叶绿体yè lǜ tǐ
植物细胞质中的一种细胞器,内含叶绿素等。1918年《植物学大辞典》:“叶绿体,Chlorophyll bodies。”
叶绿体chloroplast
植物绿色细胞所特有的能量转换细胞器。一般呈椭球形,大小约1~3×5~7×2~3μm,有双层被膜与胞质分开,内有片层膜,含叶绿素。叶绿体是植物进行光合作用的场所,自然界有机物质和氧气的供给源。
形态结构和化学组成 大多数高等植物叶绿体呈透镜状。藻类的叶绿体形态多样,有板状、网目状、螺旋状、星状等。细胞中叶绿体的数目依植物种类和组织类型以及发育阶段而异,通常一个叶肉细胞有数十个到一百个左右。叶绿体由叶绿体膜、类囊体和基质三部分构成(图1、2)。
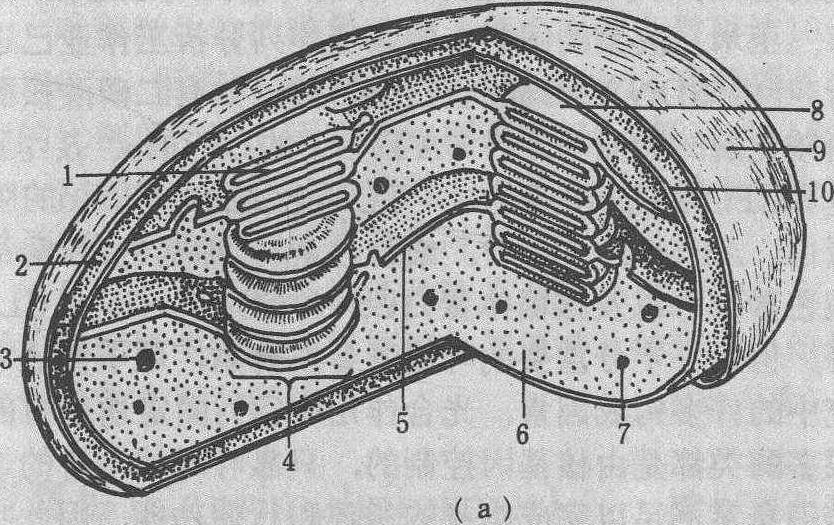
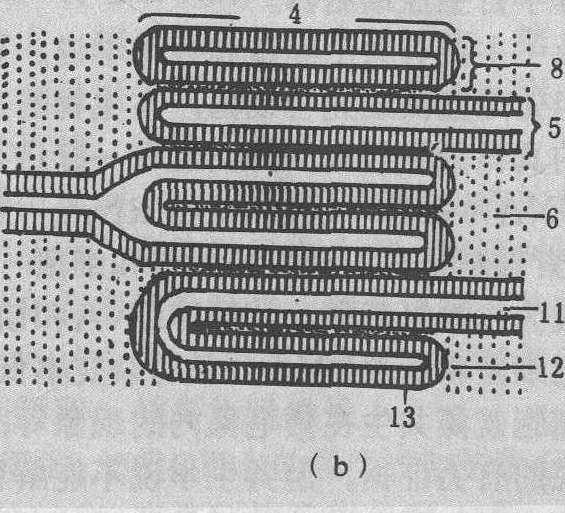
图 1 叶绿体亚显微结构模式图
(a)三维结构; (b)基质类囊体与基粒类囊体部分放大图1. 基粒类囊体间隙; 2. 膜间空间; 3. 淀粉粒; 4.基粒;5. 基质类囊体; 6. 基质;7. 油滴;8. 基粒类囊体;9. 外膜; 10. 内膜; 11. 囊腔; 12. 边缘; 13. 末端基粒膜

图 2 叶绿体电镜图像
叶绿体膜 亦称被膜,由两层单位膜组成,两膜间距为5~10nm。主要成分是蛋白质和脂类,其量为4:1。脂类含量高于类囊体,其中磷脂含量高于糖脂,尤以磷脂酰胆碱含量最高,占磷脂总量的67%。蛋白质中有ATP酶、腺苷酸激酶、糖脂合成的酶系等。叶绿体膜具有选择性透性,3-磷酸甘油酸、磷酸烯醇丙酮酸、磷酸甘油醛、二羧酸和二氧化碳等均可顺利通过叶绿体膜进入基质; ATP、ADP、NADP、糖、有机酸、阳离子、质子等难以通过,光合作用中形成的ATP等则可通过某种穿梭机制进入细胞质。外膜透性强,内膜透性差。物质是否可以顺利地通过主要取决于内膜。
类囊体 由单位膜形成的扁平小囊,是叶绿体的基本结构单位,内含光合作用色素,是进行光能吸收和转化的场所。囊膜厚约7nm,囊腔宽为4~70nm。高等植物的类囊体有两种: 一种较大且彼此不重叠,贯穿在基质中,称基质类囊体; 另一种较小,可自身或与基质类囊体重叠组成基粒,称基粒类囊体。类囊体在叶绿体中的排列方式与植物进化程度有关,植物越低等,两种类囊体的区别越不明显,排列也越不规则。
类囊体膜也和其他膜系统一样,主要成分为脂类和蛋白质。从蓝藻到高等绿色植物等放氧型光合作用生物的类囊体,糖脂是主要脂类,而磷脂较少,这是类囊体膜的一个显著特点。糖脂中主要有单半乳糖甘油二脂、二半乳糖甘油二脂、三半乳糖甘油二脂和硫代奎诺糖基甘油二脂等;磷脂中主要有磷脂酰甘油、磷脂酰胆碱和磷脂酰肌醇等。类囊体膜中全脂肪酸量的80%为亚麻酸等不饱和脂肪酸,从而使膜富于流动性。类囊体膜中还含有叶绿素a、b和类胡萝卜素等色素成分以及质体醌等醌类物质。蛋白质成分中有光合磷酸化偶联因子、NADP-铁氧还蛋白还原酶、铁氧还蛋白等表在蛋白,以及细胞色素、叶绿素-蛋白质复合体、光化学反应中心等内在蛋白,其氨基酸组成以亮氨酸、缬氨酸、苯丙氨酸等非极性氨基酸居多,而半胱氨酸含量很少。
