血红蛋白与气体运输
血红蛋白与气体运输
血红蛋白是血液运输氧和部分二氧化碳的载体,能将吸入的氧自肺部输送至各组织细胞,并将CO2自组织细胞运送至肺部而被呼出。
血红蛋白与氧的运输 血液中98%的氧是以与血红蛋白结合的形式被运输的。
氧合血红蛋白 血红蛋白中铁原子(Fe++)的第六配位键可与氧结合形成氧合血红蛋白 (HbO2),一分子血红蛋白可结合四分子氧。血红蛋白与氧的结合受氧分压的影响,反应如下。
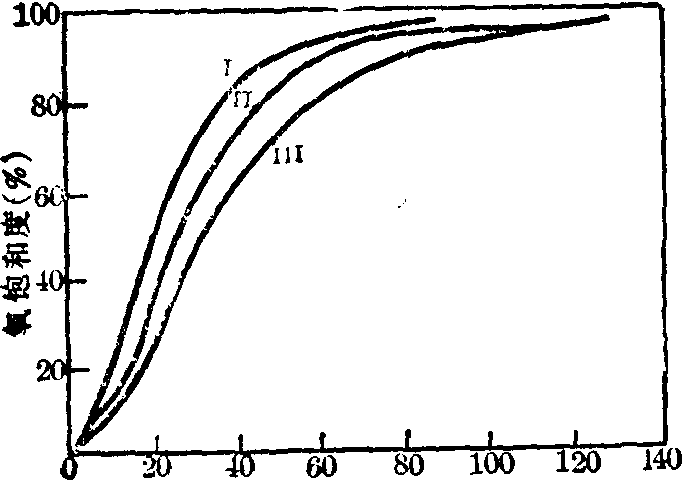
血红蛋白的氧解离曲线 按上式以血红蛋白的氧饱和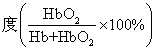 为纵座标,氧分压为横座标作图可得血红蛋白的氧解离曲线。血红蛋白的氧解离曲线呈特征性的S形(图1曲线Ⅰ),曲线的两端斜率较小,中段斜率较大。
为纵座标,氧分压为横座标作图可得血红蛋白的氧解离曲线。血红蛋白的氧解离曲线呈特征性的S形(图1曲线Ⅰ),曲线的两端斜率较小,中段斜率较大。
Po2(mmHg)
图1 pH与PCO2对血红蛋白氧解离曲线的影响
I pH7.6 PCO2 3.5kPa (25.5mmHg)
II pH7.4 PCO2 5.3kPa (39 7mmHg)
III PH7.2 PCO2 8.2kPa (61.3mmHg)
血红蛋白的S形氧解离曲线具有重要的生理意义。S形曲线的上部较平坦,因此当血液流经氧分压较高的肺部时,即使氧分压有些低落(例如高山空气稀薄),血液的氧饱和度也不致受很大的影响,仍能保证足够的氧合血红蛋白。S形曲线的中段较陡直,因此当血液流经氧分压较低的组织时,即使氧分压仅有较小的变化,氧合血红蛋白的解离却有明显的增加,从而释出较多的氧以保证组织的需要。
血红蛋白的S形氧解离曲线是由于血红蛋白分子中四个亚基与氧的结合存在着协同效应。当血红蛋白分子中的四个血红素铁依次与氧结合时,其结合常数各不相同,随着所结合氧分子数的增多,其与氧的亲和力也随之增高。Perutz等应用X光衍射晶体学分析发现脱氧血红蛋白与氧合血红蛋白的构象是不同的。脱氧血红蛋白呈“紧张态”T构象,而氧合血红蛋白则呈“松弛态”R构象。T构象时,四个亚基间存在着较多的盐键,其α亚基C末端精氨酸的羧基与胍基分别可与另一α亚基N末端缬氨酸的氨基以及第126位天门冬氨酸的β羧基形成两对盐键; β亚基C末端组氨酸的羧基也可与α亚基第40位赖氨酸的ε氨基形成一对盐酸; 同时,β亚基C末端组氨酸的咪唑基还可与同一β亚基FG转角处第94位门冬氨酸的β羧基形成另一对盐键。上述四对盐键的存在,使整个分子受到很大约束,绷得很紧,不易与氧结合。在R构象中不存在盐键,因此分子结构较松弛,与氧的亲合力增大。
目前认为,氧合过程中血红素铁从高自旋状态转变成低自旋状态是血红蛋白从T构象转变成R构象的触发因素。在脱氧血红蛋白亚基中,血红素铁(Fe2+)价电子层的3d轨道上原有四个未成对电子,呈高自旋状态,其直径较大,不能全部嵌入卟啉环平面,而突出于卟啉环平面靠近近心组氨酸(F8组氨酸)的一侧,离开卟啉环平面0.06nm处。氧合过程使孤对电子获得了电子,铁原子转入低自旋状态,铁原子直径缩小0.017nm,落入卟啉环平面中央的小孔,发生0.06nm的位移,并带动了与之相连的F8组氨酸,将后者拉向卟啉环平面。