抗药性遗传genetic of resistance to insecti-cide
从分子、细胞、个体和群体水平研究昆虫抗药性的遗传变异规律,主要包括抗药性的形成、遗传方式、连锁群、基因的相互作用、稳定性和共适应、染色体易位和倒位的影响、基因扩增、基因调控,以及基因在群体中的数量变异等。抗药性是通过个别基因组中的突变而产生的。这些突变主要是DNA碱基对取代、原先存在的抗药性基因扩增、易位和染色体倒位,或其他的DNA重排。其中DNA碱基对取代与靶标抗药性的关系较为密切,因为靶标大分子可发生变化,以致不再与杀虫剂结合; 在代谢抗药性中,基因扩增和DNA碱基对取代都是重要的。
简史 迪克森(R.C.Dickson)1941年率先对昆虫的抗药性作遗传分析,指出美国加利福尼亚州的红蚧Aonifiella aureantii对氢氰酸是性连锁,呈不完全显性。在50年代对抗药性的形成有两种解释: 前适应学说和后适应学说。60年代日本学者塚本增久应用杂交后的毒力回归线的形状来分析不同基因型的分离情况,从而推断基因遗传方式。随后由于在双翅目昆虫果蝇、家蝇和蚊虫等中,发现了许多可用作抗药性遗传分析的隐性标记品系,为确定抗药性(R)基因的连锁群、染色体定位、R基因的相互作用以及染色体畸变对抗药性的作用等研究奠定了基础。80年代分子生物学的发展促进了分子遗传的研究,随着分子生物学和基因工程的发展,正成为一个十分活跃的研究领域。计算机的应用,也促进了抗药性群体遗传学的研究,其中包括R基因在种群中的动态变化及其调节机理等。
抗药性形成学说 由于细菌的抗药性是来自基因突变,还是对环境的适应性变异,是一个长期争论不休的问题,因此对昆虫抗药性也存在两种解释: 前适应学说和后适应学说。前适应说认为抗药性是由于药剂的不断选择,使个体中的不定向变异(突变)逐渐成为群体中的遗传结构定向变异,即药剂仅是对早就存在的突变基因起选择作用。而后适应说认为原来种群中不存在突变基因,而是由于不断使用药剂使昆虫适应性发生变异。
目前大多数人倾向前适应学说,即认为是由于药剂选择的结果。因为: ❶用亚致死剂量处理昆虫不能产生抗药性,而某些昆虫在处理后的抗药性不是增加反而是减弱。一定要用高浓度处理引起大量死亡后才能使后代的抗药性增加,如果不引起死亡则不发生抗药性。
❷某些昆虫不用药剂处理,而仅是通过逐代选育生活史长的个体,也能分离出抗药性(R)品系。而且不管用什么方法选育,只要一旦在室内选育成纯系的R品系,尽管不再用药,但其抗药性就不会消失。
❸许多杀虫剂对昆虫的某些解毒酶,如多功能氧化酶和谷胱甘肽转移酶等有诱导作用,而增强了其对药剂的抵抗能力,但这种抗药能力在作为诱导剂的杀虫剂不存在时也就随之消失。也就是说,这种诱导而产生的抗药能力是不能遗传给下一代的。
遗传方式 主要包括抗药性的显隐性、性连锁、单因子和多因子遗传等。