基质 光合作用进行碳同化的场所。主要成分是光合作用中电子传递反应的最终电子受体NADP+、光合磷酸化反应的底物(ADP和磷酸)、蛋白质和脂肪合成的底物和酶及辅酶等。叶绿体本身的遗传系统及蛋白合成系统中的一些因子,如DNA、RNA和核糖体等也存在于此。此外还含有各种离子,其浓度为K+>Ca2+>Mg2+>Na+,调节叶绿体的能量转换和物质代谢,维持叶绿体的构造。
功能 进行光合作用,合成蛋白质和组装核糖体等。
光合作用是绿色植物通过叶绿体将CO2和H2O合成有机物的过程。光合作用色素吸收光能(光子)而被激活后,色素分子内的电子从低能轨道移到高能轨道,于是光能变成光合作用色素的激发态能量。激发态能量通过共振传递方式从吸收短波光的色素分子向吸收长波光的色素分子进行分子间的能量传递,到达反应中心色素(P680和P700),使反应中心色素处于激发态,释放电子给原初电子受体,本身处于氧化态。光系统Ⅱ中的反应中心色素P680接受最终电子供体H2O光解出来的电子而还原,光系统Ⅰ中的反应中心色素P700得到从光系统Ⅱ传来的电子变成还原型,以待接纳光合作用色素再次传来的激发态能量。反应中心色素释放出来的电子经电子传递链到达位于类囊膜外侧的铁氧还蛋白,在铁氧还蛋白-NADP-氧化还原酶的作用下,将电子传给NADP+,使之还原成NADPH。在电子传递过程中,形成类囊体膜内外的H+浓度差,驱动处于类囊体膜外表面的光合磷酸化偶联因子中的ATP酶,利用基质中的ADP和磷酸为底物合成ATP。
通过光合电子传递及光合磷酸化作用所产生的ATP和NADPH中的化学能不稳定,植物进一步利用这些能量还原CO2生成碳水化合物,使能量变得稳定。CO2的同化是一系列酶参与的十分复杂的过程。目前已知高等植物中CO2的同化有3种途径,即C3途径(亦称卡尔文循环)、C4途径和景天酸代谢途径。小麦、水稻等植物进行CO2同化时,生成的最初碳水化合物是一个含3个碳原子的3-磷酸甘油酸,故称C3途径。玉米、甘蔗、高粱等植物除了在维管束鞘叶绿体中有一段C3途径外,在叶肉叶绿体中还有一个C4途径,其CO2受体为磷酸烯醇丙酮酸,生成的最初碳水化合物是含4个碳原子的草酰乙酸。CO2先在叶肉细胞的胞质中与磷酸烯醇丙酮酸结合,由磷酸烯醇丙酮酸羧化酶催化生成草酰乙酸,草酰乙酸还原成苹果酸后进入维管束鞘细胞脱羧放出的CO2进入C3循环,合成糖和淀粉。可见C4途径起收集CO2的作用,碳水化合物的生成由C4和C3两个途径共同完成。在高温、强光和低CO2浓度下,C4植物有较高的光合效率。生长在干燥环境中的景天科、仙人掌科、龙舌兰科和凤梨科等植物,白天气孔一般关闭,晚间气孔张开时,CO2进入叶肉细胞与磷酸烯醇丙酮酸结合,经磷酸烯醇丙酮酸羧化酶催化形成草酰乙酸,再由苹果酸脱氢酶催化形成苹果酸,积累于叶肉细胞的液泡中。白天,苹果酸运至细胞质,经氧化脱羧形成丙酮酸并放出CO2,CO2进入C3途径被还原成碳水化合物。由于此种现象普遍存在于景天科等植物中,故称景天酸代谢途径。此途径基本上与C4途径相似,不同之点是C4途径在空间上将CO2的固定和还原分开,而景天酸代谢途径是在时间上将二者分开。
