细胞质遗传cytoplasmic inheritance
染色体以外的遗传物质所决定的遗传现象。又称非孟德尔遗传或核外遗传。
1909年,重新发现“孟德尔定律”的法国植物学家科伦斯(C.E.Correns)报道紫茉莉 (Mirabilis ja-lapa)不符合孟德尔定律的遗传现象。花斑枝条(绿、白色镶嵌)♀×绿枝条♂的子一代有绿色、白色及花斑枝条;而绿枝条♀×花斑枝条♂的子一代全是绿枝条。类似的非孟德尔遗传现象在天竺葵 (Pelargonium Zo-nale)、月见草(Oenothera)、葫芦藓(Furnaria)及柳叶菜属(Epilobium)等植物中相继都有报道。研究者们提出细胞质基因组、质体基因及胞质基因等概念。1937年法国学者海蒂(P. H. Haitier)等报道果蝇对CO2敏感性的非孟德尔遗传。20世纪40年代相继在酿酒酵母、草履虫、大肠杆菌等微生物或原生生物中发现核外遗传现象。阐明这些遗传现象分别由细胞质中的线粒体、某些共生物及质粒控制。1963~1964年确证叶绿体及线粒体存在DNA,证实某些共生体及质粒亦具DNA性质,如果蝇的CO2敏感性因子σ和草履虫的卡巴粒,分别在细胞质中自成遗传体系。70年代绘制出质粒R100的遗传图。
正反杂交,是研究核遗传或细胞质遗传常用的方法。细胞质遗传时,正交及反交子一代总表现母本性状(如紫茉莉)、子二代表现不分离现象。子一代表现母本性状是因为双亲对子代提供相等的核物质,而细胞质只由母本提供。通过注射、接合转移(见细菌接合)等方法将某些生物(果蝇、大肠杆菌、草履虫等)的细胞质转移到遗传性不同的同类生物中,若受体获得供体的某些性状,该性状必属核外遗传。不断与父本回交,得到核置换材料(胞质来自母本,核来自父本)若仍表现母本某些性状,也是核外遗传的表现。显微操作进行核移植,从其性状表现亦可判断遗传方式。
生物有不少性状由细胞质基因或胞质基因与核基因共同作用决定。植物雄性不育特性在农作物杂种优势利用中的广泛应用就是例子(见雄性不育)。利用不同物种间叶绿体的摄取、转移可作为提高低光效作物(水稻、小麦)光合效率的手段。70年代报道烟草白花细胞原生质体摄取外源叶绿体后,能再生成正常烟草植株。矮牵牛及胡萝卜中亦有类似结果。
细胞质遗传cytoplasmic inheritance
又称染色体外遗传,核外遗传,非孟德尔式遗传或母体遗传。由细胞质基因决定性状表现的遗传现象。德国遗传学家柯伦斯(C.Correns)于1909年最先发现紫茉莉(Mirabilis jalapa)花斑植株的非孟德尔式遗传现象。现已证明该性状受质体基因控制。以后在粗糙面包霉、啤酒酵母、果蝇、草履虫以及许多植物中都发现这一现象。特点是:
❶正反交结果不同,F1均表现母本性状;
❷除花斑叶外,细胞质基因控制的性状一般不表现分离现象;
❸由附加体和共生体决定的性状,往往类似病毒的转导或感染;
❹细胞质基因可以通过连续多次回交,也可通过注射、细菌或草履虫的接合等方式转移;
