钾生理功能physiological function ofpotassium
钾对植物结构、组成、代谢、生长发育及抗逆性的效应。钾能促进碳水化合物及蛋白质代谢。缺钾时,植物生长柔弱。抗逆性减低。在植物渗透调节中,K+有关键性的作用,植物细胞伸长、气孔运动以及由膨压变化引起的运动等都是与K+在渗透调节和水分平衡中的功能相联系的。钾促进蛋白质合成、促进光合作用及光合产物运往收获器官,故可增加产量。
酶的活化剂 已发现60种以上的酶可被K+活化。其中许多酶已在高等植物中发现,虽然其它一些一价阳离子对某些酶也有一定程度的活化作用,但K+在活化酶的作用方面是最有效的阳离子。例如K+是ADP葡萄糖—淀粉合成酶的活化剂,虽然Na+也有活化作用,但K+的活化作用要比Na+高3~4倍。很多催化磷酸基转移的酶都为K+所活化(表1)。K+诱导酶的构型变化,加快催化反应的速度,在有的情况也可增加酶与底物的亲和力,使反应易于进行。K+活化膜结合的ATP酶的作用,促进了K+本身的吸收和运输(见钾的吸收和运输),并使K+发挥其渗透调节及促进细胞伸长的功能。
表1 K+活化的一些酶
| 酶的名称 | 酶所催化的反应 |
| 丙酮酸激酶 | PEP+ADP→丙酮酸+ATP |
| 乙酸硫激酶 | 乙酸+ATP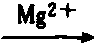 乙酰-AMP+PPi 乙酰-AMP+PPi乙酰-AMP+CoA 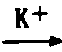 乙酰CoA+AMP 乙酰CoA+AMP |
| 琥珀酰CoA合成酶 | 琥珀酸+CoA+ATP→ 琥珀酸CoA+ADP+Pi |
| 谷氨酰半胱氨酸合成酶 | 谷氨酸+半胱氨酸+ATP→ 谷氨酰半胱氨酸+ADP+Pi |
| 甲酰四氢叶酸合成酶 | 甲酸+四氢叶酸+ATP→ 甲酰四氢叶酸+ADP+Pi |
| ADP葡萄糖-淀粉合成酶 | ADP葡萄糖+(1,4-α-D-葡糖基)n→ (1,4-α-D-葡糖基)n+1+ADP |
| 天门冬酰胺酶 | 天门冬酰胺→天冬氨酸+NH3 |
| ADP葡萄糖焦磷酸化酶 | ATP+α-D-葡萄糖-1-磷酸→ ADP葡萄糖+PPi |
| UDP葡萄糖焦磷酸化酶 | UTP+α-D-葡萄糖-1-磷酸→ UDP葡萄糖+PPi |
| 磷酸化酶 | (α-1,4-葡糖基)n+Pi→ (α-1,4-葡糖基)n-1 +α-D-葡萄糖-1-磷酸 |
| RuBP羧化酶 | RuBP+CO2→3-磷酸甘油酸 |
| 焦磷酸磷酸水解酶 | H2O+PPi→2Pi |
