶的识别信号-35位左右的序列;
❷RNA聚合酶紧密结合位点-10位左右的序列;
❸RNA合成的引发点;
❹某些调节因子的结合位点。
启动子与操纵基因相邻,接点处有时有重叠的部分。
衰减子 转录暂时停顿的信号,只存在于某些操纵子中。1974年Kasai在研究伤寒杆菌组氨酸操纵子时,发现其中存在着转录的障碍,如在启动子与第一结构基因之间造成缺失突变,可使结构基因转录增强,随后在大肠杆菌色氨酸操纵子及其他氨基酸合成的操纵子中相继发现了类似的结构。用缺失突变及体内外转录实验证明,这一部分序列的作用与操纵基因无关,定位在操纵基因及第一结构基因之间,通常称前导序列。多由140~200个核苷酸组成,其中包括一独立的转录单位,可转录一个小分子mRNA和编码一个小肽。mRNA 5′端有rRNA结合位点,3′端有GC富集区,可形成稳定的链内二级结构,紧连数个U的尾端,是典型的终止子结构,在这里叫衰减子。mRNA编码部分含有数个相连的调节密码子,即操纵子终产物氨基酸的密码子,以行使调控的职能。
终止子 转录终止的部位,在DNA上有共同的结构特点,即有GC富集区,紧跟着AT富集区,具二元对称性,所以转录出的mRNA可形成发卡环,并带有数个U的尾端。在某些操纵子中也存在于前导序列内,以行使衰减作用的调控。根据终止作用发生的情况,可分为简单终止子及复杂终止子二类。
简单终止子,即RNA聚合酶停止转录不需要其他因子参予。发卡环式二级结构对终止转录起重要作用,实验证明,凡是增强终止子发卡环的突变,均可增强终止作用,反之则减弱终止作用。
复杂终止子,RNA聚合酶在某些终止子位点上停止转录还需要一些因子参予作用,如大肠杆菌中的ρ因子,NuSA蛋白质,L因子等。ρ因子是首先在大肠杆菌内发现的可使噬菌体λ DNA转录终止的依赖因子,具有核苷三磷酸酶的活性,并可与单链RNA结合。最近已有报道,提出ρ因子具有螺旋酶的活性,可使短的RNA-DNA双链解旋,RNA易于自模板上解脱,同时水解核苷三磷酸以供给能量。
兹举乳糖操纵子和色氨酸操纵子为例说明如下。