❺细胞质基因对性状的作用有其自主遗传的特性,但某些性状受胞质基因和核基因的互作决定。应用重组分析、缺失分析、分子杂交和物理图谱分析等方法可以对细胞质基因进行基因定位。细胞质遗传已成为分子遗传学研究的一个重要项目。高等植物细胞质雄性不育基因的发现和研究,在杂种优势利用上有十分重要的价值。
细胞质遗传xibaozhiyichran
由细胞质内的遗传物质所控制的遗传现象称细胞质遗传,又称核外遗传、非孟德尔式遗传或母体遗传。细胞质遗传的特点有:(1)正反交结果不一致,杂种只表现母本性状,这是最早在高等植物中发现的判断染色体外遗传的依据。例如,1909年德国植物学家科伦斯发现在紫茉莉的杂交试验中,黄绿色叶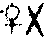 绿色叶的杂交子代都是黄绿色叶,而绿色叶♀×黄绿色叶的杂交子代都是绿色叶。这是由于真核生物的卵细胞除含细胞核外,还有大量的细胞质,而精子则除含细胞核外,仅有极少量的细胞质,所以合子的细胞质基本上来自卵细胞,当然合子的细胞质基因也基本上来自母本,那么,由这些细胞质基因控制的性状必然象母本,从而表现出母系遗传的特点。(2)细胞质遗传不遵循孟德尔的遗传规律,这是因为细胞分裂时,细胞质基因不是均等分配的,杂交后代一般不出现一定比例的分离现象 。(3)通过多次和雄亲本回交。
绿色叶的杂交子代都是黄绿色叶,而绿色叶♀×黄绿色叶的杂交子代都是绿色叶。这是由于真核生物的卵细胞除含细胞核外,还有大量的细胞质,而精子则除含细胞核外,仅有极少量的细胞质,所以合子的细胞质基本上来自卵细胞,当然合子的细胞质基因也基本上来自母本,那么,由这些细胞质基因控制的性状必然象母本,从而表现出母系遗传的特点。(2)细胞质遗传不遵循孟德尔的遗传规律,这是因为细胞分裂时,细胞质基因不是均等分配的,杂交后代一般不出现一定比例的分离现象 。(3)通过多次和雄亲本回交。
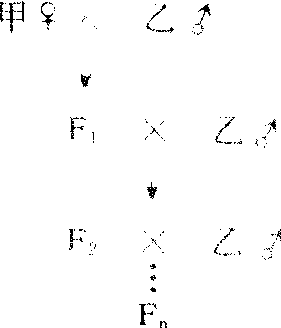
细胞质遗传cytoplasmic inheritance
系指以细胞质基因为物质基础发生的遗传现象而言。细胞质中的线粒体、中心体、叶绿体、动体等结构中亦含有DNA成分,亦能进行复制,能发生遗传并控制一定的遗传性状。高等生物细胞质多来自母体的卵细胞,因此由细胞质遗传产生的性状一般随母系遗传。
细胞质遗传
即“染色体外遗传”。
细胞质遗传cytoplasmic inheritance
亦称核外遗传。指受细胞质中基因控制的遗传性状。
细胞质遗传
细胞质遗传是指位于细胞质中的基因所控制的遗传现象,也称核外遗传或染色体外遗传。这些基因与核基因的成分相同,都是以DNA为遗传信息的载体,能自我复制和产生突变。细胞质遗传的共同特点是:
❶性状的遗传方式不符合孟德尔遗传规律。
❷子代只表现母体性状。
早在1909年有人已注意到紫茉莉(Mirabilis)叶的性状不符合孟德尔定律。以后又相继发现生物界中有许多类似现象,其中研究较透彻的有草履虫的放毒性,果蝇对CO2的敏感性,酵母小菌落变异和多种植物的雄性不育等。但直至本世纪60年代发现叶绿体和线粒体都含有DNA成分之后,细胞质遗传的概念才得到确认。现在一般将细胞质遗传分为三种类型:
