视觉过程的生物物理学
视觉是由对光敏感的组织(例如哺乳动物眼的视网膜)把光能转变成电能,把其中所含的信息从一种密码翻译成另一种密码。然后传递到大脑,引起感觉的过程。视觉过程的生物物理学着重研究下列问题: 视网膜中的视杆与视锥细胞对光量子的接收、视色素构型的改变与漂白、感受器电位的产生及其翻译成“全或无”性质的动作电位、视觉信息的传递与处理等。
1851年Muller首先描述了多种脊椎动物的视网膜呈淡红色,1876年德国科学家Boll分离了蛙的视网膜,发现经暗适应后的蛙眼曝光时,在视网膜中淡红色的视色素先变成黄色,最后无色(漂白)。1877年至1882年间,Kuhne等的一系列工作证明在视网膜中确实存在一种淡紫红色的色素,并称之为视紫红质。从二十世纪三十年代以来,视觉研究工作者揭示了许多有关光引起光感受器兴奋的机理。其中,Wald等对抽提的视色素研究,为视觉生理和生物物理的进展提供了必要的基础,因而在1967年获得诺贝尔奖金。另外一些视觉科学工作者则从电生理学的角度对视觉兴奋过程进行了较深入的探讨。早在一百多年前Holmgren就发现,当光刺激眼睛时,能从角膜表面引出一种电位,后来称之为视网膜电图(ERG)。本世纪三十年代Granit对ERG成分进行了系统的分析。尽管ERG是由光感受细胞和各类中间神经元电活动的总和所构成的,但人们也逐渐把ERG作为某些临床眼病的诊断指标。近十年来由于微电极技术的发展,不仅能对ERG各个成分的起源进行更深入的分析,而且能分别对视网膜各类细胞进行单个细胞电位引导,这对研究视觉信号如何在视网膜中进行信息处理提供了良好的基础。1972年Hagins提供了光感受细胞兴奋的Ca++离子学说,把光化学过程和电位变化的关系进一步联系起来。
视色素及其化学反应 光感受细胞(视杆和视锥细胞)的外层膜结构中,含有许多视色素,这是一种具有感光效应的物质,它由视蛋白和专门吸取可见光的生色团结合而成。这种生色团就是视黄醛,其结构为一共轭双键的链,两端分别连接一个紫萝酮环和一个醛基(图1)。视黄醛为维生素A的衍生物,它是由维生素A的醇基氧化为醛基而成。不同类型视感受细胞中的视色素是不相同的,例如视杆细胞中的视色素称为视紫红质,它是由视黄醛1(或称网膜醛)与杆视蛋白结合而成;而视锥细胞中的视色素则称为视紫蓝质,它是由视黄醛1与锥视蛋白结合而成,而有些动物(例如一些鱼类) 的视色素则由视黄醛2和视蛋白结合而成。视黄醛2与视黄醛1的区别在于前者的紫萝酮环上少了两个氢原子,多了一个双键。不同的视色素具有不同的吸收光谱峰值。
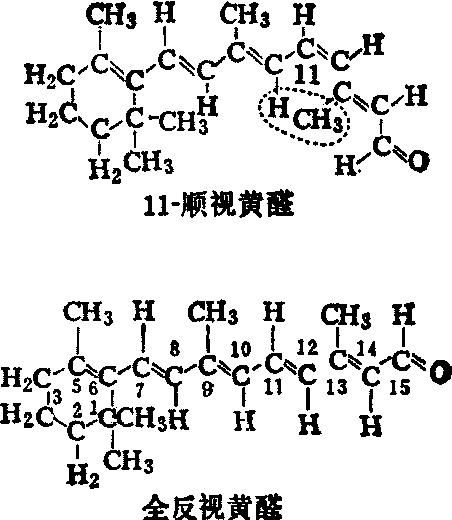
图1 11-顺视黄醛与全反视黄醛的结构
视紫红质分子吸收光量子时,其直接效应是使生色团发生异构化,从11-顺型转变成全反型,此后依次发生一系列的化学变化,并且不再需要光的作用,最终导致全反视黄醛与视蛋白分离,原基质的颜色随之消失,即所谓的漂白。溶液中视紫红质漂白过程顺序如图2所示。其动力学速率随温度和pH等实验条件而异。在漂白过程中,全反视黄醛脱离视蛋白后,还原成全反维生素A,扩散到色素上皮层,通过光或酶(包括异构酶)的作用,再转变成11-顺型维生素A,后者又氧化成11-顺视黄醛,通过席夫(Shiff)碱基键与视蛋白相连,最终又回到光感受细胞的外段中,此时又具有原来视紫红质的特性。
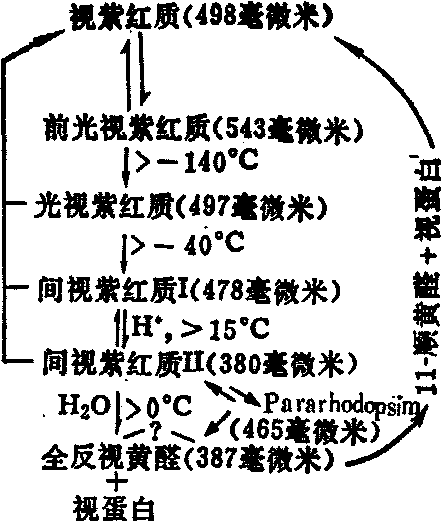
图2 视紫红质的漂白与光链反应