F8组氨酸的位移,引起亚基构象的变化,F肽段作顺时针向旋转,并向H肽段靠拢,FH间隙因而变窄,原来在FH间隙中的C端第2位酪氨酸的侧链被挤出间隙,酪氨酸的酚基与FG5缬氨酸的羰基间的氢链亦随之断裂。酪氨酸的位移牵动了C末端氨基酸,使之发生位移。由该C末端氨基酸所形成的亚基间的盐键因而断裂。因此,当一个亚基氧合时可引起α亚基间两个盐键断裂,脱氧血红蛋白分子中的约束力逐步减弱,从而促使其它亚基与氧的亲和力增强,容易与氧结合。当β亚基也氧合时,αβ间的以及β亚基内部的盐键也相继被打开。当亚基间的四对盐键全部断裂时,T构象即转变为氧合血红蛋白的R构象,完成了氧合过程。
血红蛋白氧合时由T构象向R构象的转变提供了解释血红蛋白的协同效应或S形氧解离曲线的理论基础。在氧分压低时,主要是T构象的脱氧血红蛋白,它与氧的亲和力很弱,曲线的斜率小; 在氧分压高时,大部分亚基均已氧合呈R构象,氧分压不再影响与氧的亲和力,曲线又趋向平坦; 曲线中部则是T构象转变为R构象阶段,由于氧亲和力的影响易化了HbO2的形成。
影响血红蛋白与氧的亲和力的因素 血红蛋白与氧的亲和力常以P50表示。P50是指血红蛋白的氧饱和度达50%时所需的氧分压。在不同条件下,血红蛋白与氧的亲和力有所不同,当亲和力降低时,氧解离曲线右移,P50值增大; 而当血红蛋白与氧的亲和力增高时,氧解离曲线左移,P50减小(图)。血红蛋白的氧亲和力可受pH、CO2分压(PCO2)、温度以及红细胞中2,3-二磷酸甘油酸(2,3-DPG)的浓度等很多因素的影响。
(1) pH和PCO2对血红蛋白氧亲和力的影响(图1):H+浓度增高时(pH由7.4降至7.2),血红蛋白与氧的亲和力降低,氧解离曲线右移,P50增大; 而H+浓度降低时(pH由7.4增至7.6)则P50减小。H+浓度能影响血红蛋白的构象,已知T构象的血红蛋白分子的亚基间存在着四对盐键,H+浓度的增加有利于带正电荷的基团尤其是α链N端缬氨酸的氨基以及β链C末端组氢酸的咪唑基形成盐键,使血红蛋白稳定在T构象,因此血红蛋白与氧的亲和力降低,氧解离曲线右移。相反,H+浓度的降低不利于这些基团形成盐键,因而血红蛋白易转变成R构象,易于形成氧合血红蛋白。
PCO2对血红蛋白与氧亲和力的影响基本上与pH相似。PCO2增高能使血红蛋白的氧亲和力下降,氧解离曲线右移,P50增大。因CO2形成的碳酸以及与血红蛋白亚基末端氨基结合而成的氨基甲酸血红蛋白均能离解释出H+而使pH下降。从而影响血红蛋白与氧的亲和力。同时,氨基甲酸血红蛋白离解后形成的带有负电荷的氨基甲酰基还可与处于两β亚基间带正电荷的第82位赖氨酸的ε氨基形成盐,因而使两β亚基间的正电相斥力减弱,血红蛋白稳定于T构象,血红蛋白与氧的亲和力降低。
上述H+浓度改变血红蛋白与氧的亲和力的现象称为Bohr效应。Bohr效应具有重要的生理意义。当血液流经组织时,由于细胞的pH较血液低而PCO2则较血液高,有利于氧合血红蛋白氧的释放,因而使细胞获得较单纯氧分压降低更多的氧。
