基因突变gene mutation
DNA分子上碱基顺序改变,导致基因型和表型产生变化的遗传过程,也称点突变。突变的最小单位是突变子。一个基因可以包含许多突变单位。同一基因位点上突变产生的基因,称为等位基因,等位基因多于二个时称为复等位基因。
在生物群体中,突变发生的个体和时间以及突变的方向都是随机的。绝大多数生物的基因突变率很低。在有性生殖的生物中,突变率是指每一配子发生突变的概率,用一定数目配子中的突变型配子数表示。例如,果蝇红眼(W)突变为白眼(W)的突变率为4×10-5。在无性生殖的细菌中,突变率是指每一细胞世代中每一细菌发生突变的概率,用一定数目的细菌在分裂一次过程中发生突变的次数表示。例如,大肠杆菌乳糖发酵基因由lac突变为lac+的突变率为2×10-7。基因突变具有可逆性,野生型基因经过突变成为突变型基因的过程称为正向突变。突变基因通过突变而成为野生型基因称为回复突变。
现代基因突变的研究是从1910年开始的。摩尔根在红色复眼果蝇中发现白色复眼果蝇,这是自发产生的突变。1927年,马勒(H.J.Muller)报告了X-射线诱发果蝇发生突变,第一个发现了人工诱发突变。1947年,奥尔巴克(C.Auerbach)首次使用化学诱变剂,用氮芥诱发了果蝇的突变。1959年,弗里兹(E.Freese)提出基因突变的碱基置换理论,1961年,克里克 (F.H.C. Crick)等提出移码突变理论。
类别 所有的基因突变类型,都是在DNA分子结构中由于碱基的变化所致。根据导致突变的来源,分自发突变和人工诱发突变,二者产生的突变型没有本质上的不同,诱变剂的作用只是提高基因的突变率。按照基因结构改变的类型,可分为碱基置换突变和移码突变。碱基置换突变指DNA分子上一对碱基改变而引起的突变。一种嘌呤被另一种嘌呤取代,或一种嘧啶被另一种嘧啶取代叫做转换。一个嘌呤被一个嘧啶替代,或一种嘧啶被一个嘌呤替代叫做颠换。移码突变指DNA分子上一对或少数几对相连的核苷酸增加或减少,导致这一位置以后一系列编码发生移位而产生的突变。故移码突变是由于碱基缺失或插入引起的。
按照遗传信息的改变方式,突变又可分为同义突变、错义突变和无义突变。同义突变指基因上的密码序列改变,而基因终产物蛋白质上氨基酸序列没有改变,这与密码子的简并性有关。例如GUC(缬氨酸)变为GUA仍为缬氨酸,就是同义突变。错义突变指一对碱基改变而使某一氨基酸变为另一种氨基酸。例如GUC(缬氨酸)变为GCC(丙氨酸)。有些错义突变会严重影响蛋白质的活性甚至完全失活。无义突变是指一对碱基的改变,而使氨基酸的密码子变为终止密码子。例如AAG(赖氨酸)突变为UAG(终止密码子)。无义突变使蛋白质合成提前终止,产生的不完全肽链一般是没有活性的。
机理 至今大多数的突变机理尚不清楚。一般利用诱变手段研究突变机理。碱基类似物分子结构与天然的碱基化合物非常相似,因而在一些主要的反应中能够取代碱基,诱发基因突变。5-溴尿嘧啶(Bu),是胸腺嘧啶(T)的类似物。一旦Bu取代了T,就会导致A-T→G-C的转换。2-氨基嘌呤(AP)是嘌呤的类似物,它掺入到DNA分子中能够诱发A-T→G-C和G-C→A-T两种转换。一些药物能诱发DNA结构改变,例如羟胺、亚硝酸和烷化剂等能够改变核酸的结构,其诱变效应取决于DNA复制,羟胺能和胞嘧啶专一性地反应,诱发G-C→A-T的转换。亚硝酸使鸟嘌呤、腺嘌呤和胞嘧啶脱氨基分别形成黄嘌呤、次黄嘌呤和尿嘧啶。后两种碱基的变化导致G-C→A-T或者A-T→G-C的转换。而黄嘌呤是一种无义的碱基,它出现在DNA分子中只能导致细胞死亡。烷化剂能够同DNA分子的许多部位发生作用,使DNA分子增加了烷基侧链,导致突变。吖啶类染料分子能够嵌入DNA分子中,从而使DNA复制发生差错而造成移码突变。
