细菌代谢的遗传控制
细菌代谢的遗传控制主要是指调控基因在代谢调节过程中的作用。代谢调节过程可分为转录控制、转译控制及反馈抑制。
对于环境的适应是生物最基本的属性之一。有的适应针对比较固定的环境,有的针对随时改变的环境,后者主要通过代谢调节来实现。代谢调节的遗传控制的生物学意义并不限于适应; 通过代谢调节,基因型相同的细胞发育成不同的表型,所以代谢调节的研究也涉及生物学中另一基本问题,即个体发育中的组织分化问题。酵母菌在糖发酵中的适应现象是首先受到注意的一种代谢调节现象。四十年代对酶的诱导的生理学研究较多。五十年代开展细菌的遗传学研究后,才对大肠杆菌的乳糖发酵中酶的诱导现象进行了深入的遗传学研究,终于在1961年提出了乳糖操纵子模型,开辟了代谢调节的遗传学研究的广阔天地。到目前为止,对于真核生物的基因调控问题虽然提出了一些假设,但是都还缺乏足够的实验证据。
转录控制 代谢调节首先在转录的水平上进行。作用于转录的代谢调节系统如下。
负控制代谢调节 凡是某一细胞成分的存在使某种细胞功能不能实现,而这一成分的消失或失活使这一功能得以实现,这种控制称为负控制。在这里,酶的停止合成是由于调节基因的产物——阻遏蛋白——作用的结果。当阻遏蛋白由于和诱导物相结合或由于没有和合成代谢的最终产物结合时,它就失去活性,不再能和操纵基因相结合,酶的合成便能进行。属于负控制范畴的典型例子是乳糖操纵子模型和色氨酸操纵子模型。
(1) 乳糖操纵子模型: 大肠杆菌发酵乳糖是由两种酶的作用,即β-半乳糖苷透酶和β-半乳糖苷酶。前者负责乳糖的吸收,后者负责乳糖的分解。这两个基因(Y和Z)的位置是邻接的。在用甘油作为唯一碳源的培养基上培养的大肠杆菌细胞中,这两种酶几乎都不存在。在含有乳糖的培养基上培养的细胞中,这两种酶则大量出现。这是一种对于随时改变的环境的适应。曾经得到两种失去适应能力的突变型。这些突变型称为组成型,因为这两种酶不经诱导就能合成; 可是酶的性质和野生型 (诱导型)没有不同,可见这些突变型并不是编码蛋白质的基因发生突变的结果。上述两种突变基因中,有一种突变基因的位置并不和编码有关的两种蛋白质的结构基因相连锁,是隐性的、多效的,即一个基因发生突变时两种酶都由诱导型变为组成型。这是调节基因(i)发生突变的组成型(i-)。另一种突变基因的位置和结构基因Y和Z紧密连锁,是显性的,也是多效的。这是操纵基因(O)发生突变的组成型(Oc)。除了以上这些特性以外,突变型Oc和i-还有另一重要区别,就是位置效应的有无: 调节基因发生突变时,不论在同一染色体上的结构基因或是带在F因子上的结构基因,都同样受到影响而使酶的合成由诱导型变为组成型;而操纵基因发生突变时,只使同一染色体上的结构基因受到影响。根据这些实验结果,Jacob和Monod在1961年提出了乳糖操纵子模型。乳糖操纵子是由一个操纵基因和三个结构基因(即基因Y和Z,还有一个编码β-半乳糖苷转乙酰酶的结构基因A,这酶的生理功能还不了解)所构成的功能单位,当操纵基因同调节基因产物——阻遏蛋白相结合时,转录便停止进行。阻遏蛋白能同乳糖等诱导物相结合,这种结合可以使阻遏蛋白改变构型,从而脱离操纵基因,于是转录得以进行(图1)。
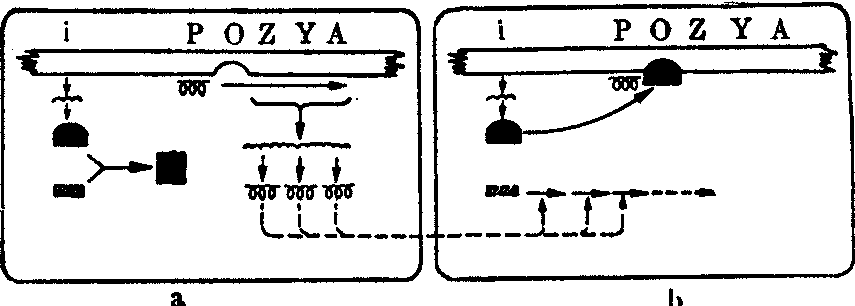
图1 乳糖操纵子在乳糖发酵中的代谢调节功能示意图