叶绿体的半自主性 叶绿体和线粒体一样,具有遗传信息系统和蛋白质合成系统,可进行分裂增殖等,表现出一定的自主性。
叶绿体DNA为裸露环状的双链分子,往往以一定的部位与膜连接,长约40μm,分子量约8.3×107,重约10-14克,占核DNA量的5%左右。叶绿体DNA为多拷贝,碱基组成不均一,编码有全部的rRNA、tRNA和部分蛋白质。叶绿体大亚基中有23S,5S,4.5S rRNA,小亚基中有16S rRNA,其中4.5SrRNA只限于显花植物和一部分羊齿类植物。它们在DNA分子上往往以1.6S-23S-4.5S-5S的顺序排列,在16S和23S之间的间隔区段中编码有叶绿体tRNA的基因。在衣藻叶绿体的23S rRNA基因中还发现有长约1kb左右的插入序列(内含子)。除豌豆和蚕豆外所有rRNA的编码基因都有两个拷贝,在DNA分子上成反向重复排列。迄今为止玉米上25个tRNA的编码基因、烟草上29个tRNA的编码基因均已定位。烟草、玉米、菠菜的tRNA的碱基序列分析工作业已进行。叶绿体DNA编码的蛋白质目前已知有二磷酸核酮糖羧化酶的大亚基,H+-ATPase的CF1的α、β、ε亚基和CF0的Ⅰ、Ⅲ亚基(DCCD结合蛋白),32kd的膜蛋白、AP700、AP600,细胞色素f、细胞色素b6、细胞色素b559以及核糖体蛋白CS19、CS4、CS14、CL2等。但是,十分重要的叶绿素合成酶系,电子传递系统中的许多构成因素,光合作用中CO2固定途径中的很多酶类都是由核基因控制的,只靠叶绿体本身的遗传信息量不足以完成本身的组建和行使功能,因此叶绿体只算半自主性细胞器。
起源 主要有共生学说和分化学说。前者认为叶绿体的祖先是蓝藻或光合细菌,在生物的进化过程中被原始的真核细胞吞噬,并共生在一起进化而来。如叶绿体DNA为裸露环状分子,mRNA一般不含多聚A,抗生素抑制蛋白质合成的效果和细菌一样以N-甲酰蛋氨酸为起始氨基酸,叶绿体DNA和核DNA分子杂交率比细菌和蓝藻DNA分子的杂交率低,现存生物中有单细胞蓝藻共生在鞭毛藻内的现象等,都是支持共生学说的有力证据。但共生学说不能解释叶绿体的许多结构和功能受两套遗传系统共同控制以及真核细胞如何进化而来的问题。分化学说认为叶绿体是原始原核细胞的质膜凹陷脱离、并包裹了一些遗传物质逐渐演化而来。两套遗传系统共同控制叶绿体,以及原核细胞和真核细胞间过渡性生物的存在是支持分化学说的证据。
叶绿体chloroplast
植物绿色细胞所特有的能量转换细胞器。一般呈椭圆形,大小约1~3μm×5~7μm×2~3μm,有双层被膜与胞质分开,内有片层膜,含叶绿素,故名。光合作用在片层膜上进行。绝大部分异养生物的有机物质与能量来源都由叶绿体进行光合作用提供。大多数高等植物叶绿体呈透镜状。藻类的叶绿体形态有板状、网目状、螺旋状等。细胞中叶绿体的数目依植物种类和组织类型以及发育阶段而异。单细胞衣藻仅具一个大型叶绿体,大型海藻刺松藻可含数百以至数千个叶绿体。通常一个叶肉细胞含50~200个叶绿体。叶绿体由叶绿体膜、类囊体和基质三部分组成。原核生物如蓝藻无成形的叶绿体,只有简单的片层膜散于细胞质中。