❶细胞器的遗传:如线粒体和叶绿体。
❷细胞质中某些颗粒的遗传(内共生体):如CO2敏感型果蝇细胞中的Sigma颗粒和放毒型草履虫细胞中的Kappa颗粒等。
❸原核细胞中质粒的遗传。
判断细胞质遗传的主要实验方法有:
❶正反交:对某一性状具不同表型的个体之间进行杂交,如果不论正、反交(甲♀×乙♂或乙♀×甲♂)该性状在子代中都与母体相似,那么可以认为这种性状是由细胞质遗传的。因为细胞质的含量在雌雄配子之间相差悬殊,合子的细胞质几乎全部来自卵细胞,因此子代个体表现出母体性状遗传。真菌中有关线粒体的遗传,高等植物中有关叶绿体的遗传均有典型范例。近代应用分子生物学技术,鉴定了不同品系实验大鼠的线粒体DNA(简写mt DNA),发现它们的核苷酸顺序稍有差异(A型和B型)。A型mtDNA大鼠与B型mtDNA大鼠交配,子鼠的mtDNA与母鼠相同。
❷细胞质成分的转移: 某一生物的细胞质可通过不同方法转移给另一生物,如果后者出现前者 (供体)的某些性状,那么可以判断这些性状是由于获得了外源细胞质而产生的,属于细胞质遗传范畴。例如大肠杆菌的致育因子(F)和抗药因子(R)都是染色体以外能够决定性状的因子,它们可通过细菌的接合生殖,转移到F-细胞和R-细胞,使后者成为“雄性”和抗药性细菌。此外还可用微注射法将一种细胞的细胞质转移给另一种不同的细胞。例如果蝇对CO2的敏感性和粗糙链孢霉的小菌落性状都可通过微注射转移细胞质而获得。
❸核移植:用显微操作技术取出某种生物的细胞核,植入另一不同的去核细胞中,如果后者未因引入新的细胞核而改变其某些原有性状,说明这些性状是由细胞质基因控制的。从变形虫的链霉素抗性实验,从HeLa细胞和小鼠细胞的氯霉素抗性实验均已证实其属于细胞质遗传范围。
❹诱变:在不影响核基因的条件下,人工诱发变异可判断某些细胞质遗传性。例如吖啶橙等染料能够引起某些抗药性细菌变为敏感型,有致育因子的F+细菌变为F-细菌以及诱发酵母菌的小菌落变异等。
线粒体的遗传 线粒体是除蓝绿藻、细菌和成熟红细胞外普遍存在于动、植物细胞的细胞器,是细胞呼吸代谢和生成ATP的主要场所。线粒体由双层膜构成,外膜包围整个线粒体,内膜向内褶,延伸成嵴(见“线粒体”条)。内膜上有电子传递系统的四种复合物和ATP酶。线粒体有自己的DNA,其密度低于细胞核DNA,可用浮力梯度分开。变性实验表明线粒体DNA (mtDNA)与大肠杆菌DNA相似,为双链的环状分子。一条为重链(H链),一条为轻链(L链)。动物细胞的mtDNA周长约为5μm,分子量约为9×106—10×106,相当于15 000对核苷酸。原生动物和植物细胞的mtDNA大于前者,其周长分别约为25μm和50μm。在线粒体中未发现有组蛋白,对mtDNA的包装情况尚不了解。mtDNA在DNA聚合酶的作用下进行半保留复制,其方式与大肠杆菌类似,复制时间贯穿于整个细胞分裂间期。细胞器和mtDNA的相对含量及其在细胞中占细胞DNA总量的比例各不相同,如表1。这些DNA分子在线粒体基质中呈簇状分布。