a: 乳糖存在时,乳糖和阻遏蛋白相结合,改变构型的阻遏蛋白不和操纵基因O结合,结合在启动区p的RNA聚合酶活动通过转录形成多基因mRNA,再通过转译合成三种酶
b: 乳糖在这些酶的作用下降解为H2O和CO2,阻遏蛋白作用于操纵基因使转录不能进行,于是酶的合成便停止

作为一个功能单位,乳糖操纵子还可以从以下三个方面说明它是一个整体:
❶协同诱导。除了乳糖以外,许多半乳糖苷能诱导β-半乳糖苷透酶和β-半乳糖苷酶的合成,不过各种结构类似物的诱导能力各不相同。实验结果说明不论是用什么物质诱导,两种酶总是按一定比例合成;
❷一个操纵子一个mRNA。在诱导物的作用下,通过转录产生包括三个结构基因的一个多基因mRNA,在转译过程中才分别合成三种酶,这是协同诱导的物质基础。
❸极性突变。距离操纵基因较近的结构基因如果发生一个终止密码子或移码突变时,非但这一种酶的活性不再出现,而且还影响距离操纵基因较远的结构基因,使这些蛋白质的合成速率也相应地下降。一个操纵子除了包括以上各部分以外,以后发现还包括一个mRNA聚合酶结合的位置,这位置称为启动区。
(2) 色氨酸操纵子模型: 大肠杆菌在含色氨酸的培养基中培养时,细胞中和色氨酸合成有关的酶就显著减少,这就是说,酶的合成处于被阻遏状态。在不含色氨酸的培养基中培养时,这些酶就显著增加,酶的合成处于消阻遏状态。曾经得到两种消阻遏突变型,不论培养基中是否含有色氨酸,它们的细胞中都含有大量和色氨酸合成有关的酶。一种突变型具有乳糖操纵子中的调节基因突变型i-的特性,它是调节基因突变型trpR-;另一种具有Oc突变型的特性,它是操纵基因突变型trpOc。从分枝酸到色氨酸这一代谢途径有5种酶进行催化作用,5个结构基因trpE,trpD,trpC,trpB和trpA按这一顺序紧密连锁,在trpE的前端便是操纵基因trpO。它们在一起构成色氨酸操纵子(图2)。作为一个功能单位,它和乳糖操纵子一样具有下列特性:
❶有关的酶按一定的比例合成;
❷通过转录形成一个多基因mRNA;
❸极性突变。
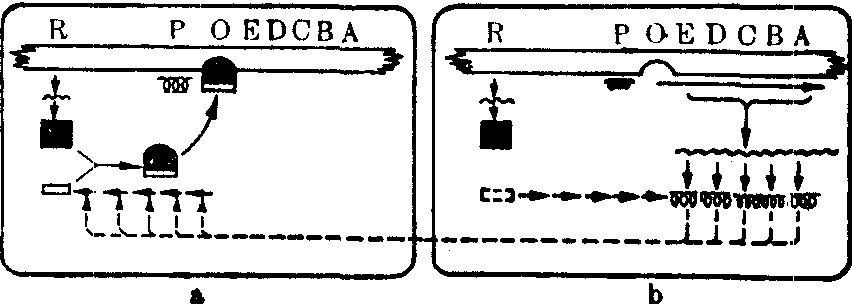
图2 色氨酸操纵子在色氨酸合成中的代谢调节功能示意图
a. 细胞中已经合成足够色氨酸时,色氨酸和调节基因产物——阻遏蛋白相结合,改变构型的阻遏蛋白和操纵基因相结合而使转录停止进行
b. 当色氨酸由于合成蛋白质而消耗时,阻遏蛋白不和操纵基因相结合,转录得以进行并通过转译合成相应的5种酶

正控制代谢调节 凡是某一细胞成分的存在使某种细胞功能能够实现,而这一成分的消失或失活使这一功能不能实现,这种控制称为正控制。在这里,酶的合成必需另一种调节基因的产物——降解物激活蛋白(CAP)的存在和活性,这种代谢调节功能称为正控制代谢调节。
在含有乳糖的培养基中加入葡萄糖时,大肠杆菌细胞中的β-半乳糖苷酶的含量便大为减少。cAMP能够抵消这种所谓葡萄糖效应。曾经发现两种在以乳糖作为唯一碳源的培养基上不能被诱导合成β-半乳糖苷酶的突变型。一种突变型在加入cAMP后便能被诱导,它是催化由ATP转化成为cAMP的腺苷酸环化酶的结构基因发生突变的结果。另一种突变型在含有cAMP的培养基上仍然不能被诱导,它是产生激活物CAP的调节基因发生突变的结果。这种降解物激活蛋白同cAMP结合后便被激活,被激活的降解物激活蛋白,促使转录进行。可见β-半乳糖苷酶的合成一方面需要消除阻遏蛋白的功能(通过乳糖等诱导物的作用),另一方面还需要CAP激活蛋白的作用。