显隐性及其测定 由于抗药性无形态特征,只能借助于对数剂量—死亡机率值回归线(logdosage-probit mortality regression line,简称LD-p线)和隐性标记品系来测定其遗传方式。在一般情况下,LD-p线只有在用于测定的品系是敏感纯合子(SS或++),抗药性纯合子(显性时为RR,隐性时为rr),以及它们的杂交子一代(F1)的杂合子(显性时为RS或R+,隐性时为r+)时才呈真正的直线关系。在子二代(F2),以及F1与SS或RR回交后的群体中,LD-p线可因其表现型分离的相对比例不同而呈不同的形状(图1)。如RR和SS杂交后F1的LD-p线接近于RR,即其抗药性与RR显性表现型一样高,则为完全显性(线a)。反之,若F1的LD-p线接近于SS纯合子,即其抗药性如同SS隐性表现型,则为完全隐性(线b)。若F1的LD-p线
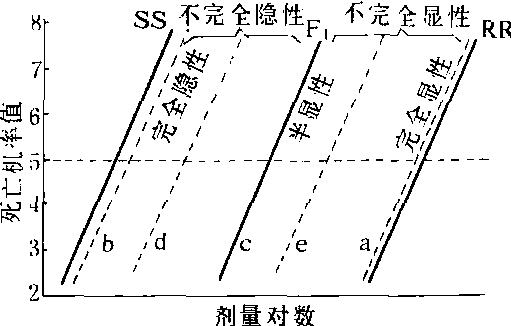
图1 RR和SS杂交后F1的抗药性显隐性图解
(据本增久等,1964)
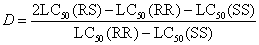
表1 RR和SS杂交后F1的LD-p线、显隐性和显性度的关系
| LD-p线的位置 | 显 隐 性 | 显性度 |
| 接近RR (a) 接近SS (b) | 完全显性 完全隐性 | 1 -1 |
| 位于a-b的1/2处(c) | 半显性 | 0 |
| 位于a-c之间(d) 位于b-c之间(e) | 不完全显性 不完全隐性 | 0 |
注:括弧中的a,b,c,d和e分别代表图1中的LD-p线。
性连锁及其测定 若互交F1的雌雄比例无明显差异,则R因子位于常染色体。如果所有的雄性是抗药性的,所有的雌性是敏感的,则表明涉及性连锁,或涉及常染色体雄性决定因子(autosomalmale-determining factor)。例如家蝇中的性限制滴滴涕抗药性就是一例。
单因子和多因子遗传及其测定 如果每个基因型的LD-p线互相不重叠,而R基因又是完全隐性,则F2的LD-p线约在死亡率75%(机率值5.675)处有一个明显的“平坡”,即表示敏感的和抗药性的表现型为3∶1(图2a)。这一平坡即为有效诊断剂量,又称区分剂量的范围。如果抗药性表现型的水平不是很高,则就不会有明显的平坡。
在完全显性时,死亡率25%(机率值4.326)处有一个明显的平坡(图2c),表示抗药性表现型(RR和R+)和敏感(++或SS)的分离比例为3:1。
若为半显性时,F2的分离应是敏感纯合子(++):杂合子(R+):抗药性纯合子(RR)=1:2:1。若它们的LD-p线互相不重叠,则F2的LD-p线在死亡率25%和75%处各有一个明显的平坡(图2b线1)。但是在大多数交配试验中,杂合子的抗药性程度取决于R基因的显性度。因此,在杂合子的LD-p线和任何一个亲本品系的LD-p线互相重叠时,也许只有一个平坡(图2b线2)。
在R+和++(或RR)回交时,其F1的LD-p
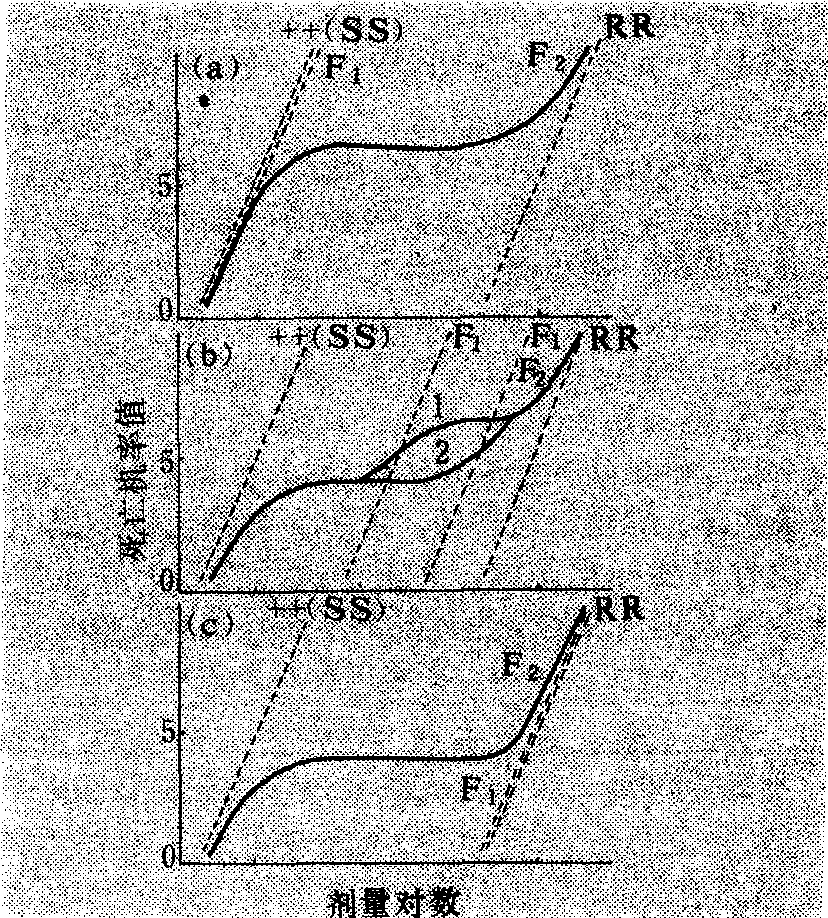
图2 单因子抗药性(RR)和敏感(++)品系杂交后F2的LD-p线的形状
(据本增久等,1964)
(a)完全隐性; (b)不完全显性; (c)完全显性
