X射线衍射Xray diffraction
X射线照射晶体时发生的衍射现象。X射线是波长很短的电磁波,当照射晶体时,晶体中各原子的束缚电子受迫振动,散射出频率与入射X射线相同的电磁波,这些散射波相互干扰而形成一定的衍射图样,由特殊的照相机或计数管衍射仪记录,其特征与晶体中原子的空间排列有关,据此测定晶体的微观结构。20世纪50年代由X射线衍射法对DNA和肌红蛋白的晶体结构分析取得突破性成功,成为分子生物学发展的重要里程碑。由于其在测定大分子能给出精确、完整的结构,至今仍是其他方法无法取代的重要技术而广泛用于物理学、化学、矿物学和生物科学;在土壤科学中,X射线衍射可有效地鉴定土壤胶体矿物。
X射线衍射
X射线衍射分析是研究物质细微结构的一种技术。目前关于无机分子、一般有机分子和生物大分子的准确立体结构知识,几乎全部来自这一技术。
任何物质的性质,都与其分子的结构有密切的关系。因此,在分子和亚分子水平了解物质的细微结构,对于认识自然和改造自然都有重要意义。迄今,对于物质细微结构的认识,有两条基本途径。其一是直接显微放大,即利用各种光学放大系统,通过直接观察获得物质细微结构知识,其基本优点是直观。但它的应用受到极大限制,因为根据光学成像理论,任何显微放大系统的分辨率(系统所能分辨的最小距离) 都不能超过其所用光波波长的一半。一般光学显微镜,即使用紫外光作光源,也有大约4000Å(1Å=10-8cm) 左右的波长, 它的极限分辨率不可能超过2000Å,加上技术实现过程中各种因素的限制,实际所能达到的分辨率比这低得多。生物大分子 (线性聚合的纤维分子除外),一般都在几百埃的数量级,而组成分子的原子间的距离都在埃的数量级,所以,用普通光学显微放大系统是无法研究物质的分子结构。电子射线的波长远<1Å,用它作光源制成的显微放大系统原则上应当能分辨原子。目前,由于技术上的一些难题(如生物样品的反差不足) 尚未解决,电镜还只能揭示分子的大小、形状、对称性和聚集态等方面的细微结构信息。
研究物质细微结构的另一基本途径是间接认识。它观察和记录一些与物质的微观结构即分子结构直接相关的物理一化学现象,利用观测到的现象与物质结构的特征关系,揭示物质的分子结构。属于这个范畴的方法和技术很多,就研究分子的立体结构而言,X射线衍射分析是目前最有效的一项技术。它利用X射线穿过晶态物质所产生的衍射效应,可以将一个分子的所有非氢原子和部分氢原子的空间位置准确地揭示出来,在原子水平全面地提供关于一个分子的立体结构信息。