| ATP磷酸水解酶(Mg2+) | ATP+H2O→ADP+Pi |
| ATP磷酸水解酶(Ca2+) | ATP+H2O→ADP+Pi |
注:PEP为磷酸烯醇丙酮酸;ATP为腺苷三磷酸;ADP为腺苷二磷酸;AMP为腺苷一磷酸;Pi为无机磷酸;PPi为焦磷酸;CoA为辅酶A;UTP为尿苷三磷酸;UDP为尿苷二磷酸;RuBP为1,5-双磷酸核酮糖
从表1可见,K+通过活化酶的作用,多方面参与植物代谢过程。
光合作用 叶绿体是光合作用的重要细胞器,适量施钾的植物叶绿体内有较多基粒;缺钾时叶绿体的基粒较少,使光合作用受到影响;图1为盆栽杂交稻威优35号,在分蘖期不施钾及适量施钾处理的叶绿体超微结构的电镜照片(放大12000倍)。从图中可见钾缺乏使叶绿体失去正常结构,其形状较短而圆,基粒明显较少;适当施钾处理的叶绿体呈扁圆形,基粒较多。观察番茄不同钾水平处理叶的叶绿体情况亦相似。
缺钾还使叶绿体发育迟缓而解体较早,图2示水稻缺钾处理(盆栽,不施钾)及适量施钾处理的叶绿体发育情况,从图2中可见,出叶后第2天缺钾叶的质体中尚有原片层体,而适量施钾的质体内则原片层体已发育成片层体。出叶25天后,缺钾叶绿体已开始解体,片层结构趋于模糊,当时叶片的净光合率明显下降,而施钾处理的叶绿体仍具有较完整的超微结构,叶片的净光合率明显高于不施钾处理。不同钾水平处理植株叶片发育过程中,其叶绿体的发育情况与叶片净光合率的动态变化进程相吻合,如图3所示。缺钾使叶片发育过程中净光合率降低而暗呼吸速率增高,故光合产物形成量减少而消耗量增多,易引起早衰。
施钾对植物光合作用的各个环节都有促进作用,包括希尔反应(Hill reaction)。光合电子传递、光合磷酸化作用、CO2的固定和同化以及光合产物的运输各方面。缺钾时,这些环节都受到抑制,所以净光合率明显降低;光合作用是产量形成的基础,因而影响产量。
渗透调节 K+是细胞液中主要的阳离子,约占总渗透浓度的40%,甚至更高,说明K+在渗透调节中的重要性。细胞伸长与其含钾量有密切的联系,K+同有机酸阴离子是细胞伸长中液泡内积累的主要溶质,例如,棉花纤维生长时,在细胞内积累K+及苹果酸阴离子,二者作为主要的溶质,使细胞膨压增高,促进纤维伸长生长。当棉纤维生长最快时,也是纤维内K+与苹果酸阴离子积累量达高峰时,到棉纤维生长速度下降时,K+及苹果酸阴离子含量以及膨压也都降低。棉胚珠在黑暗中固定HCO-3,主要产生苹果酸;当缺钾或大气中CO2不足时,细胞膨压降低,棉纤维生长受到影响,使棉纤维变短。
由于缺钾时细胞膨压减低,故生长受阻而使细胞较小,植物叶面积也因此减小。