物理诱变因素能诱发突变,常用的有紫外线、X-射线、同位素、激光和超声波等。应用最多的是紫外线,紫外线导致突变的有效波长范围是200~300nm,尤以260nm左右效果最好。紫外线照射能引起DNA分子断裂、DNA分子双链交联以及形成胸腺嘧啶二聚体T-T,使DNA双链呈现不正常的构型。
意义 基因突变在自然界广泛地存在,是遗传变异、生物进化的根源。水稻的矮生型、棉花的短果枝、谷类的糯性胚乳等性状,都是基因突变的结果。碱基置换突变常导致蛋白质中一个氨基酸的改变,例如正常血红蛋白(Hb)第6位的谷氨酸改变为缬氨酸,便成为镰刀形细胞贫血症的血红蛋白(HbS),所对应的碱基变化是GAA(谷氨酸)→GUA(缬氨酸)。用各种物理的或化学的手段对生物诱变,可产生大量多样的突变体,选择有利用价值的突变体可育成优良品种,用γ-射线和甲基磺酸乙酯(EMS)处理是最常用的诱变手段。
基因突变
又称为点突变,它是DNA分子链上碱基组成或排列顺序的变化。基因突变分自发突变和诱发突变两种。自发突变是在DNA复制、修复、重组的过程中,在自然条件下发生的突变。主要包括碱基替换和移码突变。碱基替换是指构成基因的DNA分子中,一个碱基对被另一碱基对替换。其中有嘌呤之间,或嘧啶之间的替换称为转换;嘌呤和嘧啶的相对位置发生改变,嘌呤换成嘧啶,嘧啶换成嘌呤为颠换。移码突变是指在DNA分子中,由于增加或减少一个或几个碱基对而引起的突变。这一增加或者减少的结果是使整个遗传密码发生次序紊乱。人体内自发突变多为移码突变,如体内修复功能正常,则突变后会恢复正常。诱发突变是特定环境因素,多为人工诱变因素引起的基因突变。例如X、γ射线、化学物质、药物等都可以引起突变。某些化学物质与某种碱基结构相似,则可发生碱基替换或插入到DNA分子序列中去。射线可直接作用DNA大分子,而发生突变。当基因突变扰乱了生物(包括人类)遗传基础的平衡,使蛋白质合成发生质的变化,就会产生表型效应。形成形态上的缺陷或生理功能上的缺陷。人类各种遗传病都是在进化过程中经突变而产生的。如果只是体细胞发生突变,这种突变尽管可以形成某些缺陷,但是不会遗传给子代的;如果是性细胞发生突变,形成的某些缺陷是会遗传给子代的。
基因突变jiyin tubian
基因从一种状态转到另一状态时所发生的分子结构的变化,又叫点突变。分子结构的变化可涉及DNA中碱基对的缺失、增加或代换。基因突变对表现型的影响,可分成如下几类:(1)形态突变型:如普通绵羊经突变产生的短腿安康羊,外形的变化可从表现型识别。(2)生化突变型:突变可使一个特定的生化功能丧失,常见的是营养缺陷型。如红色面色霉一般能在基本培养基上生长,但突变后,要在基本培养基上加上某种特定氨基酸才能生长。(3)致死突变型:突变导致个体死亡或生活力明显下降,如人类血友病和植物白化病都为致死突变。(4)条件致死突变型:在一定条件下致死,而在另一条件下成活的突变。如噬菌体T4的温度敏感,突变型在25℃能在大肠杆菌上生长,形成噬菌斑,但在42℃为致死。基因突变的一般规律或特点表现在:(1)多向性:如人类的ABO血型是由IA、IB和i3个复等位基因组成的。(2)可逆性:显性基因可突变成隐性基因(正突变),有时隐性基因也可变成原来的显性基因(逆突变)。突变可逆性表明,基因突变不是遗传物质的丧失,而是遗传物质的改变。(3)有害性:因生物现有性状是经长期进化形成的遗传平衡系统,基因突变会破坏这种平衡,不利于生长发育。例如,人类遗传病基本上是由基因突变引起的。当然,有害不是绝对的,在一定条件下反为有利。如雄性不育突变,对生物本身有害,但在杂交优势利用上,可免去人工去雄,提高杂交制种效率。(4)重演性:这是指同 一物种或近缘物种个体间可发生相似的基因突变。例如,大麦、小麦等经辐射后,都有相似的突变性状,如 一定比例的早熟个体和矮生个体。由于物种或近缘物种的基因有许多是同源的,使得基因突变具有重演性。