表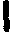 细胞器和mtDNA的相对含量及其在细胞中占细胞DNA总量的比例
细胞器和mtDNA的相对含量及其在细胞中占细胞DNA总量的比例
| 细胞种类 | mtDNA分子 /细胞器 | 细胞器/ 细胞 | 占细胞总 DNA的% |
| 大鼠肝细胞 小鼠L-细胞系 蛙卵 酵母二倍体细胞 | 5—10 5—10 5—10 2—50 | 11000 100 107 2—50 | 1 <1 99 15 |
根据对酵母菌、链孢菌、衣藻、有爪蟾蜍、小鼠和人等不同物种线粒体基因活性的研究,已知由mtDNA编码线粒体中的rRNA、tRNA和少量蛋白质基因,在线粒体核糖体上合成蛋白质。但另外还有许多蛋白质由核基因组编码,在细胞质核糖体上合成,然后输入线粒体与线粒体蛋白质共同建造结构,执行功能。但两者的遗传系统是分开的。线粒体有独立的蛋白质合成体系。实验证明:从细胞中分离出来的线粒体能在短期内继续进行DNA复制、转录和蛋白质合成;在整体细胞实验中加入蛋白质合成抑制剂,如环己胺(cycloheximide)抑制细胞质中蛋白质的合成,但对线粒体核糖体无影响,如果加入抗生素(氯霉素、四环素或红霉素)则抑制线粒体的蛋白质合成,对细胞质的蛋白质合成无影响。
线粒体基因组 酵母菌线粒体基因组包括2种rRNA基因,约30种tRNA基因和8种mRNA基因,其中7种为线粒体内膜蛋白质,1种为线粒体核糖体蛋白质。1981年Anderson等发表了人类线粒体DNA16569个核苷酸对的全部序列。其基因组含2个rRNA基因,22个tRNA基因和13个未鉴定的读码(URF)。在13个URF中有5个为有功能的蛋白质:3个细胞色素氧化酶亚基,1个ATP酶,1个为细胞色素b。1985年有人报告在其余的8个URF中6个是呼吸键NADH脱氢酶。人类mtDNA几乎全部用于rRNA和蛋白质的基因编码,与细菌基因组相似,未发现有非编码的间隔顺序(即内含子)。这一点与酵母菌不同,酵母菌的mtDNA中有大片段的内含子。牛和小鼠线粒体的基因组顺序与人类相似(图1)
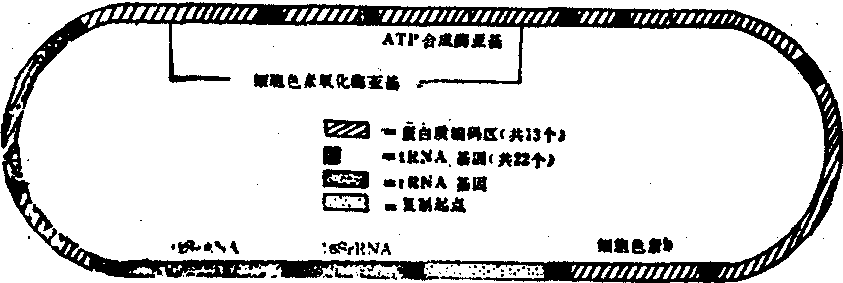
图1 人线粒体基因组
线粒体基因转录 人和小鼠的线粒体基因组中只有两个启动因子,一个位于H链,一个位于L链,其位置接近复制起点。从该处开始以两条链为模板分别沿双链的全长转录,合成两条不间断的RNA单链。这种转录方式为对称转录。核基因转录与此不同,主要以一条链为模板,而不对称转录。线粒体基因产物大部来自H链,转录后的RNA经核酸酶切割加工,产生2个rRNA,多数tRNA和约10个具有多聚腺苷酸(polyA) 的RNA分子。L链转录产物加工后产生8个tRNA和1个含少量polyA的RNA分子。其余90%为无用信息被降解。含polyA的RNA推测是线粒体的mRNA,其5′端缺少加帽。3′端带1个约55个核苷酸的polyA尾巴,是转录后由poly A多聚酶加添的。酵母菌线粒体基因组有大量内含子,有分开的启动因子。几乎全部基因从一条链转录,无对称转录证据。