X射线衍射基本原理 衍射是一种物理现象。当一种电磁辐射穿透物质时,周期递变的电场会迫使组成物质的荷电粒子(电子、原子核)振动,振动着的每个荷电粒子都是一个新的辐射源,向四周发出球形次生级射线,称为散射(图1a)。
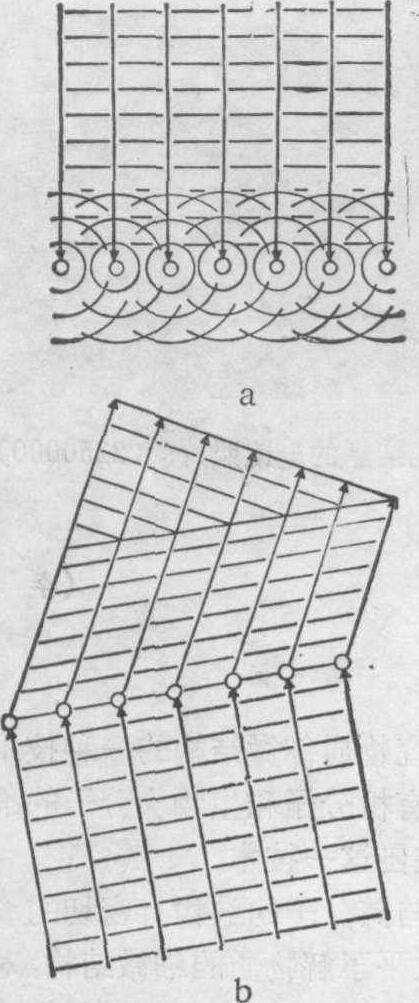
图1 衍射原理示意图
a散射 b干涉
这种散射光的主要部分,与入射光有相同的波长。在入射波长与散射元的大小差不多、而且散射元是周期地规则排列的条件下,散射光按相位关系彼此迭加 (即所谓干涉),在一些方向加强,在另一些方向减弱 (图1b),结果在入射线以外的方向,可观测到许多明暗不同的辐射,这一现象称为衍射。用于结构分析的X射线的波长约1.5A,组成分子的原子间距离大约也在这个数量级,而且在晶态物质(即晶体)中,这些原子(在三维空间中)是严格地按一定周期排列的。当X射线穿过晶体时,就产生X射线衍射效应 (图2)。如果是完整的单晶体,则在三维水平上可得到完全的X射线衍射效应; 这时,晶体就是一个三维的原子光栅。从这样的衍射分析,可以得到分子的三维结构信息。如果是准晶态即在一个方向排列规则有周期性,而在其他两个方向取向随机,如纤维,所得到的X射线衍射效应是不完全的,提供的结构信息也是局部和粗糙的,需要大量其他资料辅助方能解决一个完整的结构问题。这就形成了X射线衍射分析的两个不同领域,前者称为单晶结构分析,后者主要是纤维和多晶衍射分析。单晶结构分析威力最大,但要求的条件也苛刻,许多生物物质(如DNA、病毒、膜蛋白)要获得单晶十分困难,只能进行纤维衍射分析。在实际应用中,X射线衍射分析的这两个方面都有着重要作用。
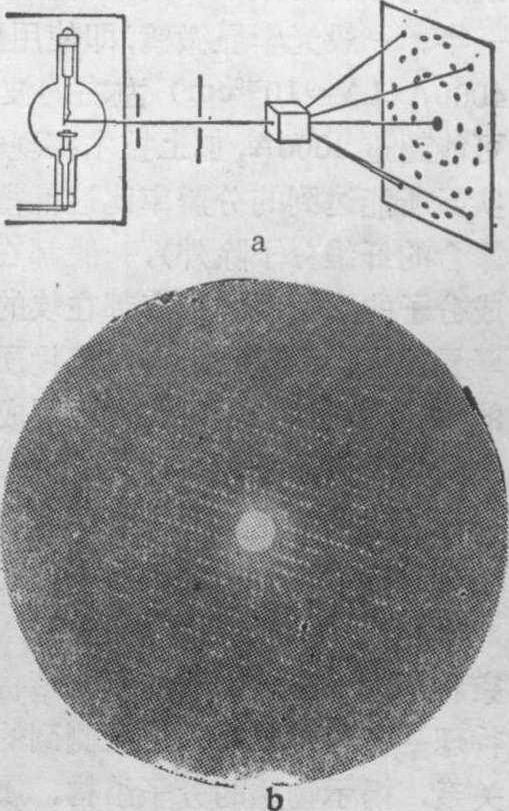
图2 X线衍射示意图
a摄取衍射照片示意图 b肌红蛋白晶体衍射照片
X射线穿过晶体时,产生散射的基本公式是Thomson公式,它表明散射强度与散射元质量的平方米成反比
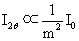
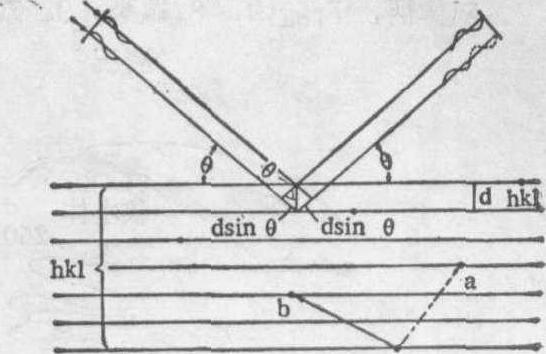
图3 衍射方向的决定(Bragg公式)
考虑X射线与电子的相互作用,其最后所得结果——X射线像,就是一个晶胞 (即晶体的一个基本单位) 中的电子密度分布图。定量描述X射线衍射效应的基本定律是Bragg(布拉格)关系式:如有一波长为λ的光射到间距为d的点阵平面上(图3 ),掠射角为θ,则当满足条件