基因突变gene mutation
又叫点突变,系指在染色体上DNA分子结构中碱基的变化而言。基因改变包括基本数目、位置的改变,可使遗传功能发生改变。如结构基因发生突变可改变蛋白质上氨基酸的排列,改变DNA上的密码使蛋白质合成不能或在蛋白质的免疫性、化学性和酶的活性上发生改变而引起疾病,如镰状红细胞性贫血就是在血红蛋白上的1个氨基酸发生了替换而引起。控制基因发生突变使合成mRNA(信使核糖核酸)的能力降低或使核糖体上mRNA的信息密码不清楚或造成信息密码的翻译功能障碍而使蛋白合成减少、缺乏、改变性质而产生疾病。突变基因在遗传中有两种表现形式。一种由显性等位基因(这些基因由于在DNA中的功能位置决定其有外显力),可使子代显示病态,发生显性遗传。此种等位基因如位于常染色体上则称为常染色体显性遗传,如位于性染色体上则称为性连显性遗传。另一种突变基因称病态等位基因(亦是由于基因在DNA上的位置决定的),在遗传给子代时不显示病理状态,只以1个遗传痕迹(或基因)——即1个病态等位基因携带者而存在,仅当2个相同的病变等位基因结合时(2个携带者),才显示疾病状态,此为隐性遗传。若病态等位基因位于常染色体上则称为常染色体隐性遗传,若位于性染色体上则称为性连隐性遗传,如带有相同的病态等位基因的2人结婚时,则子代可发生显性遗传性疾病。
基因突变
又称“点突变”。单一基因内任何可遗传的结构的改变。按基因结构改变的类型,分碱基置换、移码、缺失和插入四种;按遗传信息的改变方式,分错义和无义两类。
基因突变
即点突变。
基因突变
突变一词最先由de Vries在本世纪初提出。基因突变是指突然出现的由基因结构改变引起的可遗传的变异。以后证明,de Vries在月见草(Oenothera lamarc-kiana)中所看到的突变,大部分是染色体畸变,只有极小一部分是基因突变。广义的突变包括染色体畸变,而狭义的突变则专指基因突变,也常称点突变。如果不加说明,一般均指核基因结构的改变。突变是一个过程,通过突变而出现的基因称为突变基因。所谓突变型是指具有突变基因的个体或细胞。不经人工处理而发生的突变称为自发突变; 经人工处理而发生的突变称为诱发突变。能够诱发基因突变的化学药品和物理因素统称为诱变剂。任何一种生物都发生自发突变,也都能通过人工处理而发生诱发突变。任何生物的遗传物质都是核酸,而且所有生物的遗传密码都相同,所以,所有生物的基因突变具有共同特性。
基因突变的共同特点
(1) 稀有性:不论是人、细菌或是噬菌体,突变都是一个稀有的事件。一个突变型细胞经过一次分裂产生两个突变型细胞。在高等动物中,二个突变型精子可以来自两个也可以来自一个精母细胞,因此准确地估计突变率主要来自组织培养中的细胞。每一细胞在每一次细胞分裂中发生突变的机率是: 组织培养中的人骨髓细胞的抗8-氮鸟嘌呤突变是7×10-4,抗8-氮鸟核苷突变是1×10-6,大肠杆菌的乳糖发酵缺陷型突变是2×10-7,抗噬菌体T1突变是2×10-8,组氨酸缺陷型突变是2×10-6。其他生物的各种突变同样也都有极低的突变率。
(2) 随机性:突变的发生,不论是对于个体、细胞、基因来讲,或对于所影响的性状来讲都是一个随机的事件。在一系列试管中接种一定量不含突变型的细菌,经过一定时间的培养以后测定每一试管中的某一特定突变型的细菌数,可以看到这些试管中突变型细菌数的分布符合于按照随机分布所预测的情况。这一实验结果说明基因突变对个体来讲是随机的。如果不是观察一个基因的突变而是观察两个基因的突变,许多实验都说明,二个突变发生在同一细胞中的机率是二个基因分别发生突变的机率的乘积,这说明对于基因来讲,突变也是随机的。细菌培养物接触某种抗生素后,常出现对于这一药物呈抗性的突变型细菌。这似乎说明对于所影响的性状来讲,突变似乎不是随机的。可是彷徨实验、重新涂布实验和影印培养实验结果都有力地说明,抗药性突变多数发生在接触药品以前,因此同样说明对于性状来讲,突变也是随机的。