cAMP对于激活蛋白具有辅助作用,而葡萄糖的作用则是抑制cAMP的合成或促进cAMP的分解,因而它对于β-半乳糖苷酶诱导合成具有葡萄糖效应。所以乳糖发酵实际上受到双重控制,负控制和正控制。这一双重控制的适应意义是显然的。大肠杆菌在含有乳糖的培养基上才有合成β-半乳糖苷酶的需要,可是在同时含有乳糖和葡萄糖的培养基上。β-半乳糖苷酶的合成又不需要了,因为降解葡萄糖的酶都是组成酶,大肠杆菌可以优先利用葡萄糖而生活,直到葡萄糖消耗以后,才有必要合成β-半乳糖苷酶。
操纵子的多样性 调节子是由一个调节基因和若干操纵子所组成的代谢调节系统。大肠杆菌从谷氨酸到精氨酸的生物合成途径中,有8个结构基因参予作用,它们分处5个位置,每一个位置有一个操纵基因,构成5个操纵子。它们都受同一调节基因的控制而使这些酶的合成互相协调。虽然对于精氨酸合成的代谢调节系统目前有不同的解释,可是调节子仍然可以认为是一个有用的模型。自我调节是这样一种代谢调节系统,它的阻遏蛋白具有酶的作用,所以一个结构基因的产物为它本身所控制。鼠伤寒沙门菌从磷酸核糖焦磷酸到组氨酸的合成过程中,有9个结构基因参与作用,它们和一个操纵基因在一起构成一个操纵子。这个操纵子中的第一个结构基因的产物磷酸核糖-ATP焦磷酸合成酶和组氨酰tRNA相结合,成为作用于操纵基因的阻遏物。在这里,调节基因也是结构基因。
弱化子是操纵基因和第一个结构基因之间的起着减弱转录作用的一小段DNA。在色氨酸操纵子中,超越弱化子的转录受色氨酸浓度的影响,所以弱化子具有代谢调节作用。
在乳糖操纵子中,阻遏蛋白和激活蛋白是两个不同的基因产物。在和阿拉伯糖的分解代谢有关的阿拉伯糖操纵子中,调节基因ara C的直接产物是作用于操纵基因的阻遏物,它和L-阿拉伯糖结合后成为作用于启动区的激活物。所以这里阻遏物和激活物是同一基因产物。
转译控制 代谢调节也可以在转译水平上进行。RNA噬菌体感染大肠杆菌后,一方面进行复制,另一方面以它本身作为模板合成蛋白质。RNA噬菌体MS2只编码3种蛋白质,即外壳蛋白、吸附蛋白和复制酶。每一个噬菌体颗粒中有180个外壳蛋白分子,但只有一个吸附蛋白分子,所以它必须具备合成蛋白质的调节功能,而且这种调节功能是转译控制。MS2的RNA是单链分子,可是由于单链内部的碱基配对而形成复杂的花朵状构型,其中只有外壳蛋白基因的核糖体结合位置是暴露的,而其他二个结合位置都为二级结构所封闭,因此外壳蛋白合成得最多。外壳蛋白对于复制酶基因的转译又具有阻遏蛋白那样的作用,它和复制酶基因的核糖体结合位置相结合而阻碍复制酶基因的转译,从而使复制酶的合成只限于噬菌体繁殖的前一半时间。吸附蛋白基因的转译只发生在RNA复制过程中,而且核糖体只和新生的一个RNA链相结合,新生链稍一延长,二级结构又把结合位置封闭,因此吸附蛋白的合成只限于极短时间。
在细菌中,属于同一操纵子的基因经转录产生一个多基因mRNA,所以各个结构基因的mRNA数应该是相等的,可是细胞中愈是接近于操纵基因的酶含量却愈高。这一方面可能由于核糖体中途脱落,而脱落的可能性随着距离的增加而增加,另一方面可能由于转录和转译同时进行,而mRNA的分解则受核糖体的保护。由于前一原因,愈是接近操纵基因的结构基因被转译的机会愈多;由于后一原因,它们愈是不易被分解,也就有更多被转译的机会。
各种生物或同一种生物的各种细胞的mRNA的寿命常不相同。凡是影响mRNA寿命的因素都是一种转译控制因素。这些因素在高等动植物中研究得较多,而在微生物中研究得较少。