早期感受器电位 尽管一百多年以前,人们就曾从眼睛的角膜表面引出了视网膜电图,然而这种视网膜电图中最早出现的一个a波,则在光照数毫秒后才开始出现的,而且也就在这几毫秒的潜伏期内,前述视色素漂白过程的主要反应步骤都已完成,因此很难把光引起视色素的化学变化和感受细胞的电位变化联系起来加以考虑。1964年,人们发现如果用短的强闪光作刺激,立即能引导出一个较小的电反应,称为早期感受器电位(ERP),它几乎没有潜伏期,而且能持续数毫秒,这样它就填补了ERG开始前的时间间隙。ERP由一个角膜正相电位(R1)以及紧接着一个角膜负相电位(R2)组成,再接着则是ERG的Q波。现有充分的实验事实表明,ERP的产生与视色素的光化反应过程密切相关,例如R1的出现与间视紫红质Ⅰ的形成有关,而R2则与间视紫红质Ⅰ转变为间视紫红质Ⅱ相关联。
视觉发生器电位 所谓“发生器电位”是一种膜电位变化,其变化幅度与感受细胞所受的刺激强度有关,然而却又不呈线性关系。发生器电位能引起突触后细胞的神经纤维出现动作电位,发生器电位按刺激强弱来调制动作电位的频率。近年来利用微电极技术发现,脊椎动物视杆细胞有一个由Na+离子流维持的电流回路。在暗处,Na+离子自视杆细胞内段流出,流入外段,这就是所谓暗电流。光照时这种暗电流降低,引起视杆细胞外段超极化。超极化程度的大小,与刺激光强度有一定的关系,当然也是非线性关系。这种超极化电位一般称之为晚期感受器电位,或简称为感受器电位。ERG的Q波主要来源于晚期感受器电位,也有视网膜中其他类型细胞的电位参与。晚期感受器电位又可触发视网膜中下一级神经元的电位发生改变,直至调制视神经节细胞动作电位的发放。
关于晚期感受器电位产生的机理,或所谓视觉转换的机理,1972年美国学者Hagins提出了所谓光感受细胞兴奋的Ca++离子假说。这个假说的中心思想是钙起细胞内递质的作用,其要点如下:
❶在暗处,视杆细胞外段能把钙离子从小圆盘间空隙(在视杆外段表面膜之内,小圆盘之外),通过小圆盘膜,泵入到小圆盘内的空隙,并堆积小圆盘之内。这种泵并不直接依赖于视色素(视紫红质)的状态,而受小圆盘间空隙中钙离子浓度的调节,这种离子泵的运转需要有代谢能。有人认为这种能由三磷酸鸟苷(GTP)水解成二磷酸鸟苷(GDP)来提供。
❷在视色素光异构化反应后的一系列热转变过程的某阶段,在盘膜上产生一个Ca++能通过的孔,于是钙离子沿着电化学梯度从小圆盘内空隙漏到小圆盘间空隙。
❸钙离子在小圆盘空隙的原生质内扩散,起阻塞外段表面膜上孔道的作用,而这种孔在暗处通常是允许Na+离子在细胞膜内外自由通过的。Na+通道的阻塞则引起外段超极化。这一假说原则上既适应于脊椎动物视杆状细胞,也适应于视锥细胞。然而现在有一些学者认为,光感受细胞兴奋并非如此简单地直接依赖于Ca++离子,也有人主张Ca++离子的作用是激活磷酸二酯酶(PDE),PDE对环鸟苷酸(cGMP)的水解起催化作用,而cGMP的水解则又激活另外的酶,这种酶最终导致外段表面膜上钠通道开放。晚近也有人主张,在视觉转换过程中,视紫红质的漂白和PDE的激活之间主要是通过扩散以及这些物质的相互作用而产生的,他们认为并不存在钙或其他递质通过盘膜泵入小圆盘内的作用,因此在视紫红质漂白时,盘膜上并不形成Ca++能通过的孔道。而且有人也发现在没有游离Ca2+离子存在下,光也能导致cGMP水解。但这些作者的实验取材不很一致,因而也有可能得出不同的结果。总之,迄今有关视觉转变的机理,虽然提出了一些学说,但仍有待进一步加深研究。
感受器电位如何引起视网膜中其他细胞的激发,而把视觉信息经双极细胞以及神经节细胞等,传到更高级的中枢,直至大脑,这也是人们所关注的另一个问题。视网膜其他细胞的电活动 由于微电极技术的发展,结合运用震动插入装置,人们已经能将直径<0.5μm的微电极插到视网膜的各类细胞内引导电位,并用荧光黄一类的染料注入细胞内,使细胞标记着色,作形态学上的鉴定。除了上述光感受细胞外,对视网膜中其他各类细胞的电活动也进行了较广泛的观察:
(1) 双极细胞: 对于光刺激,双极细胞不产生动作电位而只产生梯级电反应。双极细胞的感受野是同心圆的形状,分中央部和外周部。用小光点刺激感受野的中央部与用环状光斑刺激感受野的外周部所引起的电位正好倒转。有两种双极细胞,一种是当光作用在它的感受野中央部时,出现超极化反应,称“超极化型”双极细胞,也称 “off-中央型” 双极细胞,另一种是光作用在它的感受野中央部时,出现去极化反应,称“去极化型”双极细胞,也称“on-中央型”双极细胞。双极细胞感受野中央部范围与该细胞树突所扩展的范围几乎一致,即与该双极细胞直接发生突触连接的光感受细胞则能使之产生超极化或去极化反应。然而它的外周部的范围,则大大超过了该细胞树突所覆盖的范围。因此光刺激双极细胞感受野外周部的光感受细胞时,则要通过另外的神经元(如水平细胞)才能引起双极细胞的电反应。水平细胞对光感受细胞又有负反馈作用。
(2) 水平细胞: 水平细胞也是与光感受细胞相连的第二级神经元。与前面两种细胞相似,对于光刺激,水平细胞也只产生梯级电反应。水平细胞的电反应被称之为S电位。S电位又可进一步分为两种,一种称为L电位,这是在任何波长的光刺激后都能出现的一种超极化反应;另一种是C电位,根据刺激光波长不同,既可出现超极化反应,也可出现去极化反应。S电位与感受器电位不同,它具有广泛的空间叠加效应,即用相同能量的大光斑作刺激比用小光点作刺激所引起的S电位大。然而对感受器电位而言,只要刺激光的能量相同(在相同波长下),所引起的感受器电位的大小相同,而与光斑的大小无关。(3) 无足细胞: 有两大类无足细胞,一类在给光或撤光时,出现暂时的去极化反应,另一类则产生动作电位。无足细胞的感受野,无中央部与外周部之分,当光刺激无足细胞感受野的任何部份时都出现类似的反应。然而无足细胞对刺激光强度的变化以及光点的移动非常敏感。故认为无足细胞对检测的明暗变化和物体的移动可能起一定的作用。
上述这些视网膜中不太长的细胞有可能借助某些递质的扩散或电流使原生质膜被动极化,将信息从细胞的一端传到另一端。而且有人认为都遵循如下的微分方程:
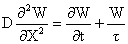
(4) 神经节细胞: 这是一类具有细长轴突 (组成视神经)的典型神经元,如果它也以梯级电反应来传导信息,则其电位必然会随传导距离延长而发生衰减,因此,毫无例外,神经节细胞都产生动作电位,利用动作电位这种神经系统的共同语言,把信息传递到更高级的神经中枢。神经节细胞电反应的形式是很不相同的,神经节细胞的感受野通常也可分为中央部与周边部。例如猫视网膜神经节细胞的感受野就是同心圆形的,并可分为“给光”(“on”)和“撤光”(“off”)两个区域。有一些神经节细胞,当光刺激其感受野中央部时,动作电位的发放增强,而光刺激其感受野的外周部时,动作电位发放抑制,这类神经节细胞称之为“on—中央型神经节细胞”。还有一些神经节细胞和上述情况相反,撤光时引起中央部动作电位的密度增加,称为“off—中央型神经节细胞”。
视网膜中各细胞的电反应需要通过细胞与细胞间的突触来传递。近年来一些学者的工作表明,视网膜内细胞与细胞间的突触传递,也要通过一些化学介质才能实现。其中目前工作开展得比较多的主要是光感受细胞与第二级神经元(即水平细胞与双极细胞)之间的突触传递。例如,有人指出,在暗处光感受细胞处于去极化状态,不断地释放出递质,这种递质作用于水平细胞的突触下膜,使之对Na+的通透性增高,于是水平细胞也处于去极化状态。光照使光感受细胞超极化,停止释放介质,于是水平细胞对Na+的通透性也下降,结果引起水平细胞超极化。前面曾经指出,视网膜电图(ERG)是视网膜各类细胞电活动的总和。二十世纪三十年代起,Granit以及其他视觉生理学工作者利用微电极记录局部ERG (LERG)、细胞内记录以及结合染色定位和阻断视网膜中央动脉血流等技术,对ERG主要成分的细胞起源,作了较细致的分析,其结果大致可归结如下:

b波——双极细胞(?)(Muller细胞?)