❷)处理回交F1,则++(或RR)和R+的分离比例为死亡数(++):存活数(++):存活数(R+)=1:1,或死亡数(R+):存活数(RR) =1 :1。
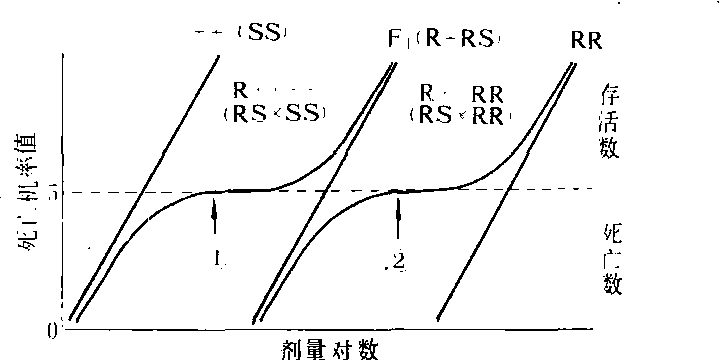
图3 R+和++及R+和RR回交F1的LD-p线
(据本增久等,1964)
❶和
❷分别为++和R+以及R+和RR的区分剂量
在RR、R+和++基因型的LD-p线部分重叠时,则必须计算F2和回交F1对每一个剂量浓度的反应期望值。在F2中:
xy=0.25w(++)+0.5w (R+)+0.25w(RR)在F1(R+)与++(或RR)回交中:
xy=0.5w (+R)+0.5w(++或RR)
上两式中,xy为剂量 y时的期望值; w为++、+R和RR基因型在剂量y时的观察值,即直接从相应的LD-p线上获得。在涉及两个因子以上的多因子时,情况就更为复杂。难于用LD-p线的形状变化来推测。一般讲,若F2的LD-p线几乎呈直线形状,则可视为多因子遗传。
连锁群分析 从上可见,抗药性可涉及一个或几个遗传因子。如要确定抗药性基因在染色体上的位置及其连锁群,则必须应用标记品系。由于大多数昆虫遗传组成太复杂,特别是农业害虫,染色体数目多达50~60对,现在抗药性遗传的研究仅局限于双翅目的果蝇、家蝇和蚊虫等。家蝇的染色体数目为2n= 12,即6对染色体,其中1对为性染色体,5对为常染色体。除性染色体外,其余5对染色体均已有多种隐性突变标记基因。其主要抗药性因子的连锁群,
各个R基因所在的染色体及其显隐性关系见表2。
表2 家蝇抗药性基因的连锁群
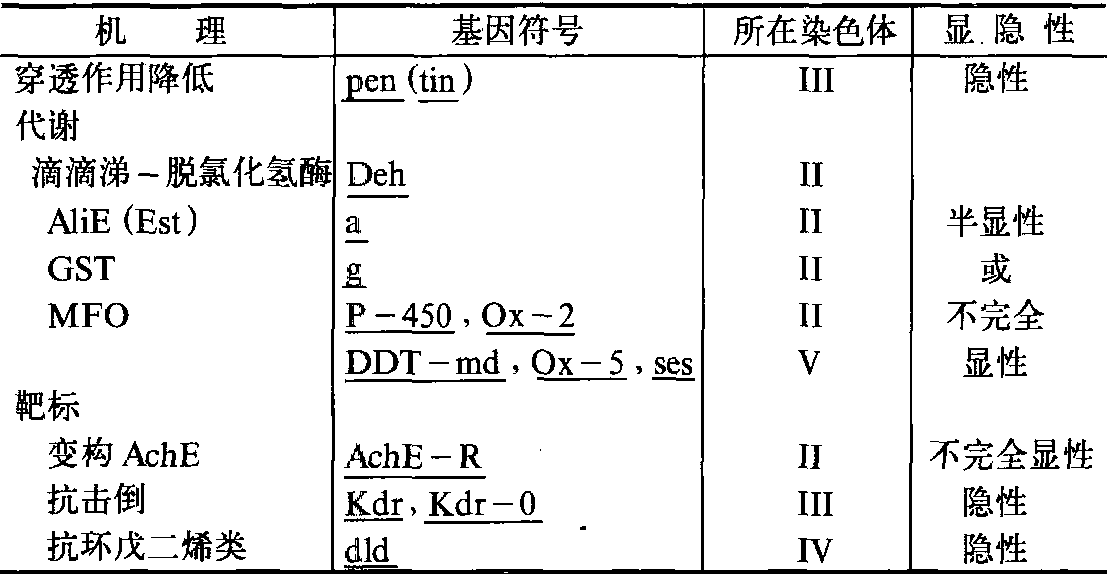
几乎所有涉及代谢的因子都位于染色体Ⅱ上,而且都呈显性。另外,控制不同形式的多功能氧化酶(MFO)的基因也是不同的。在家蝇中,与滴滴涕氧化作用有关的MFO基因(ses,DDT-md和Ox-2)位于第V染色体,而与有机磷、氨基甲酸酯抗药性有关的MFO基因(Ox-2和P-450)位于第Ⅱ染色体。
基因相互作用 R基因相互作用时对抗药性水平的影响见表3。两个基因结合时对抗药性水平的影响
表3 家蝇抗药性基因的相互作用对抗药性水平的影响
(据F.W.Jr.Plapp,1970)不是简单的相加而是倍增。其作用机制尚不清楚,但可肯定,R基因在染色体上的位置对抗药性水平影响很大。若R基因与其他基因无交换或交换率低,则抗药性就越高。
共适应与稳定性 共适应(coadaptation)是指通过选择作用和R基因的整合作用(integration)改善R基因的有害作用,使其逐步适应变化的环境。在抗药性形成的早期,抗药性不是升高,反而会下降。如作进一步选择,则抗药性变得更为稳定,即使停止药剂选择后仍较稳定。这就是共适应的结果。另外,还发现某些修饰基因在R基因与适合度因子整合过程中起着重要作用,它们可帮助将R基因整合到基因组的其余基因中去,从而产生一个协调的共适应基因组(harmoniously coadapted genetic)。R基因取代其野生型(敏感)基因可产生许多多效效应(pleiotropic ef-fects),其中有些作用似乎是有害的,所以开始时抗药性基因表现出适合度低。随后由于对修饰因子的选择而对多效效应的有害作用得到补偿,此时这种劣势会减轻或消失,使抗药性更稳定。