(3) 可逆性:突变是可逆的,也就是说,突变型会发生回复突变。例如突变型白眼果蝇可以发生另一突变,回复到正常红眼果蝇。从进化论观点看来,这是不足为奇的,因为很难说哪一个野生型基因不是突变型基因。实际上这不过意味着一个基因可以许多不同的稳定的状态存在着。
(4) 一个突变是DNA的一定位置某一结构改变。这种结构改变可以有多种,也就是说,不同性质的结构改变可以使同一基因发生突变,并且造成同一表型的变化。造成基因突变的结构改变主要是碱基替代、移码、插入,缺失。
❶碱基替代:是真正的点突变,因为它只涉及一对碱基的改变。组成DNA的碱基有腺嘌呤(A)、鸟嘌呤(G)胸腺嘧啶(T)和胞嘧啶(C)四种,如果一个嘧啶为一个嘌呤所替代,或一个嘌呤为一个嘧啶所替代,这种替代称为颠换。如果一个嘧啶为另一个嘧啶所替代,或一个嘌呤为另一个嘌呤所替代,这种替代称为转换。图1中的直线箭头表示转换,断线表示颠换。例如人的镰形细胞贫血症患者的血红蛋白β链的第六位氨基酸如果由正常的谷氨酸(GAG)变为缬氨酸(GUG),这
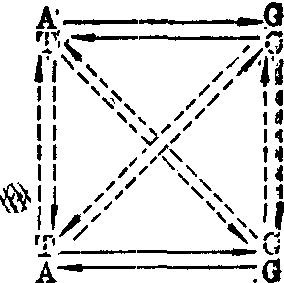
图1 碱基替换
就是说,在DNA分子上的相应的碱基由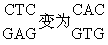 在这一例子中,中间一对碱基发生了由TA转变为AT这一颠换。
在这一例子中,中间一对碱基发生了由TA转变为AT这一颠换。❷移码:是指这种改变涉及一对或少数几对(除了三对)碱基。一对或几对碱基的缺失或重复将会改变该点后面的碱基排列顺序,从而形成错误的密码,并转译成为不正常的氨基酸。例如人的Hb Wayne(一种异常血红蛋白)就是由于α链基因发生移码突变的结果。Hb Wayne是由于α链的第139密码子(AAA)最后的碱基(A)丢失,因而在读码时,将第140密码子(UAC)的第一个碱基(U)移前,读成AAU编码门冬酰胺。然后再往后读出ACC、GUU、AAG、CUG、GAG、CCU、CGG7个密码子后才遇到UAG (终止密码),合成才告结束。

❸插入:是指一段核苷酸顺序插入某一基因,也可以使这一基因发生突变。大肠杆菌的一种温和噬菌体Mu-1能够整合到宿主细菌的染色体的许多位置上而使它成为溶源性细菌。每一整合可以使一个基因发生突变。在大肠杆菌中还发现所谓插入顺序和转座子。插入顺序是指能在质粒与质粒之间,或质粒与染色体之间转移位置的一小段DNA,但不含有任何基因。转座子也是指能在质粒与质粒之间或质粒与染色体之间转移位置的一小段DNA,但其中包括了在两端的插入顺序和夹在当中的某些基因。当这些能够转移位置的插入顺序或转座子转移到一个基因中时,就引起这一基因的突变。
❹缺失:是指某一基因内部一段核苷酸的缺失,往往也会引起突变。(5) 基因突变带来一定的表型效应: 表型改变决定于发生突变的基因,而不决定于这一基因发生什么结构改变。这是因为基因的基本功能在于编码蛋白质。各种不同的DNA结构改变虽然使蛋白质发生不同的结构改变,但是只要这一改变使蛋白质失去它应有的功能,那么它们都会带来同一表型效应。不过,如果着眼于突变所造成的遗传密码意义的改变,那么DNA结构改变所造成的改变可以有三种,即错义密码子、无义密码子和同义密码子。某一对碱基的置换使编码某一氨基酸的三联体密码子变成编码另一氨基酸的密码子时,后一密码子称为错义密码子。例如原来编码亮氨酸的密码子UUG变成编码苯丙氨酸的密码子UUU。如果某一对碱基的置换使编码某一氨基酸的密码子变成编码终止信号的密码子时,后一密码子称为无义密码子。例如上述密码子变成终止密码子UAG。如果密码子虽然改变,然而所编码的氨基酸还是原来的氨基酸,那么这一密码子称为同义密码子。例如UUG变为UUA,这两个密码子都编码亮氨酸(见“遗传信息的转录”)。此外虽然密码子的改变造成所编码的氨基酸的改变,可是如果这一改变不影响这一蛋白质的活性,那么这一改变同样不表现任何表型反应。