2dsinθ=nλ
时发生衍射,衍射光线的方向即为所取平面的反射光线的方向。式中n为整数,表示形成衍射的级别;θ是入射光与所取平面间的夹角,等于衍射线与入射线间夹角的一半;发生反射的这组平面,用一组三联指标(h,k,1)来表征,显然,它也就是相应衍射光的指标。从图3很容易导出这一关系。
X射线衍射光的方向,取决于晶体中晶胞的形状、大小和排列方式;而各衍射线的强度,则决定于晶胞中原子的空间位置。所以,测定、记录了某一物质所产生的X射线衍射在空间的分布和所有衍射线的强度,即可分析出该物质所有非氢原子在晶胞中的空间位置和排布。衍射效应与晶胞中原子位置的定量关系,可用“结构因子”的量Fhk1来描述。设晶胞的独立单位中含有j个原子,各原子的散射因子是fj,各原子的坐标是Xj,Yj,Zj,则其(hk1)平面所产生的衍射波函数Fhkl为
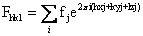
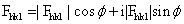
前面已经提到,X射线散射主要是由原子的电子产生的,X射线衍射分析的最后结果就是求得晶胞中的电子密度分布。由于每一个原子相当于一定数量的凝集体,所以这种电子密度分布就反映了原子在相应空间的分布(图4)。在肌红蛋白血红素三维电子密度分布图中清楚地显示出各组成原子。晶体具有三维周期的规则排列,所以晶体中的电子密度分布函数也将是一个三维周期函数,因而可用Fourier(傅里叶)级数来表征:
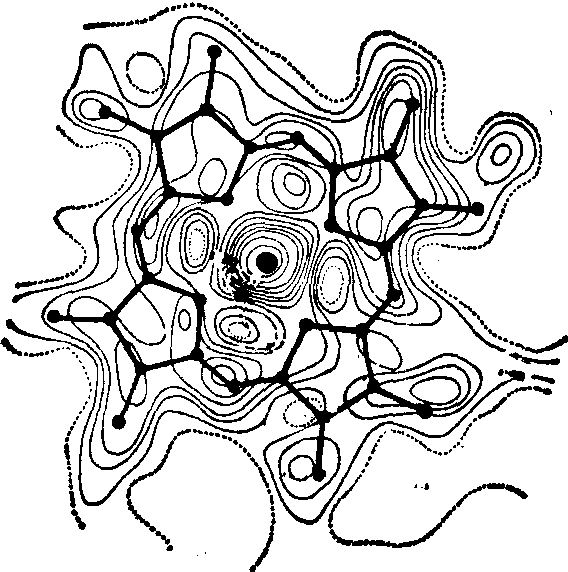
图4 肌红蛋白中血红素的三维电子密度分布图
式中ρ是在坐标为(x,y,z)的空间某一点上的电子密度,(h,k,1)是衍射线的指标,vc是晶胞的体积,Fhkl就是结构因子。显然,除Fhkl外,这些都是已知或易测量;一旦测得Fhkl,ρ(x,y,z)的计算也极易进行。Fhkl的振幅部分丨Fhkl丨很易由测量衍射光的强度I直接测出:
丨F丨2=KI
式中,K是由实验条件和晶体本身的几何、物理状态决定的一个校正量。但是Fhkl量的另一部分——相位φ,至今没有任何方法可以直接记录。这就是说,在衍射分析的实验观测中,丢失了一半信息。如何将失去的相的信息寻找到,完成电子密度函数的傅里叶加和,这就是X射线衍射分析的中心问题,即所谓“相位问题”。解决相位问题的不同途径,就形成X射线衍射分析中的不同方法,现在常用方法有尝试法、向量空间法、同晶置换法、反常散射法、直接法等。对蛋白质和核酸等生物大分子,最有效的是多对同晶型置换法。利用这一方法测定生物大分子的单晶结构,一般包括如下步骤:
(1) 培养结晶: 为进行满意的X射线衍射分析,一般要有线度在1mm左右的单晶体。这对生物材料,有一定困难,必须很好掌握影响结晶的因素,如溶解度、pH、温度、离子强度等。重要生物材料常常得之不易,故近年来特别重视发展微量生长技术。
(2) 制备重原子衍生物:在不破坏原有晶体的前提下,将重原子引入蛋白质晶体,利用重原子的衍射贡献来解决相位问题。一般至少需要两个衍生物,衍生物数量越多,质量越好,所获分析结果越佳。
(3) 收集和校正衍射数据: 收集方法主要有感光和电离两种,常用设备有旋进相机和电子计算机检测的光密度测量计以及四圆衍射仪。
(4) 测定衍射线相对相位: 通过计算分析进入晶体中的重原子的位置,求得所有衍射线的相对相位。
(5) 计算和解释三维电子密度图:由于运算量极大,电子密度函数必须在电子计算机上进行计算。结果所能提供的结构信息, 决定于分辨率。 在低分辨率 (4Å以下)一般只能得到分子的大小、形状及部分肽链和核糖-磷酸骨架的走向; 在中等分辨率时(4~2Å), 可以确定分子骨架的走向和大部份侧链的取向; 在高分辨率 (高于2A)时,可以确定绝大多数非氢原子的空间位置。
由于X射线的波长极短,所以用X射线衍射分析可以达到很高的分辨率。在单晶结构分析中常用的X射线波长是1.54Å(CuKα线), 所以它能提供高于1Å的分辨率,可以清楚地分辨原子。一般有机分子的结构分析,都已达到这一水平。对生物大分子,由于其晶体的固有弱点,至今高于2A分辨率的结果并不很多,只有极少数达到1.5A或稍高水平。由于X射线衍射分析要求对象必须是晶体,故所获精细三维结构信息都是静态的,更严格地说,是在时间上和空间上平均的结构。应用现有的技术,可以测定构象变化前、后的结构,也可以测定在化学上“凝固”起来的一些中间态的结构,但不可能用X射线衍射分析直接观测过渡性的中间态,或测定构象变化的速率。这是这一技术的局限性。
在人类认识物质结构的历史中,X射线衍射技术作出了巨大的贡献,并早已发展成一门独立的学科——X射线晶体学。它为人们提供了大量的物质结构信息,成为认识物质微观世界的一种有力武器。