(2) 2,3二磷酸甘油酸(2,3-DPG)的浓度对血红蛋白与氧亲和力的影响: 2,3-DPG是红细胞中糖酵解支路(2,3-DPG支路)的产物。此分子中的羧基与磷酸根均能解离,因此带有多量的负电荷。正常红细胞中2,3-DPG的mol浓度约与血红蛋白相同,能与脱氧血红蛋白以1:1的分子比值特异地结合而使血红蛋白稳定于T构象。因此红细胞中2,3-DPG的浓度是调节血红蛋白与氧亲和力的重要因素。
呈T构象的脱氧血红蛋白分子中,两β亚基间有一裂隙,其大小与2,3-DPG分子相当。裂隙的两侧壁有很多带正电荷的基团,包括β链N末端缬氨酸的氨基,82位赖氨酸的ε氨基以及143位组氨酸的咪唑基,它们正好可与2,3-DPG中带负电荷的羧基或磷酸根以盐键相连(图2)。
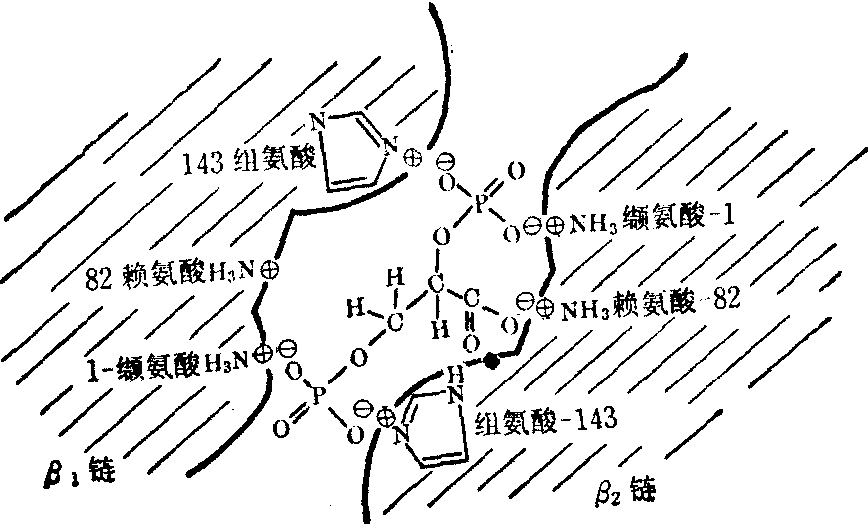
图3 血红蛋白与2,3-DPG的结合
T构象由于在原有四对盐键的基础上又增加了上述三对盐键,因此构象更趋稳定,而使血红蛋白和氧的亲和力下降,氧解离曲线右移。2,3-DPG使氧解离曲线右移的作用在氧分压较低时更为明显,而在氧分压高时则影响不大,因此当血液通过肺部时,由于氧分压较高。血红蛋白摄氧的能力受2,3-DPG的影响不大,但当血液流经组织时,红细胞中2,3-DPG的存在却能显著地增加氧的释放以供组织需要。
此外,由于2,3-DPG是一种不能透过红细胞膜的有机酸,因此也可通过使红细胞的pH下降而影响血红蛋白与氧的亲和力。当红细胞中2,3-DPG的浓度较高,超过它和血红蛋白分子结合所需的浓度而以游离的形式存在时,此机理就显得更重要。
由上可知,红细胞中2,3-DPG的浓度对血红蛋白与氧的亲和力有调节功能,它的浓度又受很多因素影响,2,3-DPG是糖酵解支路的产物,其浓度必然受糖酵解速度影响。无机磷酸盐因能加速糖酵解; 因此也能促进2,3-DPG的合成。脱氧血红蛋白因能和2,3-DPG结合,也能间接地调节红细胞中2,3-DPG的浓度,使其合成增速。一些生理或病理性的缺氧常可使2,3-DPG的浓度增高,而酸中毒则可使之降低。因此这些因素均能间接地影响血红蛋白与氧的亲和力,引起氧解离曲线移位以及P50的变化。
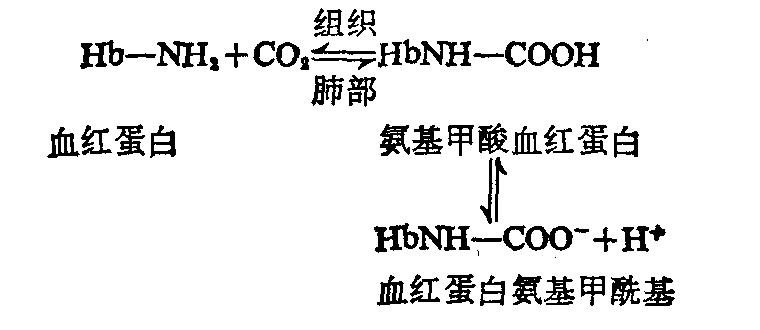
血红蛋白与二氧化碳的运输 血红蛋白可与CO2结合形成氨基甲酸血红蛋白,其结合部位是在α与β亚基的末端氨基。血红蛋白因此也是运输CO2的载体。血液中的CO2,约有1/4是以氨基甲酸血红蛋白的形式被运输的。
此外,红细胞中脱氧血红蛋白和其钾盐(KHb/HHb)以及氧合血红蛋白和其钾盐(KHbO2/HHbO2)所组成的缓冲体系又可对CO2在运输过程中所形成的碳酸起缓冲作用,因此使血液的pH不致发生太大的变动。
当血液流经组织时,组织细胞代谢产生的CO2进入血液,由于红细胞中PCO2低于血浆,故CO2不断由血浆弥散进入红细胞。红细胞中的碳酸酐酶促成碳酸产生,后者解离释出H+。H+浓度的增高有利于血红蛋白亚基间盐酸的形成,使血红蛋白由R构象转变成T构象,氧亲和力因而降低,氧合血红蛋白解离释出O2而转变成脱氧血红蛋白。同时组织中的氧分压较低,PCO2较高也有利于HHbO2的解离形成脱氧血红蛋白。由于分子构象的改变,脱氧血红蛋白的酸性较氧合血红蛋白弱,结合H+的能力增强,故能从介质中摄取H+,中和碳酸的酸性。上述变化过程可表示如下:
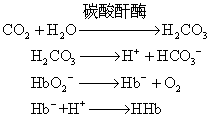 由此可见,氧合血红蛋白的脱氧是血液能大量携带CO2而不致显著降低pH的重要因素。同时由于碳酸释出的H+不断被脱氧血红蛋白所中和,因此进一步促进了CO2与H2O结合形成碳酸,有利于血液自组织摄取CO2。
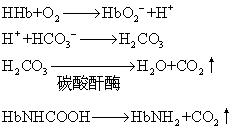
由此可见,氧合血红蛋白的脱氧是血液能大量携带CO2而不致显著降低pH的重要因素。同时由于碳酸释出的H+不断被脱氧血红蛋白所中和,因此进一步促进了CO2与H2O结合形成碳酸,有利于血液自组织摄取CO2。当血液流经肺部时,由于肺部的PCO2低于静脉血,血浆中物理溶解的CO2首先弥散至肺而被呼出。红细胞中由碳酸以及氨基甲酸血红蛋白分解释出的CO2亦随之经血浆弥散至肺泡呼出。但红细胞内碳酸的减少却并不引起pH的明显增加。这是因为当血液流经肺部时,由于肺泡的氧合血红蛋白而使酸性增加,解离释出H+。H+可与来自血浆的碳酸氢根结合生成碳酸,不断补充碳酸的消耗。其变化过程如下:
☚ 血红蛋白 血红素的分解——胆红素代谢 ☛
- 道济是什么意思
- 道济是什么意思
- 道济(1)是什么意思
- 道济(2)是什么意思
- 道济垫款问题是什么意思
- 道济长城是什么意思
- 道济颠狂是什么意思
- 道深昙愍等造像记是什么意思
- 道清铁路工会是什么意思
- 道溥是什么意思
- 道滕代伊是什么意思
- 道潜是什么意思
- 道潜是什么意思
- 道潜是什么意思
- 道潜是什么意思
- 道澄是什么意思
- 道熙是什么意思
- 道独是什么意思
- 道狭草木长,夕露沾我衣。是什么意思
- 道猛是什么意思
- 道理 理是什么意思
- 道璨是什么意思
- 道璿是什么意思
- 道生是什么意思
- 道生是什么意思
- 道生是什么意思
- 道生是什么意思
- 道生是什么意思
- 道由白云尽,春与青溪长。是什么意思
- 道登是什么意思
- 道盛是什么意思
- 道盛是什么意思
- 道真仡佬族苗族自治县是什么意思
- 道真仡佬族苗族自治县概况是什么意思
- 道真仡佬族苗族自治县自治条例是什么意思
- 道祖是什么意思
- 道立于两是什么意思
- 道章是什么意思
- 道端等三百人造像记是什么意思
- 道纪司是什么意思
- 道经总论是什么意思
- 道统是什么意思
- 道统是什么意思
- 道统是什么意思
- 道统是什么意思
- 道统是什么意思
- 道统传经是什么意思
- 道统录是什么意思
- 道绰是什么意思
- 道绰是什么意思
- 道绰是什么意思
- 道者是什么意思
- 道者天下当然之理是什么意思
- 道者所以充形也是什么意思
- 道者无形,平和而神是什么意思
- 道者,令民与上同意也,故可以与之死,可以与之生,而不畏危是什么意思
- 道者,天地之始,一其纪❶ 也。物之所造,天之所生,包宏❷ ,无形❸ ,化气,先天地而成。莫见其形,莫知其名,谓之“神灵”。故道者,神明之源,一其化端❹ 。是以德养五气❺ ,心能得一❻ ,乃有其术。是什么意思
- 道者,德之本也是什么意思
- 道者,无之称也是什么意思
- 道者,福之本是什么意思