染色体易位的影响 布莱克曼(RL.Blackman)等在1978年分析了世界各地桃蚜的第Ⅰ和第Ⅲ染色体(A1,3)之间的易位,对有机磷抗药性的影响后,发现A1,3易位和R基因是完全连锁的。易位可以通过抑制交换来维持其抗药性。另外,易位可使R基因产生有利的重排,把A1,3上的R基因带入同一连锁群。易位还可使异染色质(chromatin)因位置的改变而缩短了与R基因间的距离,从而对R基因产生一定的影响。
染色体倒位的影响 如表2所示,家蝇的代谢R基因尽管它们的作用功能不同,但它们大都位于第Ⅱ染色体上。并发现 在R家蝇品系的第Ⅱ染色体上普遍存在倒位,而敏感品系则没有。染色体倒位可加强这段染色体不含着丝点,则减数分裂的倒位和未倒位的染色体配对时会抑制这段染色体的交换。生物体常通过这种作用使一些功能相关的基因连锁起来成为一个基因群,从而加强它们对新陈代谢的作用。同一种生物,染色体倒位频率高的群体对环境的适应力要高于倒位频率低的群体。如家蝇第Ⅱ染色体上的倒位对这些代谢基因具有连锁作用。拥有R基因的个体,抗药性增加的幅度是有限的,必须加上染色体倒位才能使抗药性达到高峰。如含有R基因而不具倒位的家蝇品系的生殖力很低,即使用药剂筛选,抗药性仅增加3倍,而具有R基因和染色体倒位的家蝇则十分健康,而且抗药性很高。
基因扩增作用 基因扩增是指同一染色体上相同基因出现两次以上,即有两个以上的拷贝,故又称之为“串连重复”(tandem duplication)。R基因复增是抗药性进化中的一种普遍现象。桃蚜的抗药性和酯酶(E4)活性呈正相关。后来德文希尔(A.L.Devonshire)和萨维斯基(R.M.Sawicki)在1979年发现桃蚜的七个无性系的E4量,以级比为1.91的几何级数,从一个对有机磷敏感的无性系的E4量为1单位,依次增加到高抗药性系的64个单位,首次用桃蚜证明了抗药性基因的扩增。
在1986年莫切斯(C.Mouches)等将Tem-R(抗双硫磷)库蚊品系中的酯酶B1在噬菌体λgt11中克隆,结果发现此酯酶的基因拷贝数至少是敏感品系的250倍,进一步证明杀虫剂筛选可引起编码解毒酶的基因扩增。
抗药性调控机制 与抗药性有关的突变基因既可是结构基因,也可是调节基因。有些结构基因的产物可以是酶、受体,或核糖体、微管蛋白等其他细胞组分。这些产物一般是药剂的靶标,而结构基因的突变能对基因产物产生一种至关重要的改变,如靶标部位敏感度降低,或增加代谢药剂的能力。调节基因的产物可控制结构基因转录的速率。还可识别并与药剂结合,从而控制对相应解毒酶的诱导作用。