叶绿体
植物绿色细胞中所特有的能量转换细胞器。呈扁球形,长约5~7 μm,短径约2~3 μm,高约1~3 μm。外有被膜,由2层单位膜组成,厚约5~10 nm。内有几个至几十个呈柱形的绿色基粒,每个基粒由10~100个盘形片层构造重叠而成。
叶绿体yeluti
绿色植物细胞中广泛存在的一种含有叶绿素等色素的质体,是植物细胞进行光合作用的场所。高等植物的叶绿体一般为椭圆形或卵圆形,平均直径为4~6微米。一个细胞中可能有10~100个叶绿体,多在核周围或近壁处集中,并能随光线的强弱而移位。电镜下观察,叶绿体为封闭的双层膜结构,内外膜之间平均约为20毫微米的膜间隙。由叶绿体膜所围成的叶绿体腔中、充满液体基质和类囊体膜。每个类囊体是由周围闭合的双层膜组成的扁囊。基粒类囊体的直径约为0.25~0.8μm,厚约0.01μm,由5~30个基粒类囊体叠置成一个基粒,每个叶绿体腔中约有40~80个基粒。基质类囊体横贯基质,延伸出的分枝网管贯穿于两个以上的基粒之间。类囊体与基质接触的外表面有两种颗粒:一是含有RuBP羧化酶的方形颗粒;一是具有ATP酶活性的呈多角形的偶联因子1颗粒。类囊体膜中含有叶绿素和类胡罗卜素等各种光合色素,这些色素与专一蛋白质结合形成复合物颗粒:一是具有光系统Ⅰ活性的小颗粒;一是具有光系统Ⅱ活性的大颗粒。类囊体膜中还有光反应的各种酶系。类囊体腔中充满水溶液。叶绿体腔内基质中,含有少量的环状DNA、RNA和小核糖体,以及大量的RuBP羧化酶颗粒和其他代谢活跃的物质等。叶绿体是光合作用的场所,光反应在类囊体部分进行,类囊体膜中的光合色素分子吸收和传递光能,在反应中心光能转化成化学能,形成ATP和NADPH,同时使水分解放氧;暗反应在叶绿体基质中进行,利用光反应形成的ATP和NADPH,同化CO2形成贮能有机物。叶绿体DNA能以半保留方式复制,并有自己的蛋白质合成系统。叶绿体中的重要蛋白质,分别由核基因和叶绿体基因编码形成。此外,叶绿体基因与细胞质遗传有一定关系。在个体发育中,叶绿体是由前质体分化而来。发育中的幼龄叶绿体分裂能使叶绿体增殖。在光照条件下,白色体可变成具有正常结构和功能的叶绿体。

图509 叶绿体结构示意图
叶绿体
绿色植物细胞中广泛存在的有色质体。形如双凸透镜,直径3—10微米。外包双层膜,内部为由膜层形成的许多扁囊堆积成的基粒,其间充满胶状基质。每一基粒含有百万以上叶绿素分子和酶,能进行光合作用,基质中含有环状DNA分子和70S核糖体。它在遗传上有相对的独立性,能合成自己的蛋白质,但与细胞整体协调统一。
叶绿体
叶绿体为植物细胞内含有叶绿素并能进行光合作用的质体。质体为植物细胞内的细胞器之一。可根据质体含有的色素和功能不同,把它们分为三类,即白色体、有色体和叶绿体。各类质体的基本结构很相似,表面被膜,内含基质,基质内分布有许多基粒。各种色素附着于基粒上,白色体无色,见于胚细胞和生殖细胞中,它们可以积累蛋白质、淀粉和脂肪等物质。有色体含有叶黄素、胡萝卜素和类胡萝卜素等,呈现黄色或橘黄色,存在于花瓣、果实或根中。通常,由于有色体所含有的叶绿素退化,其光合作用能力处于不活跃状态。