反馈抑制 无论是转录控制或是转译控制,控制的对象都是酶的合成。代谢调节的另一种方式是控制酶的活性。反馈抑制作用是合成代谢的最终产物对于合成途径中的第一个酶的可逆性的抑制作用。当合成代谢的最终产物大量存在时,它便和第一个酶结合,从而抑制了它的催化作用,其他反应也就停止进行。当最终产物的量减少时,酶的活性恢复,最终产物继续合成。
在以上这许多代谢调节系统中,小分子物质,不论是诱导物、代谢最终产物、cAMP都通过和一些大分子物质相结合而显示他们的诱导作用、消阻遏作用或激活作用,这些大分子物质便是调节基因所产生的阻遏蛋白或降解物激活蛋白基因所产生的激活蛋白。在反馈抑制这一种代谢调节作用中,代谢最终产物直接和酶结合而显示它的抑制作用,所以不存在调节基因,同反馈抑制直接有关的基因便是编码第一个酶的结构基因。
变构蛋白和区域化 以上所讨论的几种代谢调节功能有一个共同的特性,那便是都有一种变构蛋白参与作用。变构蛋白是一类能和特定结构的小分子物质结合而改变它的构型和活性的蛋白质。在糖发酵的诱导系统中,阻遏蛋白是一种变构蛋白,同诱导物相结合后的阻遏蛋白失去阻遏作用。在氨基酸合成的阻遏系统中,阻遏蛋白同代谢最终产物结合后出现阻遏作用。在乳糖发酵的正控制系统中,降解物激活蛋白和cAMP结合后出现激活作用。在阿拉伯糖发酵的正控制系统中,阻遏蛋白和阿拉伯糖结合后出现了激活作用。在反馈抑制系统中,第一个酶同代谢最终产物结合后失去催化作用。除了通过变构蛋白的作用而进行的代谢调节作用以外,在真核微生物中还有基于另一原理的代谢调节作用,那就是代谢作用的区域化。真核生物的细胞中存在着以膜为界的一些胞器(例如线粒体、叶绿体和胞核)。在这些胞器中,某些反应可以相对独立地进行,从而起到某种程度的代谢调节作用。
- 经济形式是什么意思
- 经济形式是什么意思
- 经济形式是什么意思
- 经济形式是什么意思
- 经济循环矩阵是什么意思
- 经济心理学是什么意思
- 经济心理学是什么意思
- 经济思想史是什么意思
- 经济性分析是什么意思
- 经济性双重征税是什么意思
- 经济性发行是什么意思
- 经济总量是什么意思
- 经济恐慌是什么意思
- 经济情报是什么意思
- 经济惩罚制是什么意思
- 经济成分是什么意思
- 经济成分(分fèn)是什么意思
- 经济成本是什么意思
- 经济成长阶段论是什么意思
- 经济成长阶段论是什么意思
- 经济成长阶段论是什么意思
- 经济成长阶段论是什么意思
- 经济成长阶段论是什么意思
- 经济战是什么意思
- 经济战是什么意思
- 经济战是什么意思
- 经济手册是什么意思
- 经济手段是什么意思
- 经济手段是什么意思
- 经济手段是什么意思
- 经济手段是什么意思
- 经济手段是什么意思
- 经济手段是什么意思
- 经济才是什么意思
- 经济扩张是什么意思
- 经济扩张是什么意思
- 经济批量是什么意思
- 经济批量是什么意思
- 经济批量法是什么意思
- 经济承包法是什么意思
- 经济技术协作交流洽谈会是什么意思
- 经济技术协作合同是什么意思
- 经济技术合作协定是什么意思
- 经济技术咨询服务是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区是什么意思
- 经济技术开发区投资优惠是什么意思
- 经济技术开发区法是什么意思
- 经济技术开发区的组织管理是什么意思