由于突变往往使一种蛋白失去原有的活性,所以突变基因常是隐性的,而且在一般情况下不利于生存。一切基因突变都具有上述这些特点,诱发突变和自发突变所不同的只在于突变率的高低,此外没有任何区别。
诱发突变的分子机理 诱发突变早在1927年由Mul-ler在果蝇中发现,可是对突变的分子机理的了解主要在五十年代通过微生物中的化学药品诱变的研究而取得。诱发碱基替代的化学药品有5-溴尿嘧啶等天然碱基结构类似物,亚硝酸、羟胺、氮芥和N-甲基-N′硝基-N亚硝基胍等烷化剂。物理因素如X射线和紫外线等也能诱发碱基替代,可是对于它们的作用机理则了解得较少。5-溴尿嘧啶是胸腺嘧啶的结构类似物,所以可取代胸腺嘧啶而参入DNA中。在DNA双链分子中,胸腺嘧啶(T)和腺嘌呤(A)构成一双碱基(AT)。5-溴尿嘧啶取代胸腺嘧啶后和腺嘌呤构成一对碱基(ABU)。
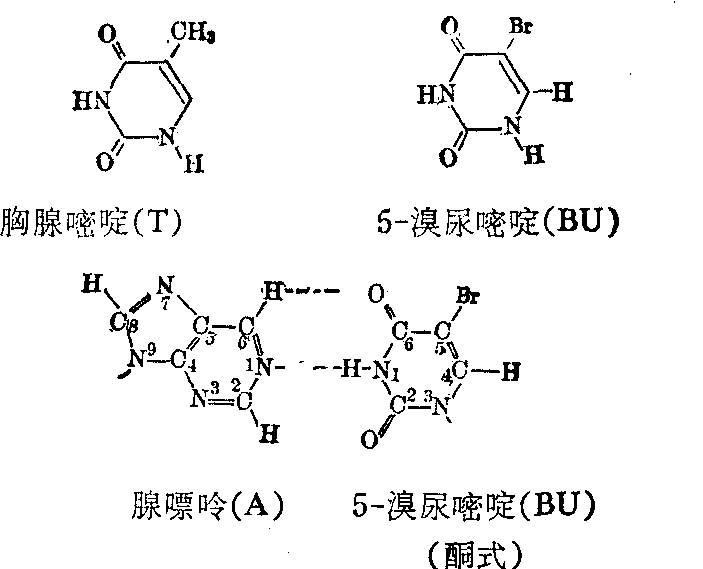
胸腺嘧啶和5-溴尿嘧啶都有酮式和烯醇式两种互变异构形式,而且都以酮式占绝对优势。可是,5位上的溴原子的存在使5-溴尿嘧啶较多地以烯醇式出现。当DNA复制到达ABU碱基对时,5-溴尿嘧啶刚好以烯醇式出现,它便在新生DNA链中和鸟嘌呤配对。
再经过一次DNA复制时,就出现GC碱基对。原来的AT碱基对便转换成为GC。从BU参入DNA到完成AT→GC这一转换过程是:
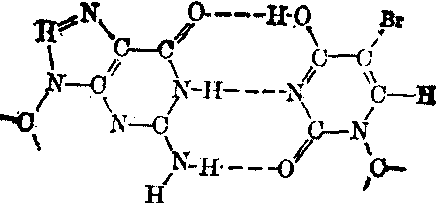
鸟嘌呤(G) 5-溴尿嘧啶(BU)(烯醇式)
如果5-溴尿嘧啶参入DNA时呈烯醇式,那么参入时它不是取代胸腺嘧啶而是取代了胞嘧啶。如果在参入以后的复制中,它呈酮式,那么在新生DNA链中出现ABU对,然后再通过一次复制时便转换成为AT对。
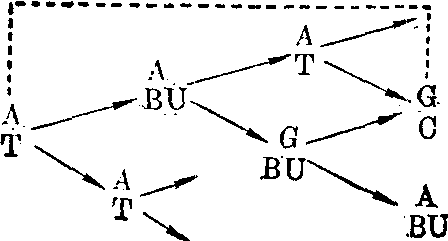
AT转变为GC
亚硝酸对于碱基起氧化脱氨作用。它使腺嘌呤(A)转变为次黄嘌呤(H),使胞嘧啶(C)转变为尿嘧啶(U)。因此同样地能够诱发
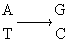
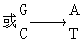 这两种转换。
这两种转换。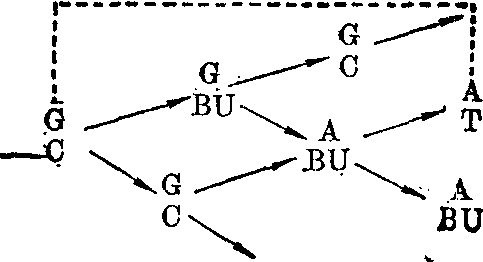
GC转变为AT
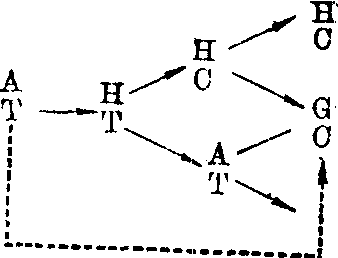
作用于腺嘌呤,引起AT→GC转换
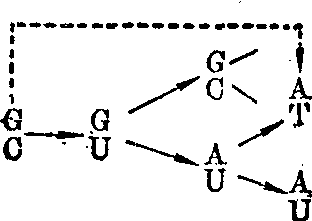
作用于胞嘧啶,引起GC→AT转换
羟胺几乎只和胞嘧啶发生反应,所以不同于5-溴尿嘧啶和亚硝酸,它几乎只诱发GC→AT这一转换。诱发移码突变的诱变剂主要是吖啶类染料以及它的衍生物,如ICR-191等。
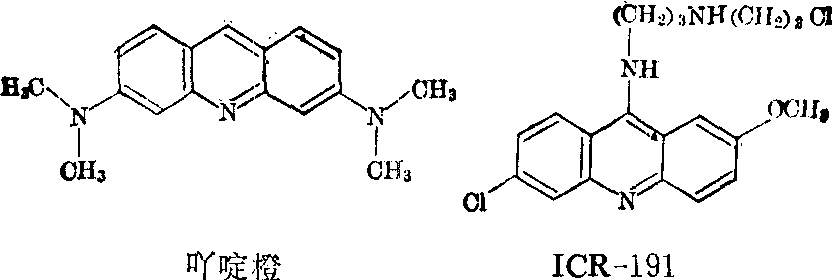
这类化合物能够嵌入DNA双螺旋中,一般认为这一嵌入使DNA复制过程中出现个别碱基对的缺失或重复,这便是它们诱发移码突变的主要机理。
紫外线的主要作用是促使DNA分子上两个邻接的碱基转变成为二聚体,特别是胸腺嘧啶二聚体。虽然许多事实都说明二聚体的形
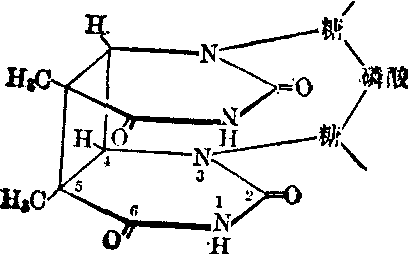
胸腺嘧啶二聚体