X射线衍射在生物医学领域中的应用 在生物学和医学领域,近二、三十年来,X射线衍射分析成功地应用于生物大分子主要是核酸和蛋白质的三维结构分析,取得了十分突出的成就。五十年代初期,英国Wilkins获得了DNA纤维的极好的X射线衍射照片(图5)。正是以这一DNA的衍射图象为基础,英国物理学家Crick与美国生物学家Watson合作研究,在1953年阐明DNA的分子结构是一右手双螺旋,这就是著名的Watson-CrickDNA双螺旋结构模型(图6)。
这一结构模型提出后,遗传密码很快全部“破译”,遗传信息的传递得到完全了解,人类对生命现象的认识很快被推进到一个全新的深度——分子水平。在这一基础上,分子生物学应运而生,遗传工程迅速发展,推动了生物学的大革命。所以有人认为,Watson-Crick右手DNA双螺旋结构模型是分子生物学的一块基石,在生物学中可与达尔文的进化论、孟德尔的遗传定律相媲美。1973年后,美国麻省理工学院的Rich小组等又用X射线单晶衍射分析,测定了第一个tRNA——酵母苯丙氨酸转移核糖核酸的三维结构,分辨率达到2.5Å,图7是它的原子结构模型。1979年
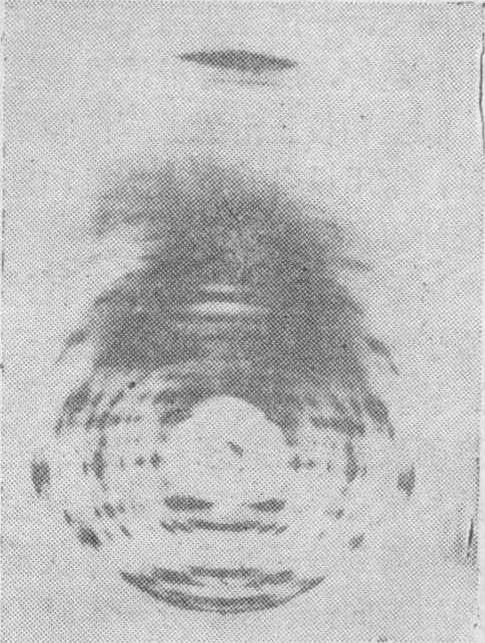
图5 B型DNA纤维的X射线衍射图
12月,Rich小组又报道完成了一含六个碱基对的 DNA片段的X射线单晶衍射分析,其核苷酸顺序是 d(CpGpCpGpCpGp)。结果分辨率达到0.9A,因此看到了除氢以外的所有原子。这是人类首次在原子水平观察到一个DNA片段。令人惊奇的是,他们发现,观察到的结构是一左手双螺旋,它与Watson-Crick右手双螺旋模型具有明显不同的结构特征,主要是它的脱氧核糖——磷酸骨架具有反常的Z字形走向,并以左手方式旋转,他们将它命名为Z 型DNA。图8示Z-DNA与B-DNA(Watson-Crick模型)两种结构模型,图中用粗实线将磷酸基团的位置连接起来,显示出分子骨架的走向。从图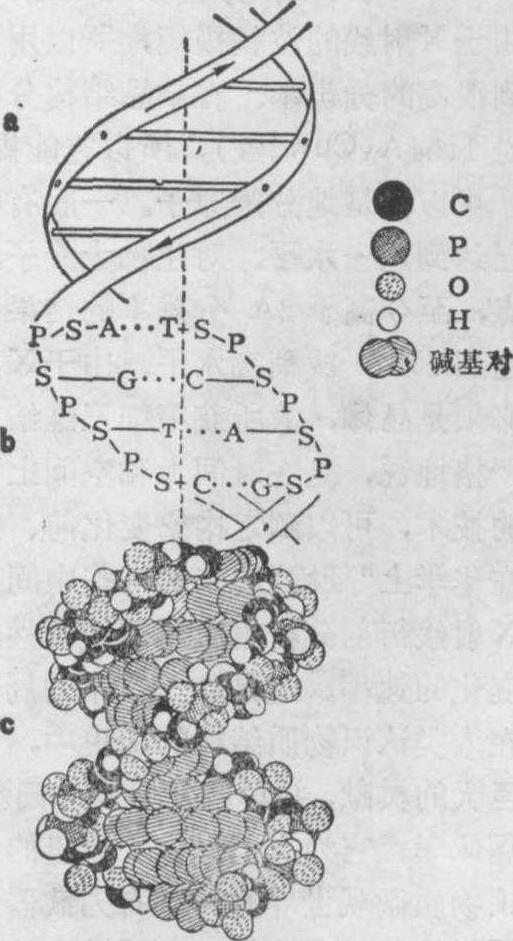
图6 Watson-Crick DNA双螺旋结构模型(B型DNA)
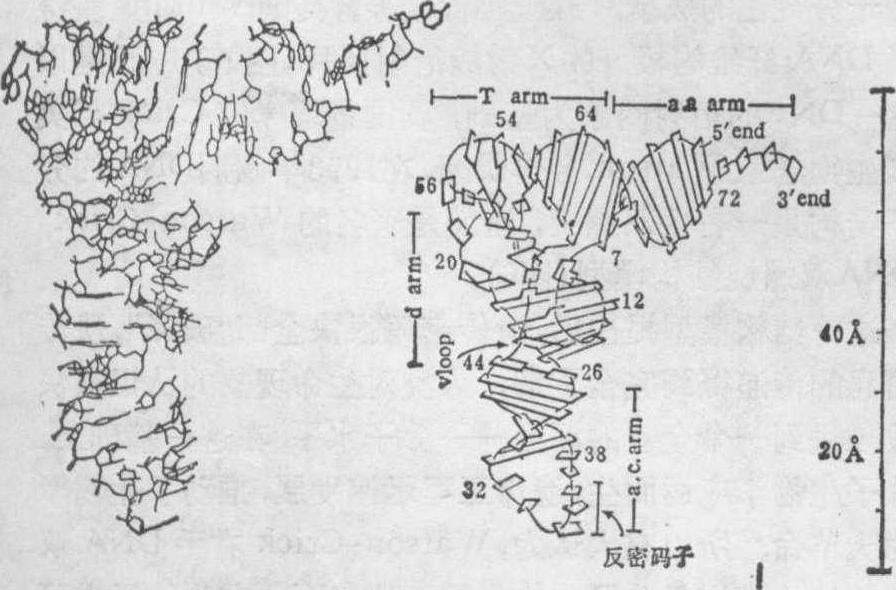
图7 酵母苯丙氨酸tRNA的三维结构
左图为原子结构模型,右图为核苷酸骨架和碱基示意图
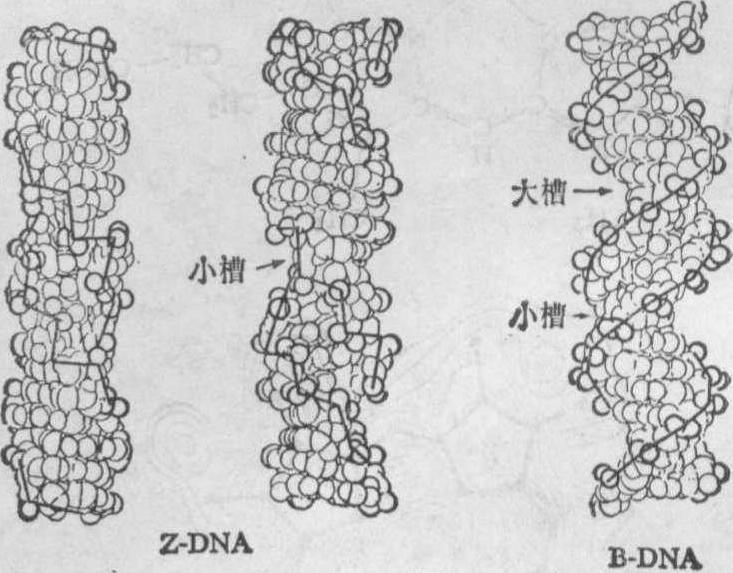
图8 Z-DNA(左)与B-DNA(右)的比较