细胞伸长涉及大液泡的形成,这需要增加细胞壁的可塑性同延伸性,吲哚乙酸(IAA)具备这种功能,它促进细胞伸长。但在缺钾时,IAA诱导的伸长即降低甚至停止,因为IAA诱导的细胞伸长是与H+排出相联系的。用胚芽鞘试验的结果表明,H+排出与K+内流保持电化学平衡的关系,缺K+时就影响H+排出,因而导致IAA诱导的伸长受到抑制。赤霉酸(GA)诱导的茎伸长亦需要K+,缺钾时也受抑制。伸长生长对缺钾最敏感,往往在光合作用受抑制之前,伸长生长即受到抑制。
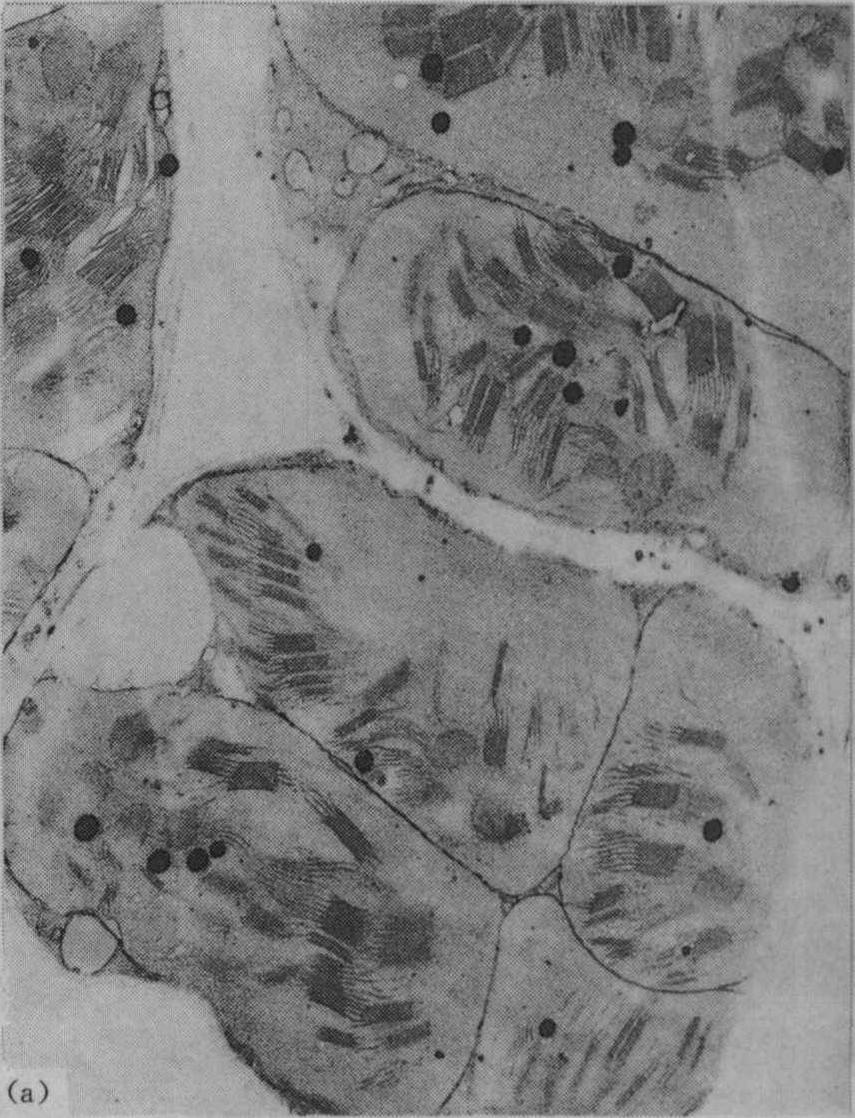
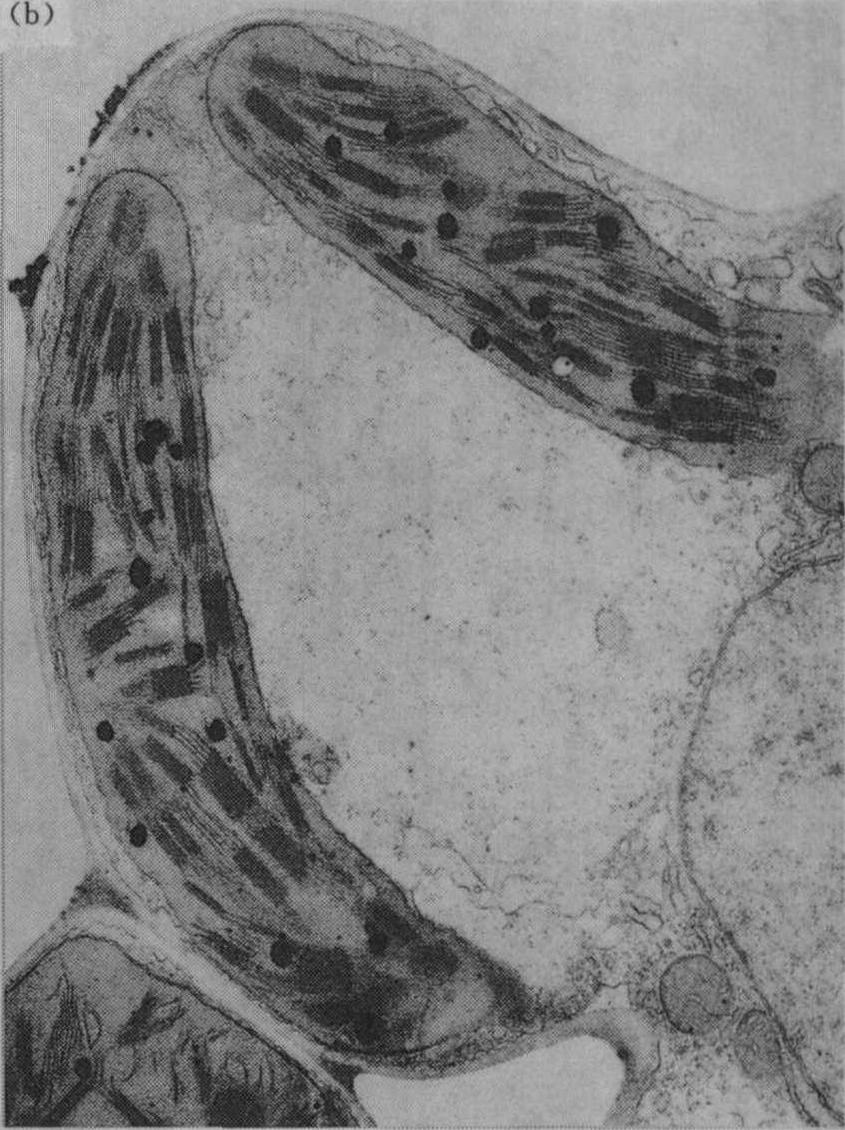
图1 盆栽水稻施钾与不施钾肥叶绿体的超微结构
(土壤中含速效钾25mg/kg)
(a)不施钾(K0)处理;
(b)施适量钾(K1)处理(4.5g/15kg)
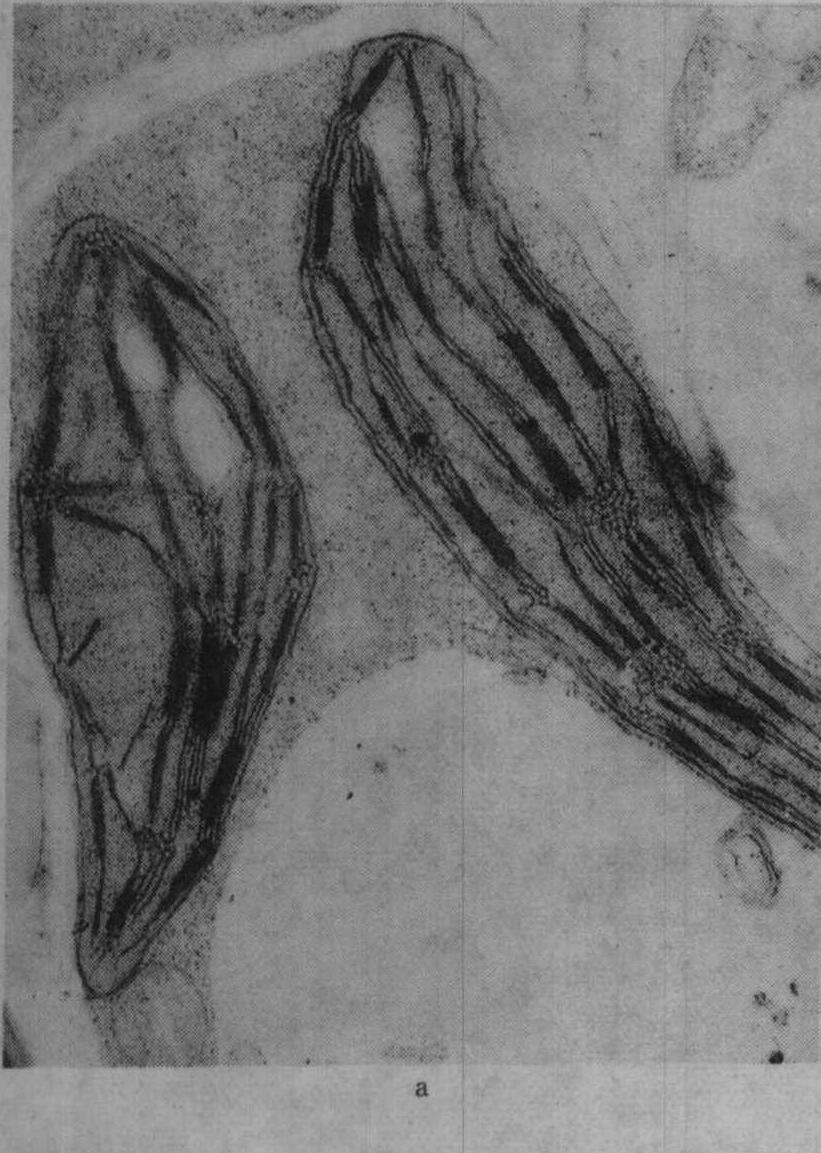
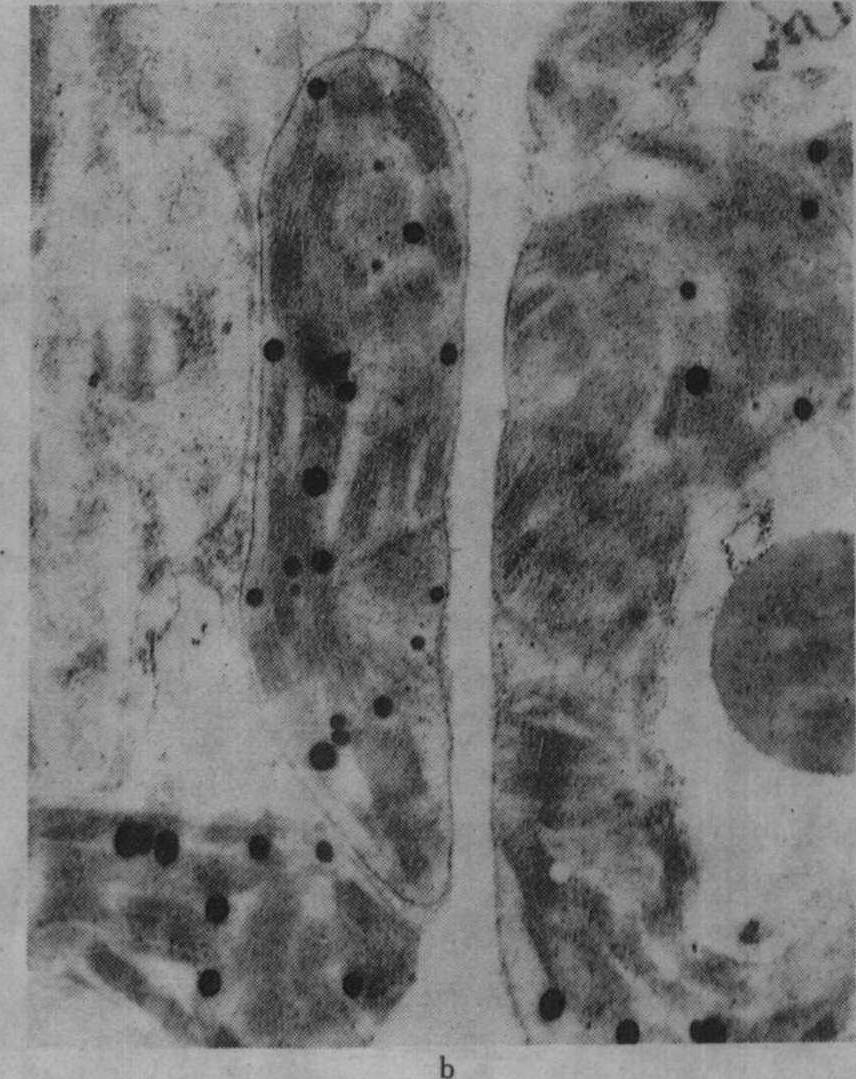
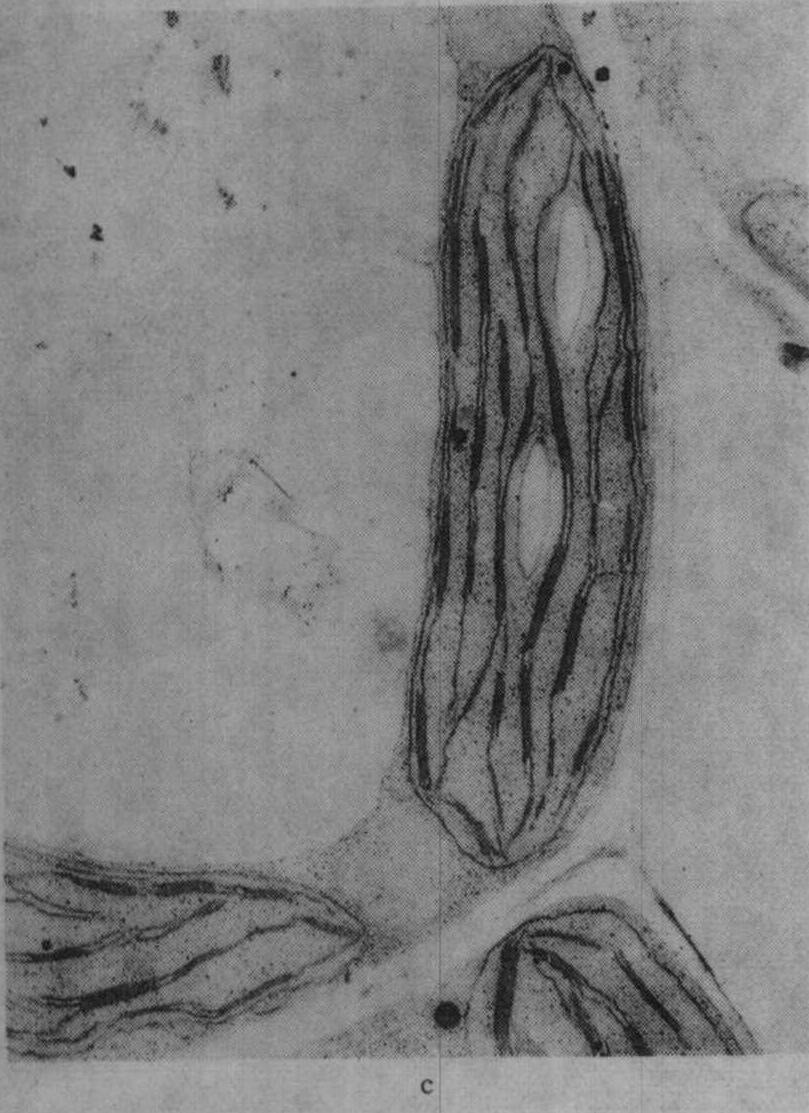
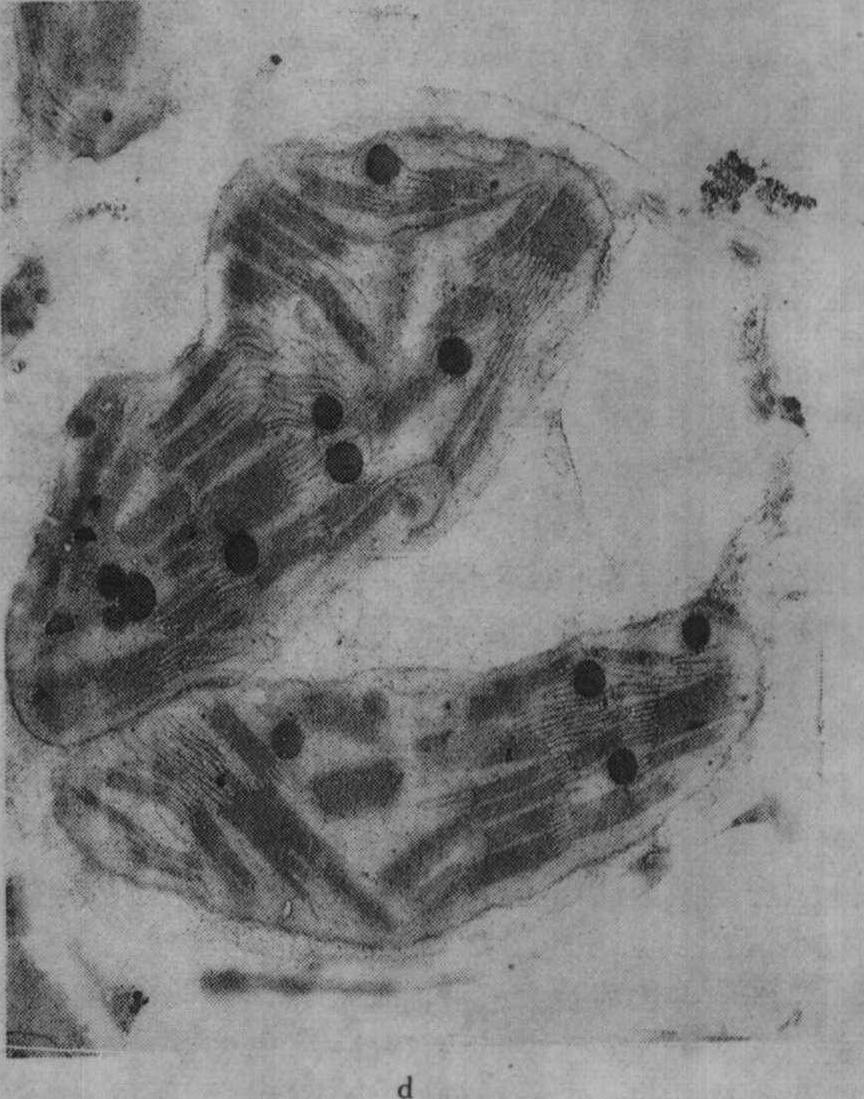
图2 不同钾水平水稻叶绿体的发育情况
(第10叶,放大15 000倍)
a.不施钾处理(K0)的叶绿体,针叶抽出后第2天;b.K0叶绿体,针叶抽出后第25天;
c.施钾处理(K1)的叶绿体,针叶抽出后第2天;d.K1叶绿体,针叶抽出后第25天
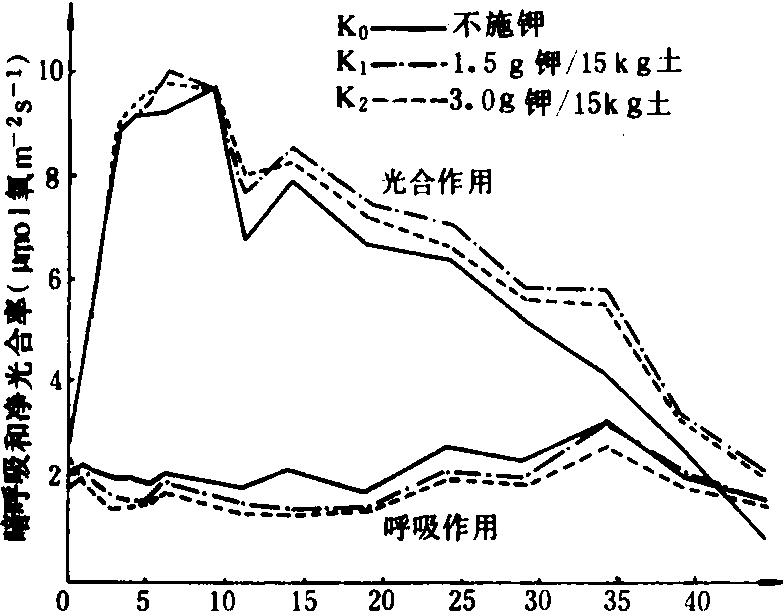
出叶后天数(d)
图3 水稻叶片发育过程中光合作用的动态变化
(杂交稻威优35,盆栽)
气孔开闭是保卫细胞及周围细胞间膨压平衡变化的结果。保卫细胞内K+浓度增高,水势降低,从相邻细胞吸水,使其膨压增高,气孔张开;气孔关闭的情况则反之。在气孔运动中,K+的渗透调节功能不能用其他阳离子代替。脱落酸(ABA)抑制气孔张开或诱导气孔迅速关闭,主要是由于ABA引起K+从保卫细胞外流量增加。光通过结合在保卫细胞质膜内的H+泵ATP酶的作用,诱导K+在保卫细胞内积累,因为ATP酶使ATP水解,从保卫细胞内泵出H+,与此同时,保卫细胞外的K+即进入,所需ATP由保卫细胞内叶绿体的光合磷酸化作用提供。
植物的感夜运动及感震运动也是膨压调节的运动,K+同样有关键性的作用。如合欢树,其羽状复叶在白天张开,夜间闭合,叶片的开闭是由于叶柄基部的叶枕膨压变化所引起。叶枕内的H+泵ATP酶在光下泵出H+,使K+进入,导致水分进入而引起膨压增高,叶片张开;黑暗中则由于K+外流引起其膨压降低而使叶片关闭。含羞草叶的感震运动也是由于叶枕膨压变化所引起,受到刺激时叶枕内的K+再分配,导致含水量及膨压改变,膨压降低时,叶闭合;膨压增高时,叶张开。