c波——色素上皮细胞。
从上述分析中可以看出在视觉系统中,视网膜是一个相当复杂的神经网络,在其中不仅要进行原初的视觉转换,而且也要进行初步的信息处理。不同种属动物视网膜的结构与功能的复杂程度也是不同的。例如蛙和鸽几乎没有视皮层,他们的视束纤维几乎都只进入视顶盖,其视网膜的结构和功能则比较复杂。而猫和猴等这一类较高等哺乳动物,由于它们都具有较发达的视皮层,其绝大多数的视束纤维都经过外侧膝状体中继,再进入大脑视皮层,然而它们的视网膜的结构和功能则比较简单。
研究视觉兴奋的生物物理过程,不仅是一个十分重要的基础理论问题,而且对临床诊断眼科疾患也提供了理论依据或有用手段。例如ERG不仅能作为反映视网膜功能的一个重要指标,用来研究视网膜的适应、颜色分辨、时间分辨等特性,而且在临床上也普遍地应用于视网膜疾病(如视网膜色素变性、视网膜剥离、视网膜中央静脉阻塞、铁锈症等)的诊断及疗效和预后分析。自从发现ERP以后,也有人试图把ERP用于临床诊断,例如有人指出,视网膜功能缺陷(如眼球结核)引不出ERP,而视网膜广泛损伤 (如特发性视网膜全剥离和原发性视网膜色素变性等)ERP消失或减少。尤其值得指出的是ERP主要反映光感受细胞外段的完整性和视色素的含量,而不涉及视网膜中其他神经细胞。例如单纯性视神经萎缩病人的ERP与正常人的基本相似,这既说明ERP可用作视网膜疾患鉴别诊断的手段; 也说明神经节细胞和视神经对ERP的产生无多大关系。另外,近年来还了解到,在正常情况下携带视觉信息的动作电位,经视神经传导和低级中枢(如外侧膝状体)中继,最后刺激大脑视区不同的神经元,使人们产生了丰富多彩的视觉。与此同时,随着脑外科技术的迅速发展,有可能用电脉冲(相当于动作电位)直接刺激距状裂附近的大脑皮层或枕叶,而不用经过视网膜和外侧膝状体等,就能使病人看见光点。人们利用这些科学成果和原理,已为盲人制作了直接刺激大脑视区而产生人工视觉的助视器。
- 秦陵铜鼎是什么意思
- 秦陵防水堤是什么意思
- 秦陵陶水道管是什么意思
- 秦陵马厩坑陶文是什么意思
- 秦陶文是什么意思
- 秦陶瓦是什么意思
- 秦陶窑是什么意思
- 秦陶窑遗址是什么意思
- 秦陶管是什么意思
- 秦陶量是什么意思
- 秦隋钱币是什么意思
- 秦隐君诗集是什么意思
- 秦隶是什么意思
- 秦隶书是什么意思
- 秦难见闻记是什么意思
- 秦雍城是什么意思
- 秦雍城遗址是什么意思
- 秦雪梅吊孝三元记是什么意思
- 秦雪玲是什么意思
- 秦雷是什么意思
- 秦震是什么意思
- 秦震鈞是什么意思
- 秦霑是什么意思
- 秦霖是什么意思
- 秦霸西戎是什么意思
- 秦青是什么意思
- 秦青川是什么意思
- 秦青韩娥是什么意思
- 秦靖是什么意思
- 秦靖军是什么意思
- 秦非是什么意思
- 秦非子是什么意思
- 秦韩是什么意思
- 秦韩围梁是什么意思
- 秦韬玉是什么意思
- 秦韬玉·贫女是什么意思
- 秦韬玉《贫女》是什么意思
- 秦韬玉的诗,秦韬玉的诗集,秦韬玉的古诗词大全秦韬玉是什么意思
- 秦韬玉诗集是什么意思
- 秦韶华是什么意思
- 秦风是什么意思
- 秦风·无衣是什么意思
- 秦风·蒹葭是什么意思
- 秦风谜刊是什么意思
- 秦香莲是什么意思
- 秦馨菱是什么意思
- 秦骃玉牍是什么意思
- 秦骊山汤遗址是什么意思
- 秦高奴簋是什么意思
- 秦髯是什么意思
- 秦鬟妆镜是什么意思
- 秦魏家墓地是什么意思
- 秦魏攻齐是什么意思
- 秦鱼池是什么意思
- 秦鲁一是什么意思
- 秦鳌是什么意思
- 秦鸡是什么意思
- 秦鸣雷是什么意思
- 秦鸣鹤是什么意思
- 秦鸿是什么意思