奥本诺思(F.J.Oppenoorth)在1967年认为结构基因负责蛋白质合成的“质”的成分,而调节基因则操纵着蛋白质的“量”的调节。而普拉泼(F.W.Jr.Plapp,1984)认为调节基因存在两类,它们在遗传和生化方面是不同的。一类表现出“全或无”遗传,即完全显性或完全隐性,这类主要涉及合成蛋白质的量的改变,这类基因的调节称为“近”调节基因(“near”regu-lator); 第二类表现为共显性遗传,主要涉及合成蛋白质的性质的改变,即在抗药性昆虫中产生变构形式的解毒酶,这类基因的调节称为“远”调节基因(“distant”regulator)。哈斯特伦(I.P.Hallstron)在1984年认为类同于贾科布—梅诺德(Jacob-Meonod)模式,即有三个基因水平:结构基因、整合基因(integrator)和传感器(sensor)。上述的远调节基因相当于传感器主要是通过识别杀虫剂之类的外部信号起作用的。
抗药性群体遗传学 主要研究抗药性种群中的遗传结构及其变化规律,应用组建的数学模型和计算机模拟来研究群体中抗药性基因频率、基因型和基因型频率以及影响这些频率的因素,以此来探讨抗药性演化的过程,从而预测抗药性发展的趋势,为制定抗药性治理的策略提供依据。
- 饮食服务业会计是什么意思
- 饮食服务业会计是什么意思
- 饮食服务业技术职称是什么意思
- 饮食服务业收入是什么意思
- 饮食服务业服务质量是什么意思
- 饮食服务业统一核算、分计盈亏是什么意思
- 饮食服务业能力利用指标是什么意思
- 饮食服务业营业收入是什么意思
- 饮食服务业营业收入明细帐是什么意思
- 饮食服务业费用是什么意思
- 饮食服务公司是什么意思
- 饮食服务机械是什么意思
- 饮食毛利率是什么意思
- 饮食治大病是什么意思
- 饮食男女是什么意思
- 饮食男女是什么意思
- 饮食男女,人之大欲存焉是什么意思
- 饮食疗法是什么意思
- 饮食疗法是什么意思
- 饮食疗法是什么意思
- 饮食疗法是什么意思
- 饮食疗法是什么意思
- 饮食禁忌是什么意思
- 饮食禁忌是什么意思
- 饮食管理制度是什么意思
- 饮食考 清 徐养源是什么意思
- 饮食营养是什么意思
- 饮食营养大全是什么意思
- 饮食规则是什么意思
- 饮食起居是什么意思
- 饮食起居是什么意思
- 饮食须知是什么意思
- 饮食须知是什么意思
- 饮馔服食笺是什么意思
- 饮馔服食笺是什么意思
- 饮馔服食谱是什么意思
- 饮马于河是什么意思
- 饮马投钱是什么意思
- 饮马投钱是什么意思
- 饮马歌是什么意思
- 饮马流花河是什么意思
- 饮马漓江边是什么意思
- 饮马长城窟行是什么意思
- 饮马长城窟行是什么意思
- 饮马长城窟行是什么意思
- 饮鸩止渴是什么意思
- 饮鸩止渴是什么意思
- 饮鸩止渴是什么意思
- 饮鸩止渴是什么意思
- 饮鸩止渴是什么意思
- 饮鸩止渴(止渴饮鸩)是什么意思
- 饯是什么意思
- 饯是什么意思
- 饯彦九郞还日本诗轴是什么意思
- 饰是什么意思
- 饰是什么意思
- 饰是什么意思
- 饰心无彩缋,到骨是风尘是什么意思
- 饰情矫行是什么意思
- 饰纹姬蛙(Microhyla ornata)是什么意思