阴阳离子平衡及pH调节 K+无论在平衡细胞结构内大分子的阴离子电荷或是在液泡、木质部及韧皮部内可转移的阴离子电荷中都是最主要的阳离子,K+在细胞质及叶绿体内保持较其他部位为高的浓度,这是平衡无机及有机阴离子电荷所必需,同时也可保持这些部位的pH值在7~8,这是大部分酶促反应的最适pH值,使其中的酶类能保持较高的活性。例如,硝酸还原酶当pH值从7.7降至6.5时,其活性几乎完全被抑制。
细胞的质膜及液泡膜结合的H+—ATP酶泵出H+,同时K+即进入,保持细胞质一定的pH值,虽然其他几种一价阳离子在一定范围内可取代K+,例如NH+4,但它在高浓度下对细胞有毒害作用;Rb+的一些性质和K+相似,但自然界很少;Na+不能代替K+对酶的活化作用。
钾促进植物根对NO-3的吸收,并促进NO-3在植物体内的运输,K+是NO-3和苹果酸阴离子在植物体内运输过程中的反离子(counter ion),如图4所示。根吸收的NO-3从根运往地上部的过程中,K+可平衡其电荷;另一方面,地上部磷酸烯醇丙酮酸固定CO2形成草酰乙酸,后者还原形成苹果酸。苹果酸阴离子经韧皮部运往根系的过程中,K+也有平衡电荷的作用。在根内苹果酸脱羧产生的HCO-3释放到土壤溶液中时,NO-3则吸入根内,根内的K+又可在NO3 经木质部运入地上部过程中平衡其电荷,可见K+可促进NO-3的吸收和运转。
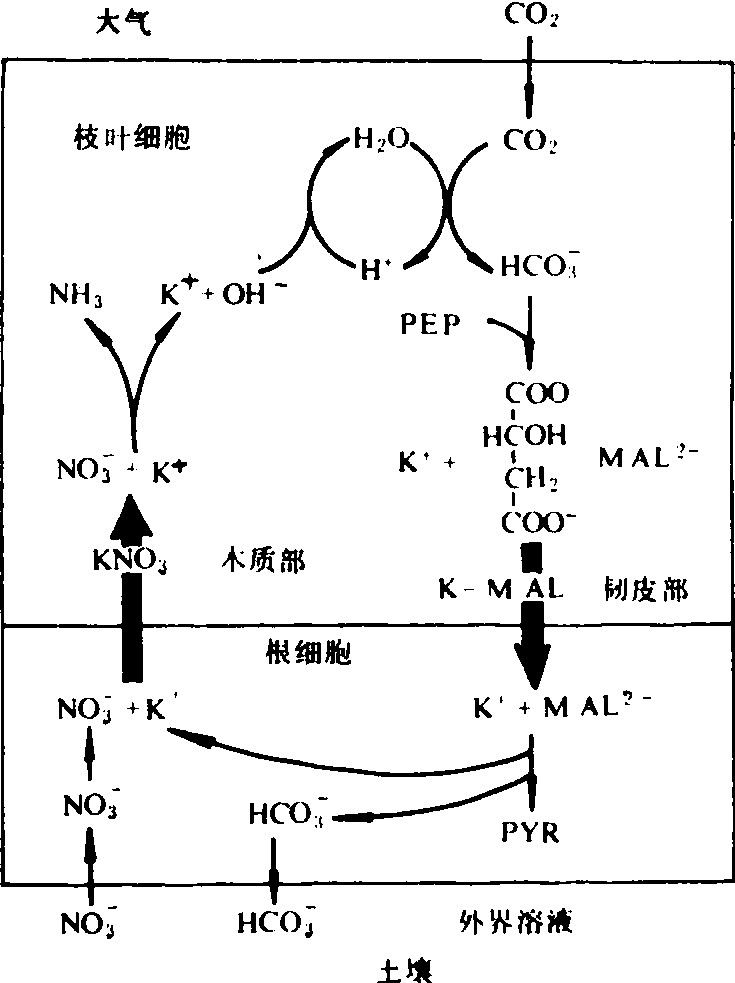
图4 K+促进NO3 的吸收和运转图解PEP 为磷酸烯醇丙酮酸阴离子;MAL2为苹果酸阴离子; PYR 为丙酮酸阴离子
蛋白质代谢 蛋白质含量较高的作物,其茎叶内亦含有较多的钾。收获种子的作物其种子中蛋白质含量较高的,钾含量也较高,二者间呈正相关(表2)。种子中的蛋白质体亦含有大量的钾。在植物体内蛋白质大量合成的分生组织,其含钾量亦较高。在体外试验中,维持蛋白质合成的最高速度需要K浓度为每升100~150毫摩尔,若K浓度低于此值,则蛋白质合成速度下降。
表2 几种作物种子中钾含量和蛋白质含量间的相关性
| 作物 | 种子中含K量(g/kg) | 粗蛋白含量(g/kg) |
| 大豆 菜豆 棉花 向日葵 大麦 燕麦 小麦 黑麦 高粱 玉米 | 17.7 13.8 12.0 7.5 5.5 4.8 4.7 5.2 3.9 3.3 | 380 253 231 179 126 121 120 113 110 90 |