线粒体mRNA 的翻译 翻译过程与大肠杆菌相似。蛋白质合成从N-甲酰蛋氨酸开始。线粒体mRNA因缺乏5′端和3′端的非编码区,相邻基因之间无间隔,核糖体如何与之结合而开始翻译尚不了解。mRNA的3′端更简略,翻译至任一终止符号立即停止。线粒体的一个终止符号为UAA,但许多mRNA中UAA是在转录后加添的。据模板核苷酸顺序分析,原始转录物切割后mRNA的3′端有U或UA,加添polyA后形成终止密码UAA,可见线粒体基因组编码之简练。线粒体核糖体与细胞质核糖体相似,含大小两个亚基,各亚基由蛋白质和rRNA组成。线粒体核糖体沉降系数为55S,比细胞质核糖体低。两亚基沉降系数分别为39S和28S,前者的rRNA分子为16S,后者为12S。线粒体中无细胞质核糖体的5S和5.8SRNA分子。线粒体tRNA与细胞质tRNA形态相似,结构和功能上存在重要区别。线粒体tRNA含59—75个核苷酸,有少数修饰碱基。比细胞质tRNA更有易变性。线粒体的某些氨酰tRNA与细胞质中的读码不同,线粒体的遗传密码与“统一遗传密码”表的差别见表2。
表2两种线粒体遗传密码与“统一遗传密码”比较

此外AUA和AUU也与AUG同样可用作起始密码。N-甲酰蛋氨酸是所有多肽链合成的第一个氨基酸。
叶绿体的遗传 叶绿体是绿色植物进行光合作用的细胞器,其结构为双层膜包裹的片层系统,内部包括许多由小片层构成的基粒,基粒之间为基质(见“叶绿体”条)。1970年有人用渗透震荡法(Osmotic shock) 研究伞藻(Acctabularia)的叶绿体,发现最完整的DNA片段长约419μm,约含1.23×106碱基对。一般高等植物的叶绿体DNA(简写cpDNA)为环形的双螺旋结构,分子量约85×106—97×106,相应为1.3×105—1.5×105碱基对,在叶绿体基质中也呈簇状分布。cpDNA的复制与叶绿体的分裂贯穿于整个细胞分裂间期。叶绿体与线粒体相似有全套的遗传体系,有自己的RNA和核糖体。其核糖体也对某些抗生素敏感,对细胞质的蛋白质合成的抑制剂无明显效应。叶绿体本身具有特定的表型性状,如绿色和光合作用的能力等。
cpDNA比mtDNA复杂,与细菌的遗传系统更相近,cpDNA和叶绿体mRNA能在细菌的转录一翻译系统中精确地进行转录和翻译,甚至可在杂交核糖体上进行蛋白质合成。所谓杂交核糖体是指叶绿体的核糖体小亚基与细菌核糖体大亚基的结合物。叶绿体核糖体的亚基与同一有机体的细胞质核糖体亚基则不能结合,说明叶绿体核糖体与细胞质核糖体的遗传体系不同。叶绿体合成蛋白质也与线粒体和细菌相似从甲酰基蛋氨酸开始,合成后在酶的作用下切除。