叶绿体的形态、大小、数目和分布 可因植物种类而异,但在同一组织的细胞中,则比较稳定。在低等植物,如藻类,叶绿体呈网状、带状或星状,其大小可达100μm。在高等植物,叶绿体则呈球形、卵圆形或盘状,厚约2-3μm,长约4-10μm。叶绿体的数目在各类植物细胞中并不一定。在低等植物的细胞中数目少,如在藻类中,有的仅为一个。高等植物的细胞中约有20—40个叶绿体。有人计算,在蓖麻叶片的栅栏组织细胞中约有36个,在海绵组织中约有20个。叶绿体在细胞体内的数目可自行调节,不足时,可经分裂而增加;过多时,可由退化而减少。叶绿体一般较均匀地分布于细胞质中,但也常常靠近细胞核或集聚于细胞壁。叶绿体可因细胞质流动或光线强弱而改变其在细胞内的位置。
叶绿体的结构 在电镜下,高等植物的叶绿体为膜层结构,表面被以两层单位膜,即外膜与内膜,内部充满基质,基质内分布有40—80个基粒(在低等植物,如绿藻
中,这些膜形成扁平的囊或盘而没有基粒)。基粒的大小随植物种类而不同;从0.3到0.7μm不等。基粒由5到30个圆盘状,直径0.25—0.8μm的封闭小囊组成,这些小囊称为基粒类囊体。从切面上看,基粒类囊体是由两层平行排列的膜所构成,两端密封,每层膜的厚度约7nm左右。两层膜之间叫做基粒类囊体间隙,间隙的大小差别较大,可从4—70nm。这些基粒类囊体似小圆盘一个紧挨一个地互相叠在一起形成柱状,构成了叶绿体内膜系统的基粒片层。叶绿体中除基粒类囊体之外,还有另一种类囊体,名基质类囊体。基质类囊体为位于基粒之间的类囊体,由它们构成叶绿体内膜系统的基质片层。在类囊体的两层膜中,曾发现有大小约20×20×10nm的颗粒嵌入其中,这些颗粒叫做光能转化体。据认为光能转化体是进行光合作用时光能转化的部位。
叶绿体的化学组成 叶绿体主要由蛋白质和脂类组成。此外,还有少量糖类及核酸。从高等植物的叶绿体化学成分来看,其中蛋白质约占干重的35%—55%,脂类约占干重的20%—30%。大约80%的蛋白质是不溶性的,它们与脂类结合,形成脂蛋白。组成叶绿体蛋白质的氨基酸成分,虽可因季节及制备法的不同而发生差异,但从菠菜叶绿体的化学分析,仍可观察到下列特点:疏水性氨基酸含量丰富,半胱氨酸含量贫乏;酸性氨基酸及其酰胺含量较多,碱性氨基酸含量较少等。脂类主要包括中性脂、固醇、磷脂和蜡等。叶绿体中含有叶绿素和类胡萝卜素。高等植物细胞的叶绿体具有两种,即叶绿素a和叶绿素b。它们均匀地分布于基粒片层和基质片层之中。低等植物尚发现有叶绿素C和叶绿素d等。叶绿素的吸收光谱与光合作用效率相一致,这说明叶绿素是主要的捕捉能量的分子。类胡萝卜素包括β胡萝卜素和叶黄素等。通常被叶绿素所掩盖,当叶绿素含量减少时,它们才显示出来。叶绿体中RNA的平均含量为干重的5%左右。在叶绿体的基质中发现有DNA,它的存在可由电镜照片予以确定,但含量很少,只占 .5%左右。 除此之外,在叶绿体的类囊膜上还存在电子载体蛋白质,如细胞色素、质体蓝素和铁氧化还原蛋白等。在进行光合作用时,这些电子载体蛋白质具有传递电子的作用(见图)。