成和紫外线的诱变作用有关,可是它的分子机理还不清楚。紫外线所诱发的突变既有替代突变,也有移码突变。电离辐射的诱变机理尚不很了解。它们的一部分作用是间接的,那就是由电离辐射产生自由基,这些自由基再作用于DNA而造成突变。它们的另一部分作用是直接的,那就是由电离直接造成一些化学键的断裂,所以电离辐射常诱发染色体畸变。但是电离辐射同样能诱发点突变。X射线所产生的一些突变型也能为X射线诱发而回复成为野生型。
缺失可以造成基因突变。电离辐射可以由于它的电离作用而诱发缺失。许多烷化剂被称为拟辐射物质,因为它们具有类似于电离辐射的一些生物学效应,包括诱发缺失和其他染色体结构畸变在内。可是在沙门菌中曾证明,最有效的缺失突变诱发剂不是快中子、紫外线、氮芥、亚硝酸胍,而是亚硝酸。它的诱发作用的分子机理还不清楚。
突变诱发过程:诱变剂作用于核酸而诱发突变,所以能在高等生物中诱发基因突变的化学药品也能在微生物中诱发基因突变。但是诱变剂在接触核酸以前先得通过细胞表面和细胞质。诱变剂对于核酸引起了损伤或变化以后,必须通过DNA复制才造成突变基因所具有的结构。在这中间,细胞会对这些损伤部分进行修复。凡此种种都影响着基因突变的发生。而且基因突变发生以后,这一细胞并不一定立即表现突变型性状,这中间还有一个过程。例如各种生物的DNA对于紫外线的敏感性并没有显著的差别,可是紫外线对于高等植物的诱变力较弱,这主要是由于细胞表面穿透性的不同所造成的。沙门菌的深度粗糙突变型对于各种诱变药物的敏感性都高出于野生型大约一个数量级。这是由于这突变型的细胞表面的脂多糖发生了变化,因而使得诱变剂更容易进入细胞的缘故。许多物质在进入细胞后,受到细胞中某些酶的作用而发生变化。例如陆蒽酮本身对于沙门菌没有诱变作用,可是在体外经过肝脏抽提物作用后,便转变为具有诱变作用的海蒽酮。无疑地像陆蒽酮这样的物质在未经动物试验以前,就应该认为是危险的药物。相反地,一些诱变剂可能在细菌中遭到破坏而降低它们的诱变作用。
陆蒽酮
海蒽酮
羟胺几乎只和胞嘧啶发生化学反应,所以对于游离的噬菌体来讲,它只诱变GC→A T这一转换。可是在细胞中它和一些物质发生反应而产生过氧化氢,而过氧化氢是一种没有特异性的诱变剂,所以对于一般细胞来讲,羟胺的诱变作用的特异性就不那么强。电离辐射部分地由于它在细胞中产生自由基而诱发突变,而自由基的产生和氧的存在有关,所以细胞中的氧分压对于X射线的诱变作用有一定的影响。
诱变剂造成DNA损伤以后的修复作用有多种。以紫外线所造成的二聚体的修复为例,有光复活、切补修复和重组修复。许多生物的细胞中有为可见光所活化的光复活酶,它能分解胸腺嘧啶二聚体,从而消除潜在的基因突变。切补修复在四种酶的协同作用下进行,这四种酶是核酸内切酶、核酸外切酶、DNA聚合酶I和DNA连接酶。二聚体核酸内切酶和核酸外切酶切除,DNA聚合酶进行修补合成,最后在DNA连接酶的作用下完成修补作用。在大肠杆菌中曾经获得对于紫外线格外敏感的突变型。它们的切除酶系发生了缺陷。在着色性干皮病患者的皮肤成纤维细胞培养物中,可以看到核酸内切酶发生缺陷。这种病人对于紫外线格外敏感,接触日光后容易发生皮肤癌。没有光复活酶和切补酶系所消除二聚体的DNA还能由于重组而修复。DNA损伤的修复无疑地具有消除潜在的基因突变的作用,但是,某些修复过程本身也会导致差错而造成基因突变。发生了基因突变的细胞一般并不立即表现为突变型,这一现象称为表型迟延。表型迟延的原因有二,即分离性迟延和生理性迟延。发生突变的细胞一方面要通过分离而出现纯合细胞,另一方面原有的酶稀释和破坏以后才能有突变型表现。