X射线衍射技术在蛋白质三维结构研究中的应用,取得了更加突出的成就。在X射线衍射现象发现不久,就被用于研究蛋白质的结构。这要归功于这一技术的创始人Bragg及他建立的结晶学派,包括Astbury、Bernal等著名学者,他们不仅致力于发展晶体结构分析技术,而且对生命物质表现了强烈的关心。1930年前后,Astbury研究了一系列蛋白质和核酸纤维的衍射花样,他发现头发伸长时产生的衍射图样与不伸长时是不一样的,后者称为α型,前者称为β型。这使他认识到,宏观形态的变化必然伴随着分子结构的变化。这在今天是一个极为普通的概念,但在当时却是一个全新的思想,它使人们开始意识到,生物大分子的结构及其变化对了解生物学过程具有重要作用。这一思想发展,导致开辟了一个新的研究领域——分子生物学。所以,尽管Astubry提出的所有蛋白质模型,后来都被证明是错误的,但它的这种开创性作用始终为人们所赞誉。直到1951年,Pauling和Corey在用X射线衍射分析测定大量氨基酸和小肽的精细分子结构基础上,首次提出正确的蛋白质结构模型,这就是有名的α螺旋和β折叠结构(图9)。
Bernal领导的另一个小组,包括Hodgkin、Perutz等人,从三十年代中期就开始用X射线衍射分析去解析胃蛋白酶、血红蛋白这一类蛋白质单晶体的结构。经过长时间的摸索,1953年Perutz首次提出应用多对同晶置换法来解决蛋白质晶体衍射分析中的相位问题。经过几年技术上的完善,在1960年,同一实验室的Kendrew首次应用这一方法测定了肌红蛋白的三维结构(图10),由此开辟了蛋白质结构的新天地。由于方法和技术逐渐成熟,最近几年每年有将近10~15个蛋白质结构测定出来。目前,已在3.5Å以上分辨率测定了50多种100多个蛋白质的三维结构。 同晶置换途径达到的最高分辨为1.8Å,结合外推相位通过修正达到的分辨率已高于1.5A,甚至可见到
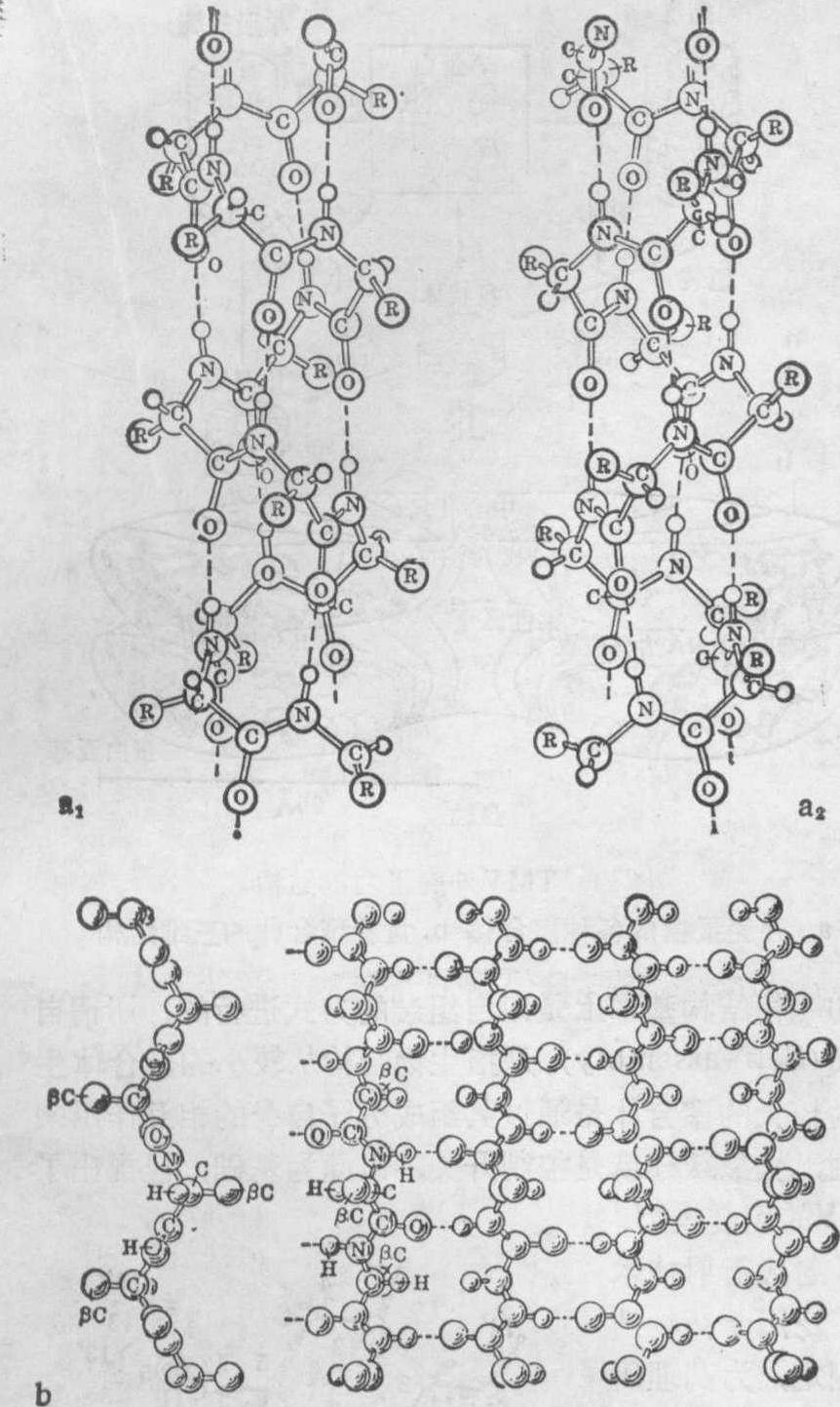
图9蛋白质结构模型
a. α螺旋(a1左手螺旋,a2右手螺旋) b·β折叠结构
❶酶;
❷氧化还原蛋白;
❸运载和贮存蛋白;
❹激素;
❺抗体蛋白。其中以酶的数量最多,约占60%。此外,尚有少数光合作用蛋白、植物凝集素、肌肉蛋白、蛇毒蛋白的结构也已测定。我国科学工作者在1969~1973年期间先后在4A、2.5A和1.8A分辨率,测定了激素蛋白胰岛素的晶体结构,达到世界先进水平(图11)。
X射线衍射技术的成功应用,使我们有可能在比过去深透得多的尺度上了解蛋白质结构与功能的关系,并直接揭示蛋白质一些重要作用的具体过程。近年来这方面取得的成效是十分显著的,可举下列四个方面作为代表:
(1) “分子肺”的发现: 血红蛋白呼吸功能的阐明。由于血红蛋白分子在机体生理——生化过程中的关键性作用,所以了解它的作用机理具有十分重要的意义。Perutz小组先后在1960、1968、1970年用X射线衍射分析测定了氧合血红蛋白(HbO2)和脱氧血红蛋白 (HbA)的晶体结构,分辨达到2.8A。通过这两种结构的比较发现,从结

图10 肌红蛋白的三维结构

图11 胰岛素二聚体的三维结构模型
合氧到失去氧,血红蛋白分子的四级结构有显著变化,结果亚基间距离在HbA中要比在HbO2中增大6.5Å(图12)。这种随着氧的得失,血红蛋白的一张一合的情况正与我们肺的呼吸相似。由此发现,血红蛋白并非是一个氧气瓶,而是一个“分子肺”。进一步的研究探明了这一微妙的分子机构的作用机理,并首先了解一个蛋白质变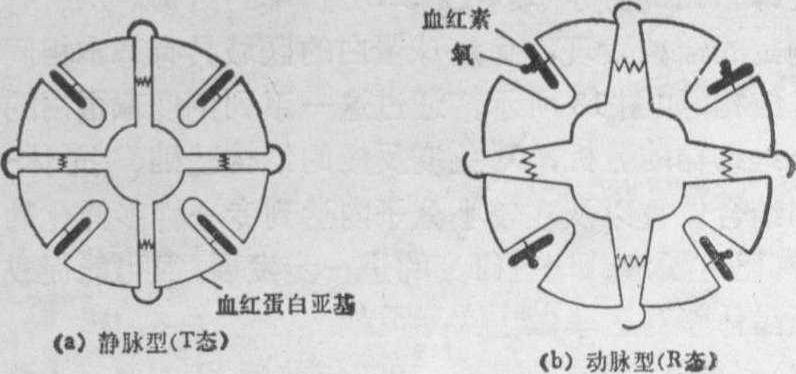
图12 血红蛋白构象示意图
(a)脱氧血红蛋白 (b)氧合血红蛋白构象示意
(2) 一些酶的作用机理——酶与底物的过渡态互补:自1965年测定第一个酶(溶菌酶)的晶体结构以来,在过去十几年里已高分辨率地测定了四十多个酶以及一些酶与其竞争性抑制剂或类底物复合物的结构,所有这些结果都直接显示出酶与底物的作用是诱导配合式的动态过程。在这一过程里,酶分子以及底物都要发生相当明显的构象变化,其作用是导致空间和电的张力状态,产生最佳的熵因素(如反应物的接近),从而使整个体系趋向反应的过渡态,促成酶与底物的过渡态互补。
(3)免疫球蛋白的三维结构——抗体—抗原作用的结构基础: 1973年以来,一系列免疫球蛋白的片段已用X射线衍射分析在高分辨率测定,包括IgG New的Fab片段、小白鼠IgA(K) McPC603的Fab片段、Bence-Jones
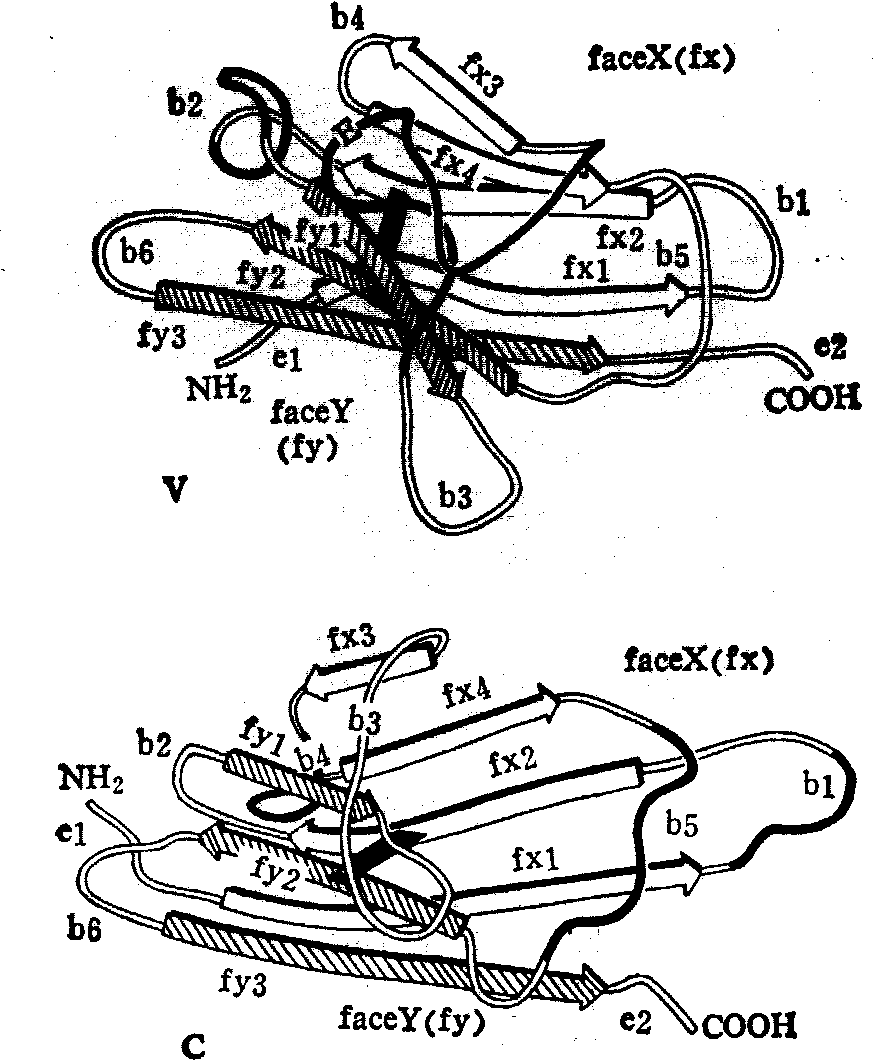
图13 免疫球蛋白的可变区(V)和恒定区(C)的三维结构
本周蛋白Meg、REI等。此外1976年已得到一个完整的免疫球蛋白的分子(IgG1) 5Å分辨率的结构。 从所有这些测定的结构发现,免疫球蛋白的区域具有基本相同的三维结构,如图13所示。通过这一系列免疫球蛋白的片段三维结构的分析,对免疫反应的结构基础、抗体-抗原的结合位置以及在较小分子内达到专一性多样化的方式,都已有了解。这些研究的进一步发展,很可能促成蛋白质晶体学对医学做出实际贡献。(4) 分子体系的自组装: 1976年前后,Butler等人测定了烟草花叶病毒(TMV)外壳蛋白的三维结构(图14),结合噬菌体以及细菌鞭毛体外组装的研究,他们提出超分