| r=0.98 | ||
钾在植物蛋白质代谢中的功能为:❶促进根对NO3-的吸收与运转。在根吸收NO-3与NO-3向地上部运转的同时,K+与NO3-一同经木质部向枝叶运输;枝叶内的K+经韧皮部向根系运输时伴随着苹果酸阴离子的运输。苹果酸在根内脱羧产生的HCO-3从根释放至土壤溶液中,同时土壤溶液中的NO-3则与HCO-3交换,进入根内(见图4)。在木质部汁液中K+与NO3-的比值常为1:1左右,足见二者间上述相伴随的关系。
❷促进氨基酸向蛋白质合成部位运输。在小麦灌浆期用15N-氨基酸分别处理高钾及低钾植株,发现15N结合到高钾处理植株谷粒中的量明显高于低钾处理;用蓖麻试验结果亦说明钾促进氨基酸通过韧皮部的运输。
❸促进蛋白质合成。用苜蓿试验,将缺钾苜蓿植株叶在光下用不同浓度的KNO3预处理20小时。然后用14C-亮氨酸饲喂动物,测定14C-亮氨酸结合到叶内RuBP羧化酶/加氧酶(Rubisco)中的量,其结果如表3。从表3可见,在充分供钾的植株叶片中,14C-亮氨酸结合到Rubisco中的量最多,而缺钾叶片中,14C-亮氨酸结合到Rubisco则受到抑制。植物叶片的可溶性蛋白中,Rubisco的含量约占一半,这说明钾不足影响蛋白质合成。
表3 钾对14C-亮氨酸结合到缺钾苜蓿植株叶片的影响
| 预培育介质 (KNO3 mmol/L) | 14C-亮氨酸结合到Rubisco中的量 [dpm/(mg Rubisco·24h)] |
| 0.0 0.01 0.10 1.00 10.00 对照(K+充分植株) | 99 167 220 526 526 656 |
C4植物的Rubisco及总蛋白含量均较C3植物低,但C4植物中Rubisco及K+的利用效率均较C3植物高,因为C4植物光合作用中碳同化除具备C3途径外,还有C4途径,C4途径是在维管束鞘细胞内进行。其中有充分的CO2,故促进Rubisco催化羧化反应。C4植物K+的利用效率也较高,用牧草所做的试验报道表明,C4牧草K+的利用效率平均约68 500克干重/千克K+,C3牧草为41 700克干重/千克K+,说明消耗同样多的钾,C4植物能形成更多的干物质。
电荷平衡 K+可平衡酸性氨基酸中的负电荷,使蛋白质结构稳定。种子中大部分贮藏蛋白质富含酸性氨基酸谷氨酸、天门冬氨酸,K+可平衡其负电荷。含高蛋白质的种子内高浓度的K+可能也是平衡脱氧核糖核酸(DNA)及核糖核酸(RNA)电荷并活化蛋白质合成机构(protein synthesis apparatus)所必需。很多酶蛋白中含有一些酸性氨基酸,K+亦可平衡其负电荷,使酶稳定,不易破坏。