自发突变的原因 研究诱变的机理有助于我们对于自发突变的了解。如果把诱发基因突变数和辐射剂量的关系作图,往往可以看到两者呈直线关系,而且这一直线通过原点,说明任何低剂量的辐射可以诱发突变。因此一部分自发突变可能是地球表面的宇宙线、紫外线等所诱发的。生物还可能不自觉地接触环境中的诱变物质,从而被诱发基因突变。这些自发突变实际上是诱发突变。许多生物可产生诱变物质,在储藏长久的洋葱种子中曾经得到具有诱变作用的抽提物。许多生物产生咖啡碱、重氮丝氨酸等具有诱变作用的物质。过氧化氢是一种诱变剂,也是一种正常代谢产物。因此,一部分自发突变可能是自身所产生的诱变物质所诱发的突变,对于细胞来讲它们是自发突变,对于基因来讲则是诱发突变。从诱发突变的研究中可以看到5-溴尿嘧啶通过互变异构效应而诱发基因突变。胸腺嘧啶本身也有两种互变异构形式。可以推想在无5-溴尿嘧啶参入的情况下,由于碱基本身的互变异构效应,同样可以导致碱基替代。这是真正意义的自发突变。计算结果说明,由互变异构所预测的自发突变率符合于实际的自发突变率。
诱变剂的检测 关于癌症病因的一个假设认为,癌细胞来源于体细胞基因突变。如果这一假设反映了客观实际,那么应该预期致癌物都是诱变剂。用Ames所首创的检验方法进行检测的结果,阳性符合率达到95%,所以诱变剂的检测对癌症预防具有非常重要的意义。基因突变的检测方法主要有以下几种:
(1) Ames法: 用一组沙门菌组氨酸缺陷型的回复突变作为检测指标。这一系列缺陷型的基因突变的本质是已知的,某些菌株是替代类型,某些是移码类型。除了组氨酸缺陷型以外,它们都是切补修复和深度粗糙突变型,所以它们对于诱变剂格外敏感。把待测细菌满铺在培养皿中的基本培养基上, 在培养皿的中央放上少量待测物质。经过培养以后观察是否出现一圈菌落,而判断所测物质是否具有诱变作用以及诱变作用的强弱。如果待测物质没有诱变的作用,则缺陷型细菌不能形成菌落。如果待测物质具有诱变作用,则被诱发发生回复突变的细菌便能形成菌落。诱变作用愈强,则诱发的回复突变愈多,出现的菌落也愈多。此外在培养基中还可以加上大鼠肝脏抽提物,以检测本身虽然没有诱变作用,可是进入人体后能转变为诱变剂的物质。这一方法的优点是快速、简便、费用小,所以是被广泛采用的一种方法。不足之处是待测细胞是细菌而不是动物细胞。
(2) 扩散小室法:把待测细胞放在小的塑料容器中,把这小室埋藏在动物体内,将待测物质注入动物,使待测细胞在活体中接触待测物质。接触一定时间以后,取出小室的细胞,测定有多少细胞发生了突变。在这一方法中,可以用细菌作为待测细胞,也可以用人体细胞作为待测细胞。
(3) 显性致死突变法:用待测物质饲养雄性小鼠,然后使它们和未处理的雌鼠交配,死胎数便是由待测物质所诱发的显性致死突变数。这一方法可以用来测定通过食物和其他途径长期接触的物质的诱变作用。不足之处是费用较大,需要时间较长,而且不易区分待测物质的诱变作用和引起畸胎的作用。
(4) 隐性致死突变法:用待测物质处理雄性小鼠,然后使它们和带有同样隐性突变型标记的雌性小鼠交配。子代中出现具有这些表型的个体的多少就表示待测物质诱变作用的强弱。这一方法除了具有显性致死突变法所有的优点以外,并且可以确实知道所测定的是待测物质的诱变作用而不是别的作用。不足之处是费用大而时间长,而且必须具备一定品系的小鼠。
胞质基因突变 胞质基因和核基因一样,会自发突变,也会被诱发发生突变。胞质基因突变和核基因突变一样,具有稀有、随机、可逆等特性,而且每一个突变反映胞质基因组的一定位置的某一结构改变,也带来一定的表型效应。胞质基因突变在下述三个方面不同于核基因突变:
❶胞质基因突变的表型效应不象核基因突变那样广泛。这是因为胞质基因组远较核基因组为小,它们所编码的蛋白质也远较核基因所编码的为少。叶绿体和线粒体中有不同于细胞质中的核糖体,这上面的核糖体蛋白质一部分由叶绿体和线粒体基因本身所编码。已经知道许多抗生素抗性突变是核糖体蛋白基因发生突变的结果。此外,也由于抗药性突变最容易被察觉,所以在胞质基因方面发现得最多的是抗药性突变。线粒体蛋白质中包含着和氧化磷酸化有关的酶。酵母菌的营养性小菌落突变型是线粒体基因突变的结果,它发生缺陷的是一部分细胞色素成份。总起来讲,胞质基因突变的表型效应不是那么广泛。此外由于叶绿体和线粒体的某些成份为胞质基因和核基因所共同编码,所以同一表型效应也可以是核基因突变的结果。例如酵母菌的分离性小菌落。