乳糖操纵子: 大肠杆菌分解乳糖的酶类由乳糖操纵子(lac)合成,全长6.5kb(千碱基),由结构基因lacZ、lac-Y、lacA组成多顺反子,按顺时针方向转录。lacZ由3.063kb组成,编码β-半乳糖苷酶,分解乳糖。lacY由1.251kb组成,编码乳糖通透酶,为乳糖进入细胞所必需。lacA由0.81kb组成,编码硫代半乳糖苷转乙酰酶。lacI由1.080kb组成,编码阻遏物,通常称为调节基因,有其自身的调控系统。基因转录的调控序列由122bp(碱基对)组成,在lacI与lacZ之间,按照调控功能可分为:
❶mRNA转录起始点,通常以这一位点的核苷酸定为1,上游为负(-),下游为正(+)。
❷启动子(lacP),自-84位至-1位,-10位与-35位的核心序列分别为5′TAT-GTTG与5′TTTACA,是RNA聚合酶结合位点,有人提出-22位左右,还有一弱的P2启动子。自-84至-55位是CAP结合位点。
❸操纵基因(lacO),自+1至+28位,是阻遏物结合部位,另有10个碱基对是在lacO与第一结构基因lacZ的间隔区。lacO与lacP 之间常有几个核苷酸的重叠,并存在二元对称结构。
乳糖操纵子的基因表达与环境中的乳糖浓度有关。当细菌生长不需自乳糖获取碳源时,阻遏物与lacO结合,呈负调控状态。此时转录极低,只有几个分子的β-半乳糖苷酶生成。生理状态下的诱导物是异乳糖,由乳糖经转糖基化作用生成,原来细胞中存在的少量β-半乳糖苷酶可催化异乳糖的合成。当乳糖成为细菌生长的唯一碳源时,诱导物即与阻遏物结合,操纵基因开放,转录立即进行。以β-半乳糖苷酶的产量计算,诱导后,可提高1000倍,这是正调控。但是只有诱导物存在还不够,乳糖操纵子的完全表达还需要CAP-cAMP参予。CA P是环磷酸腺苷(cAMP)受体蛋白,其作用可能是使RNA聚合酶结合位点下游30~50bp处的双螺旋松弛而增加转录的有效性。cAMP可促进CAP与启动子结合。环境中葡萄糖存在时,使cAMP浓度降低,所以葡萄糖对乳糖操纵子的表达有抑制作用。乳糖操纵子及其调控方式见图1。
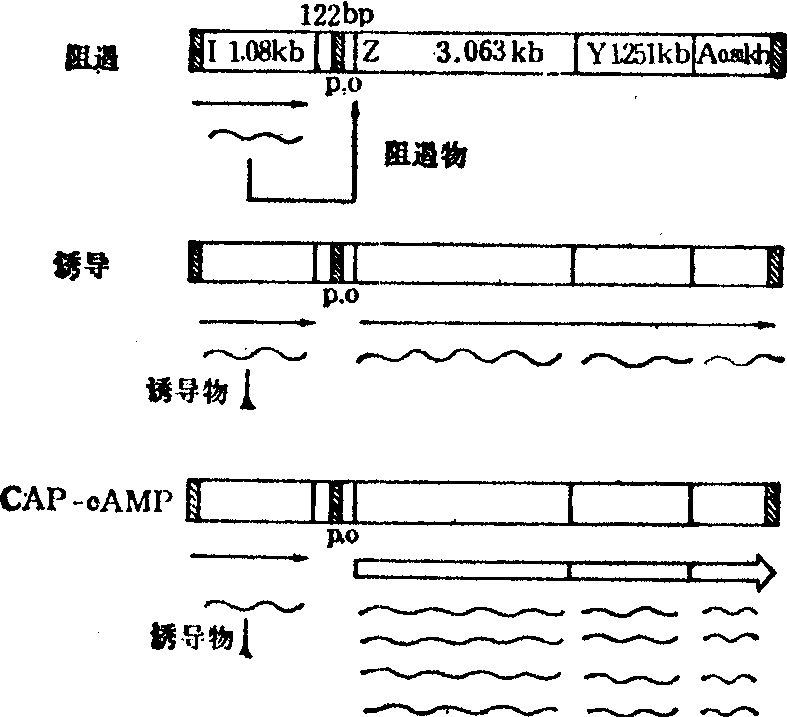
图1 乳糖操纵子及其转录的调控
色氨酸操纵子: 大肠杆菌合成色氨酸的酶类由色氨酸操纵子(trp)合成,由结构基因trpA、trpB、trpC、trpD、trpE组成多顺反子,按反时针方向转录,即自trpE依次至trpA。trpR编码阻遏物蛋白,有本身转录的调控系统。