此外,K+还是天冬酰胺酶的活化剂,天门冬酰胺酶催化天冬酰胺水解,生成天门冬氨酸及NH3。天门冬酰胺和谷氨酰胺都是氮的贮藏形态,其水解产物又在蛋白质及氨基酸合成中被利用。
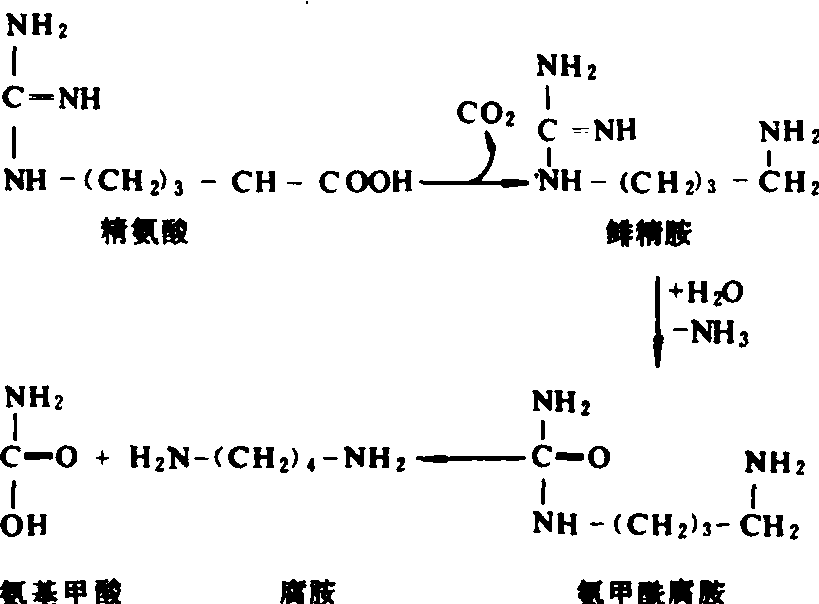
缺钾情况下蛋白质合成受阻,氨基酸、酰胺及NO-3等积累,氮代谢不正常,植物体内常形成有毒的胺,例如鲱精胺和腐胺。腐胺是氮代谢失调的产物,但在缺钾情况下,它对阴阳离子平衡及保持细胞一定的pH值方面也有作用。
植物生长与收获物品质 钾营养的充分供应是促进植物生长及提高收获物品质必需具备的条件。不论分生生长、伸长生长及输导组织形成都必需钾。分生组织是蛋白质代谢很活跃的部位,如缺钾则蛋白质代谢受抑制,导致分生生长受阻。细胞伸长与钾密切相关,K+是最重要的渗透溶质,使细胞能保持一定的膨压,这是细胞伸长的先决条件,同时细胞伸长还需要增加细胞壁的可塑性和延伸性。K+促进质膜结合的H+—ATP酶的作用。该酶从细胞质泵出H+到质外体,使质外体pH值降低,活化水解酶,细胞壁易延伸。吲哚乙酸(IAA)及赤霉酸(GA)都促进细胞伸长,K+与二者都有协合作用(synergistic effect)。
植物缺钾时膨压降低,在水分胁迫下易萎蔫,植物组织及细胞器常发育不正常,木质部及韧皮部形成较皮层组织受到更大的抑制,木质化受阻,维管束厚壁组织及角质层均不发达(图5)。禾谷类作物缺钾时硅化程度低,如图6所示,故叶片较软弱易披叶,植株易倒伏。
缺钾细胞膨压降低,细胞小,叶面积减小,整个植株的光合面积减小,影响产量形成。图7示植物在不同钾水平处理下的生长情况。缺钾的番茄果实中有较多脱落酸(ABA),易脱落,成熟果实中多各种病态果实。一般果实也较小,色较黄而不红,维生素C含量较少(表4),所以缺钾时,其产量和品质均降低。
表4 钾营养条件对番茄产量及品质的影响
| 处理 | 每株果数 | 每个果重 (g) | 果实产重 (g/株) | 果实中维生素C含量 (mg/100g,鲜重) |
| K0 K1 K2 | 17.3a 20.1a 20.8a | 35B 53A 55A | 590.4B 1107.6A 1120.8A | 20.65B 24.76A 25.96A |
表中数据为10次重复的平均值。
由于缺钾时光合产物运输受抑制,同时激素代谢亦受到影响,果实内ABA含量增高,库活性急剧下降,因而通过韧皮部运至果实及种子内的同化物及再利用的无机养分都明显减少,使种子活力降低,播种后出苗延迟,扎根不深。