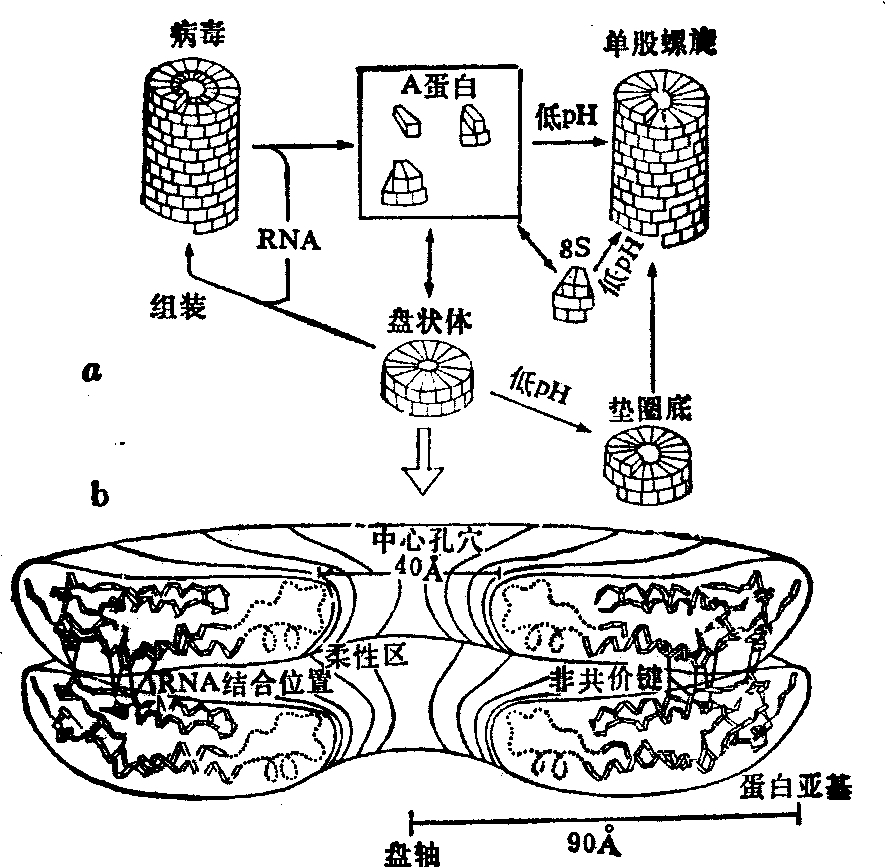
图14 TMV外壳蛋白的结构
a.外壳蛋白的各种聚合体b.盘状聚合体的三维结构
X射线衍射技术也广泛用于药物结构测定,为药理研究、药物设计和合成提供结构基础。在解决一些化学分析的歧误和疑难方面,这一技术具有特殊的作用。我国科学工作者已测定了不少中、西药的晶体结构,图15是由我国首次测定的抗疟药物青蒿素的结构。
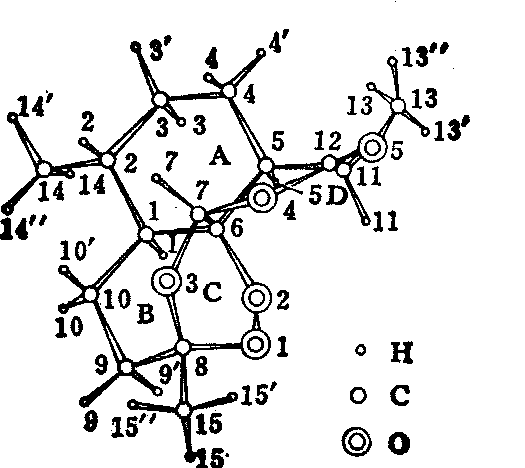
图15 青蒿素的晶体结构
- 李家庄的变迁是什么意思
- 李家恒是什么意思
- 李家成是什么意思
- 李家房子渡口是什么意思
- 李家拳是什么意思
- 李家振是什么意思
- 李家教是什么意思
- 李家文是什么意思
- 李家明是什么意思
- 李家智是什么意思
- 李家权是什么意思
- 李家村是什么意思
- 李家村民兵连是什么意思
- 李家村遗址是什么意思
- 李家杰是什么意思
- 李家沟反维持记是什么意思
- 李家沟村是什么意思
- 李家沟石疙瘩遗址是什么意思
- 李家沟贝墓是什么意思
- 李家沱工人俱乐部是什么意思
- 李家河村是什么意思
- 李家河白氏夫妇墓是什么意思
- 李家河石窟是什么意思
- 李家治是什么意思
- 李家泉是什么意思
- 李家洼村是什么意思
- 李家海是什么意思
- 李家渠镇是什么意思
- 李家湾遗址是什么意思
- 李家煌是什么意思
- 李家珍是什么意思
- 李家琳是什么意思
- 李家瑞是什么意思
- 李家益是什么意思
- 李家短枪是什么意思
- 李家章是什么意思
- 李家耀是什么意思
- 李家老屋新莽墓是什么意思
- 李家联是什么意思
- 李家胡同村是什么意思
- 李家能是什么意思
- 李家英是什么意思
- 李家营子金银器是什么意思
- 李家训是什么意思
- 李家辑是什么意思
- 李家道是什么意思
- 李家鈺是什么意思
- 李家鏊是什么意思
- 李家钰是什么意思
- 李家铨是什么意思
- 李家馨是什么意思
- 李家駒是什么意思
- 李家驹是什么意思
- 李家骏是什么意思
- 李家骥是什么意思
- 李家骥(1)是什么意思
- 李家骥(2)是什么意思
- 李家骧是什么意思
- 李家鳌是什么意思
- 李家鼎是什么意思