色氨酸由分枝酸合成,可分为五个步骤,前两步将分枝酸变成氮-5-磷酸核糖酰-邻位氨基苯甲酸盐(N-5′-phosphoribosylanthranilate)的酶,是trpE (1.8kb)及trpD(1.8kb)基因的产物,trpC (1.356kb)编码的酶具有二种活性(即磷酸核糖酰邻位氨基苯甲酸异构酶和吲哚甘油磷酸合成酶)。最后一步所需的色氨酸合成酶是trpA(0.8kb)和trpB(1.191kb)基因的产物,五个基因产物的协同作用最终合成色氨酸。
操纵子的基因转录调控序列由76个碱基对组成,主要启动子trpP由-59位至-6位,操纵基因trpO与启动子有较多的重叠,-10位左右存在着明显的二元对称序列。此外在trpD内还有一弱的启动子P2。
基因调控序列与第一结构基因trpE之间存在着前导序列L,由162碱基对组成,是衰减子trpa的部位。
基因转录的调控首先由阻遏与去阻遏作用作进行调节。色氨酸操纵子表达的结果,最终合成了色氨酸,色氨酸又是辅阻遏物,与trpR编码的阻遏物蛋白一起与操纵基因结合,阻遏转录,当培养基中色氨酸供应不足时,由于缺乏辅阻遏物而出现去阻遏作用使操纵基因开放,转录进行。所以环境中的色氨酸浓度对调控转录起主要作用。但是trpD内的P2启动子不受trpO与trpA调控的影响,它启动的trpC、trpB、trpA的转录即使在高色氨酸的环境中也可进行,因此在阻遏状态下,P2启动子完成的转录可占全部产物的80%。色氨酸操纵子及转录调控方式见图2。
除阻遏作用外,还有衰减作用的调控。由于操纵子的前导序列可转录一个140核苷酸的mRNA,上面有rRNA结合位点,AUG起始码(在第27~29位)和UGA终止码(在第69~71位),可翻译一个14肽,肽链第10和11位的密码子是两个相邻的UGG,编码色氨酸,称为调节密码子,在3′端有GC富集区和多U序列是终止子构型,称为衰减子。前导序列中的二元对称结构存在着多重性,可形成变化的发卡环。当色氨酸缺乏时,细菌需要自己合成,这时14肽的翻译由于缺乏色氨酸而中止在第10、11位密码子处,rRNA占位使原来1~2、3~4二组二元对称结构的1~2发卡环破坏,而2~3成环。