钾对植物抗性的效应 由于钾促进蛋白质合成,施肥时,注意适当配合钾肥,可改变寄主及寄生物的环境,且调节代谢,在寄主体内积累抗毒化合物,故能减轻病害。钾使植物组织坚韧,不易倒伏。由于钾的渗透调节和代谢功能又使植物对高温及霜冻的抵抗力增强。
抗病害 高水平的钾及养分平衡可减轻大豆种子由于大豆黑点病(Diaporthesojae)引起的种子霉烂;马铃薯晚疫病(Phytophthora infestans)在只施氮肥和磷肥时较严重,增施钾肥即可减轻;禾谷类作物在高氮水平下易发生锈病(Puccinia spp.),增施钾肥亦可减轻; 高水平的钾亦可减轻烟草的野火病(Pseudomonas tabaci),并可缓解高氮的影响。
钾增加植物对病害的抵抗力主要是与其代谢功能有关,缺K植物由于蛋白质合成受抑制,在导管液内积累了低分子的含N物质,为病原菌生长提供了更好的条件。钾通过调节植物代谢亦可减轻病害,抗病性强的植物在感病部位的周围积累植物抗毒素(phytoalexins)、酚及生长素,阻止孢子发芽和菌丝生长,并使植物迅速形成愈伤组织,例如,钾肥提高烟草对感染花叶病(Nicotiana glutinosa)及葡萄对花腐病(Botrytis cinerea)的抵抗力即与K调节植物代谢,迅速形成愈伤组织有关。此外钾营养充分的植物组织坚韧,表皮上角质层发达,禾谷类作物硅化程度高,病菌也不易侵入。
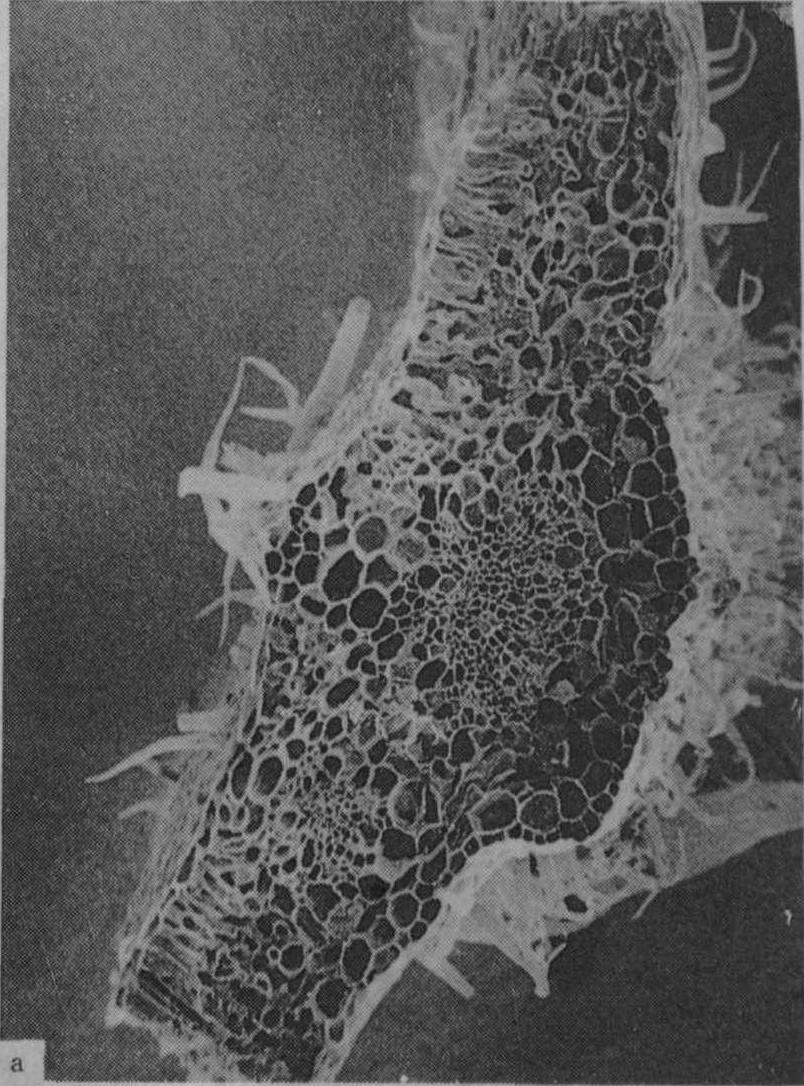
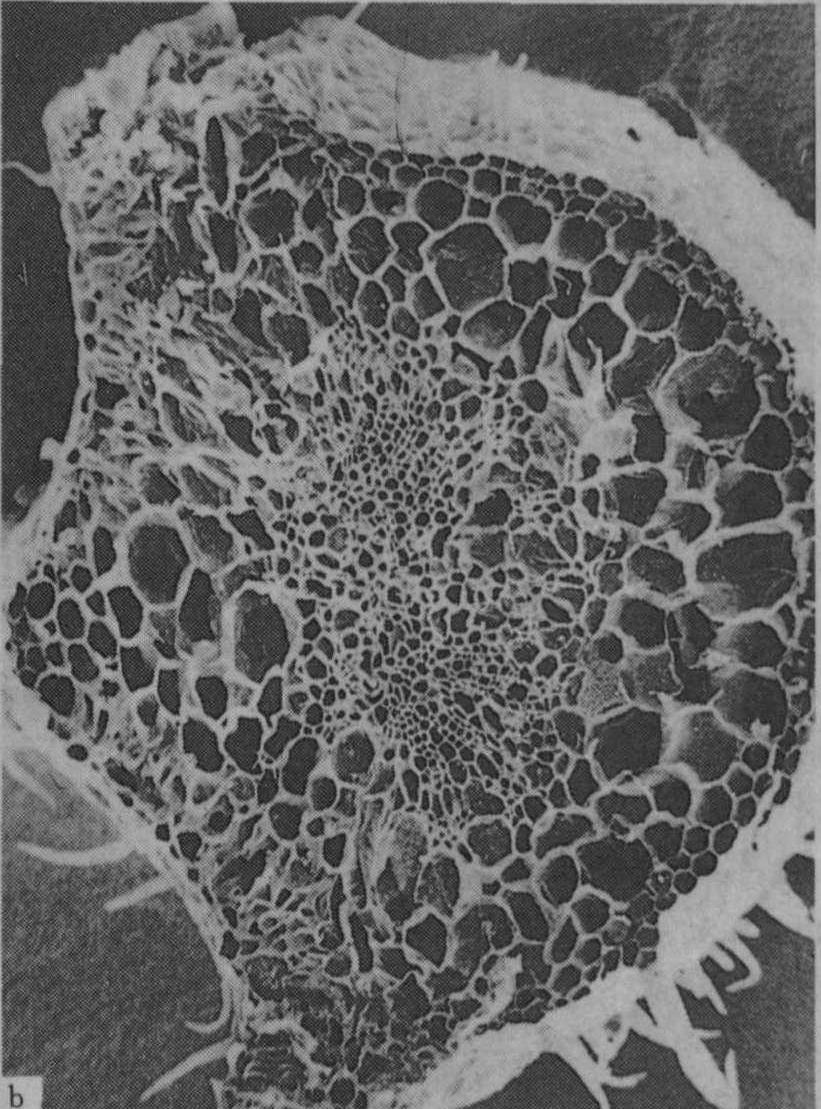
图5 番茄叶柄横切面在不同钾水平处理下维管束的发育情况
a. K0(盆栽,不施钾,土壤含速效钾25mg/kg);
b. K1(每5克土中施K2O 3.0g)


图6 水稻叶表面在不同钾水平处理下,叶片硅质及栓质化乳突情况
a.K0(不施钾,盆栽用土含速效钾25mg/kg);
b.K1(每15克土中施K2O 4.5g)
抗高温 缺钾植物在高温条件下,易失去水分平衡,引起萎蔫。棉花、丝瓜和南瓜等叶片较大的植物尤为明显,夏日中午缺K植株的叶片常萎蔫,影响光合作用的进行。K+有很好的渗透调节功能,供K良好的植物在高温条件下仍能保持较高的膨压,维持正常代谢进程。

图7 不同钾水平处理的番茄果实
左:K0的病态果实;右:K1果实
抗冰冻 冰冻对作物的损伤可能是由于细胞膜结构遭到破坏,引起溶质渗漏,甚至使代谢的区域化(compartmentation)受到影响。在土壤冻结时就影响植物水分及养分的吸收,根系发育差的植物也可能因土壤冻结而受到机械的损伤。钾营养充分且养分平衡的作物在温度逐渐降低的过程中,光合作用速率虽然也逐渐降低,但因低温下生长较慢,故积累同化物较多,细胞液内含糖量较高,组织内束缚水/自由水的比值增高,不易结冰,且膜结构亦较稳定。同时根系发育良好,活力亦较强,在低温下仍能从土壤中吸收水分和养分;在土壤冻结时不易受到机械损伤,故抗冰冻能力较强。