.5%左右。 除此之外,在叶绿体的类囊膜上还存在电子载体蛋白质,如细胞色素、质体蓝素和铁氧化还原蛋白等。在进行光合作用时,这些电子载体蛋白质具有传递电子的作用(见图)。
叶绿体的功能 叶绿体的主要功能是进行光合作用。光合作用是叶绿体内的叶绿素利用日光将二氧化碳和水合成碳水化合物的过程。
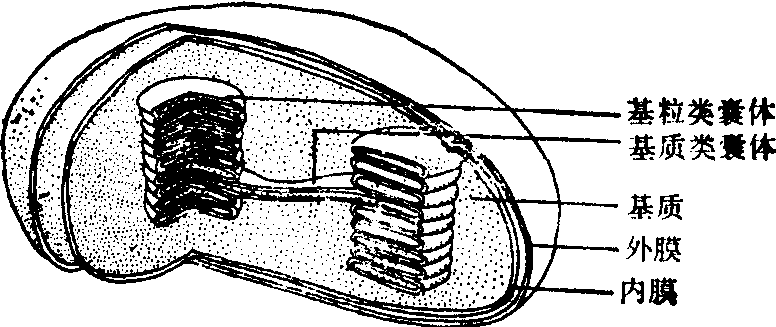
叶绿体结构图解

+化学能2 819kJ(674 000cal)
上述化学反应过程说明叶绿素能将捕捉的日光能转变为化学能,並贮存于生成物质的化学键中。光合作用涉及两种连续反应的过程,即光反应和暗反应。光反应需要在日光下进行,其主要作用是叶绿素吸收日光的能量将水分解为H和OH。其中OH自行合成H2O2之后再分解而生成O2。所以,光合作用生成的O2不是来自CO2,而是来自水。由水分解出来的H再分裂为H+和电子,电子经过电子传递系统,可通过铁氯蛋白,把NADP(辅酶Ⅱ)还原为NADPH(还原型辅酶Ⅱ),并在此过程中通过光合磷酸化作用生成ATP。暗反应并不需要日光的参与,此过程是利用NADPH和ATP,经过一系列复杂反应,把CO2还原为碳水化合物。
叶绿体
chloroplast;chlorophyll body
叶绿体
chloroplast
- 器度是什么意思
- 器度狭窄急躁是什么意思
- 器座是什么意思
- 器异是什么意思
- 器弩是什么意思
- 器弩悉弄赞普是什么意思
- 器形是什么意思
- 器彩是什么意思
- 器待是什么意思
- 器必试而后知其利钝,马必驾而后知其驽良是什么意思
- 器怀是什么意思
- 器性是什么意思
- 器技是什么意思
- 器抱是什么意思
- 器敬是什么意思
- 器木是什么意思
- 器材是什么意思
- 器材分类编码是什么意思
- 器材掩蔽所是什么意思
- 器材消耗是什么意思
- 器杖是什么意思
- 器械是什么意思
- 器械与其备二之不精,不如精其一之为约,二而精之,万全之虑也。是什么意思
- 器械体操是什么意思
- 器械体操规则是什么意思
- 器械体操训练是什么意思
- 器械对练是什么意思
- 器械形变弹力是什么意思
- 器械感是什么意思
- 器械手推车是什么意思
- 器械掉地是什么意思
- 器械断裂是什么意思
- 器械用具是什么意思
- 器械结构是什么意思
- 器械训练是什么意思
- 器械轴是什么意思
- 器械辅助按摩法是什么意思
- 器械运动图解是什么意思
- 器械锻炼是什么意思
- 器泳是什么意思
- 器满则倾是什么意思
- 器满则溢,人满则丧是什么意思
- 器满则溢,人满则丧。是什么意思
- 器满则覆是什么意思
- 器满将覆是什么意思
- 器满意得是什么意思
- 器满招覆是什么意思
- 器爱是什么意思
- 器物是什么意思
- 器物上便于提系的部分是什么意思
- 器物上便于握持的部分是什么意思
- 器物上像棍的部分是什么意思
- 器物上像疤一样的痕迹是什么意思
- 器物上可开关的机件是什么意思
- 器物上将要裂开的痕迹是什么意思
- 器物上用于操作、转动的部分是什么意思
- 器物上用来结系或攀手的带是什么意思
- 器物上的花纹装饰是什么意思
- 器物上的装饰品是什么意思
- 器物上象颈的部分是什么意思