2~3成环后就破坏了原来的3~4终止子的构型,其结果是反终止作用而使RNA聚合酶转录顺利进行,全部结构基因得以表达,而合成出色氨酸。当环境中色氨酸过量,细菌不需要自己合成时,由于14肽的生成,rRNA无占位,1~2发卡环存在,自然3~4也成环,这时3~4环是终止子结构,使R-NA聚合酶识别而转录暂时停顿,这一衰减反衰减作用也是由色氨酸进行调节的(图3)。
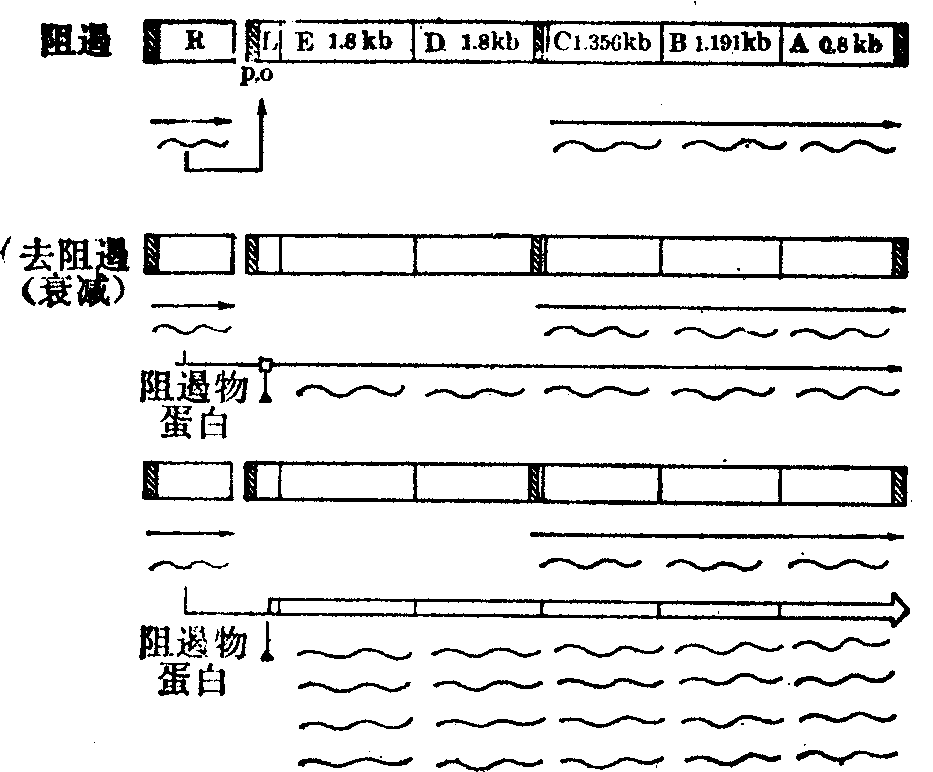
图2 色氨酸操纵子及其转录的调控
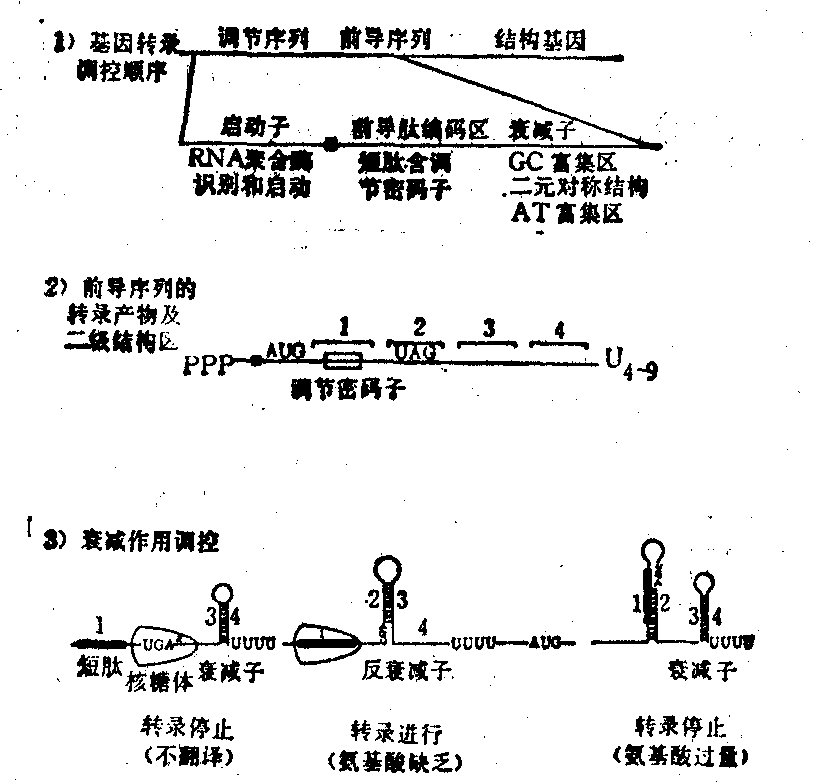
图3衰减子调控机制
操纵子operon
操纵子结构是原核基因组的突出结构特点。是指数个功能上相关联的结构基因串联在一起构成信息区,连同其上游的调控区和下游的转录终止信号构成基因表达单位。
操纵子
operon
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名之下,其实难副是什么意思
- 盛名铭是什么意思
- 盛唐是什么意思
- 盛唐遗恨是什么意思
- 盛国英是什么意思
- 盛国荣是什么意思
- 盛国荣是什么意思
- 盛国荣医学论文集是什么意思
- 盛增秀是什么意思
- 盛墩是什么意思
- 盛夏是什么意思
- 盛大 浩大是什么意思
- 盛大在线是什么意思
- 盛如梅是什么意思
- 盛如梅是什么意思
- 盛如梓是什么意思
- 盛妆浓饰是什么意思
- 盛婕是什么意思
- 盛孝基是什么意思
- 盛宏正是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀是什么意思
- 盛宣怀(1844—1916)是什么意思
- 盛宣怀传是什么意思
- 盛宣怀未刊信稿是什么意思
- 盛宴是什么意思
- 盛家伦是什么意思
- 盛家兴是什么意思
- 盛寿藻是什么意思
- 盛少俊是什么意思
- 盛展武是什么意思
- 盛巽昌是什么意思
- 盛希侨是什么意思
- 盛希侨是什么意思
- 盛年难再是什么意思
- 盛庆琜是什么意思
- 盛度是什么意思
- 盛度是什么意思
- 盛度是什么意思
- 盛度是什么意思