抗倒伏 钾营养良好的植物厚壁组织增厚;合成细胞壁组成物质较多,细胞壁较厚;表皮上角质层发达;禾谷类作物硅化程度增高,组织坚韧,茎秆强健,不易倒伏。
总之,充分的钾营养及养分平衡,促进植物蛋白质代谢及碳水化合物代谢,使植物生长强健,增强抗病能力及对不良气候条件的抵抗力,这对提高作物品质和产量都是有利的。
- 秦淮花柳添憔悴,玄武旌旗空寂寥。是什么意思
- 秦淮诗是什么意思
- 秦淮门是什么意思
- 秦淮风光带是什么意思
- 秦清芬是什么意思
- 秦清锡是什么意思
- 秦渔问是什么意思
- 秦渠是什么意思
- 秦渡大米面皮是什么意思
- 秦渡镇是什么意思
- 秦渭水渡是什么意思
- 秦游草是什么意思
- 秦游诗是什么意思
- 秦湛是什么意思
- 秦湛《卜算子》是什么意思
- 秦湛《卜算子春情》是什么意思
- 秦源是什么意思
- 秦源目断,碧云暮合,难认仙家。是什么意思
- 秦溪外史是什么意思
- 秦溪蒙难(四首) - 冒襄是什么意思
- 秦漢印型是什么意思
- 秦漢印存是什么意思
- 秦漢印章拾遺是什么意思
- 秦漢印範是什么意思
- 秦漢印簡是什么意思
- 秦漢印統是什么意思
- 秦漢子是什么意思
- 秦漢玉印圖録是什么意思
- 秦漢瓦當圖是什么意思
- 秦漢瓦當圖記是什么意思
- 秦漢瓦當文字是什么意思
- 秦漢金文録是什么意思
- 秦漢魏晋篆隸字形表是什么意思
- 秦潤卿是什么意思
- 秦濛生是什么意思
- 秦瀚是什么意思
- 秦瀛是什么意思
- 秦火是什么意思
- 秦火之劫是什么意思
- 秦火之后,三代制作湮灭几尽。汉时购书之赏重,故汉儒附会之书多。其幸存者,则焚书以前之宿儒尚存而不死,如伏生口授之类;好古之君子壁藏而石函,如《周礼》出于屋壁之类。后儒不考古今之文,概云先王制作而不敢易,即使尽属先王制作,然而议礼制度考文,沿世道民俗而调剂之,易姓受是什么意思
- 秦灭东周是什么意思
- 秦灭义渠是什么意思
- 秦灭二周是什么意思
- 秦灭六国是什么意思
- 秦灭大荔是什么意思
- 秦灭巴蜀是什么意思
- 秦灭梁是什么意思
- 秦灭楚之战是什么意思
- 秦灭燕代是什么意思
- 秦灭荡社之战是什么意思
- 秦灭蜀是什么意思
- 秦灭西周是什么意思
- 秦灭豲是什么意思
- 秦灭赵是什么意思
- 秦灭赵之战是什么意思
- 秦灭韩是什么意思
- 秦灭魏是什么意思
- 秦灭魏之役是什么意思
- 秦灭齐是什么意思
- 秦灭齐之战是什么意思