❷胞质基因突变的诱发频率特别高。例如用吖啶黄处理酵母菌可以得到几乎100%的小菌落突变,同样现象也曾在构巢曲霉中报道。
❸从诱变剂来讲,胞质基因和核基因有相同之处,也有不同之处。紫外线、亚硝基胍、亚硝基甲替尿烷等有效的核基因诱变剂都能诱发小菌落突变,可是亚硝酸和X射线却未见有报道。更为特殊的是链霉素对于核基因不是一种诱变剂,可是对于莱因哈德衣藻(Chlamydomonas reinhardi)的叶绿体基因来讲,却是一种有效的诱变剂,它能够诱发链霉素抗性突变,也能诱发其他突变。总之,对于胞质基因突变的了解还远远不及对于核基因突变的了解。
基因突变gene mutation
基因在一定外界因素影响下,碱基的结构发生改变。
基因突变
gene mutation
基因突变
gene mutation
基因突变
gene mutation
基因突变
gene mutation
基因突变gene mutation
由一些非属于正规的孟德尔组合因素而引起的遗传物质的跳跃式变化。突变是遗传物质的一部分(即它们属于遗传型的),虽然它们对个别机体表现型的影响可能观察不到。大多数突变涉及到个别的基因,当然也可能会涉及到许多基因,引起染色体的总的改变。突变也可能发生在细胞体内(称体细胞突变),并通过该细胞的有丝分裂进行传递。从某种突变对个体适应价值的观点来看,其结果完全是随机的,它们在进化中的作用是通过自然选择的过程而发生的。一般来说,大的突变对有机体是有害的,故不可能继续下去;而小的突变,根据一般的观点认为对进化是有利的。
基因突变gene mutation
脱氧核糖核酸分子中发生碱基对的增添、缺失或改变,而引起的基因结构的改变。广义包括染色体畸变。狭义指点突变。通常发生在脱氧核糖核酸复制时期,即细胞分裂间期,包括有丝分裂间期和减数分裂间期。和脱氧核糖核酸的复制、脱氧核糖核酸损伤修复、癌变和衰老都有关系。是生物进化的重要因素之一,研究基因突变除了本身的理论意义外,还有广泛的生物学意义。
- 净净条条是什么意思
- 净净的是什么意思
- 净几明窗是什么意思
- 净几横琴晓寒,梅花落在弦间。 我欲清吟无句,转烦门外青山。是什么意思
- 净出口是什么意思
- 净出口与净进口是什么意思
- 净出口国是什么意思
- 净出口额是什么意思
- 净出鬼是什么意思
- 净出鬼点子是什么意思
- 净出(进)口是什么意思
- 净初级生产能力是什么意思
- 净利是什么意思
- 净利差是什么意思
- 净利差率是什么意思
- 净利息是什么意思
- 净利润是什么意思
- 净利润投标是什么意思
- 净利润率是什么意思
- 净别扭是什么意思
- 净到是什么意思
- 净制是什么意思
- 净刷是什么意思
- 净刹是什么意思
- 净剩余是什么意思
- 净剩分是什么意思
- 净办是什么意思
- 净功率是什么意思
- 净化是什么意思
- 净化中心站是什么意思
- 净化剂是什么意思
- 净化剂净化是什么意思
- 净化器是什么意思
- 净化塔是什么意思
- 净化室内空气是什么意思
- 净化工作台是什么意思
- 净化思想是什么意思
- 净化效应是什么意思
- 净化水是什么意思
- 净化污水是什么意思
- 净化空气运动是什么意思
- 净化空调系统是什么意思
- 净化能力是什么意思
- 净化舞台是什么意思
- 净化药剂是什么意思
- 净化语言是什么意思
- 净化说是什么意思
- 净化身心,清除杂念是什么意思
- 净化饮水是什么意思
- 净半是什么意思
- 净华庵是什么意思
- 净卖臭味是什么意思
- 净去是什么意思
- 净叉是什么意思
- 净叉(差)是什么意思
- 净发须知是什么意思
- 净发鲜披是什么意思
- 净口是什么意思
- 净口业是什么意思
- 净叫介是什么意思