在高等植物中,花粉中的叶绿体不进入合子并保持严格的母体遗传者,大约占2/3。其余植物有的花粉叶绿体也进入合子。这样,细胞质遗传的性状可来自双亲。例如植物的杂色叶,即合子中有正常的叶绿体和缺陷型的叶绿体。通过生长过程中的细胞有丝分裂,叶绿体逐渐分离,产生两种含不同叶绿体的细胞,使叶面上出现绿色和白色斑块。绿色斑块的细胞中含正常叶绿体,白色斑块中含缺陷型叶绿体,说明叶片颜色决定于细胞质而不是决定于细胞核。
细胞质遗传与核遗传的关系 细胞中的线粒体和叶绿体两者都是有机体的能量细胞器。构成这两种细胞器的蛋白质除去它们自身合成的以外,都有大量蛋白质来自细胞质。例如线粒体含蛋白质约数百种,远远超出线粒体基因组的编码限度。事实上线粒体本身仅合成少数几种蛋白质,其余均由细胞核基因组编码,在细胞质中合成。线粒体中的多聚酶、合成酶、可溶性因子和线粒体蛋白质等就是依赖一种需能的运输过程进入线粒体的。一旦进入便立即与线粒体中的蛋白质共同组装各种内膜复合物。这些由核基因组编码的蛋白质是建造细胞器不可缺少的组成部分(图2)。它们在两种不同的遗传体系中如何沟通与协调,现在尚未充分证明。但在整体细胞中细胞器的蛋白质合成主要受细胞核的控制,至少在线粒体中如此。当线粒体的蛋白质合成被阻断时,则引起细胞过量生产各种涉及mtDNA、RNA和蛋白质合成中输入的各种酶。似乎细胞接受了来自细胞器的反馈信息,以克服阻断作用对蛋白质合成的影响。
从进化角度看,这些细胞器可能起源于古老的真核生物与细菌或藻类所建立的稳定共生体系,以后紫细菌发展为线粒体,蓝绿藻发展为叶绿体。虽然它们还具有独立的遗传系统,但其中大多数蛋白质基因已整合到细胞核基因组中(见“线粒体和叶绿体的内共生起源学说”条)。
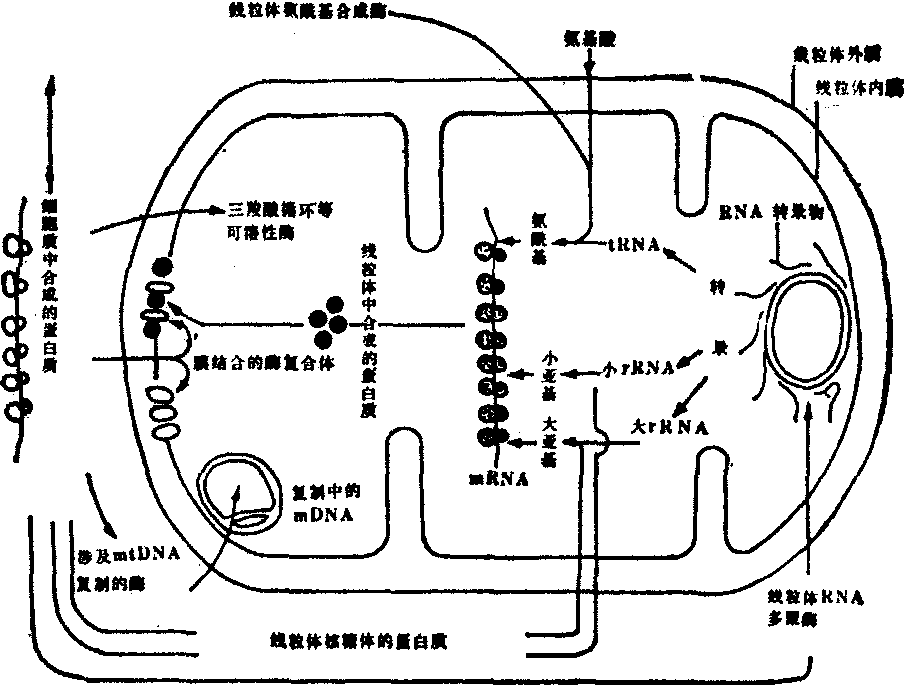
图2 线粒体中输入蛋白质的作用示意图
细胞质颗粒的遗传
CO2敏感型果蝇sigma颗粒的遗传 草履虫中有的品系对CO2高度敏感,在低浓度CO2环境中短暂停留竟麻醉致死。同样条件处理野生型果蝇。回到空气中不久即可恢复正常。这种性状是由细胞质中的Sigma颗粒引起的。Sigma颗粒形似致病病毒(VSV),在正常环境中对果蝇无明显致病效应。电镜观察Sigma颗粒呈枪弹形,长140—180nm,直径为70nm。含DNA成分,具突变性。在细胞内不依赖细胞核而独立增殖。主要通过卵细胞向后代传递,偶尔也可来自精子。人工注射敏感型无细胞提取物,可使野生型果蝇变为敏感型。Sigma颗粒做为细胞质成分存在,但显然为感染的外源性颗粒。其引起宿主细胞对CO2敏感的机制目前尚不了解。
放毒型草履虫Kappa颗粒的遗传 某些草履虫可产生一种草履虫素,对其他个体有杀伤作用,因而称放毒型草履虫。这种毒素由细胞质中的Kappa颗粒产生。其形态似小型细菌,长约1—5μm,直径约0.2μm,成分含DNA、RNA以及一些酶系和细胞色素。每一放毒型草履虫含Kappa颗粒的数目不等,可高达1600个。草履虫结合生殖时Kappa颗粒可通过细胞质桥进入非放毒型个体。由细胞质遗传给下代。Kappa颗粒的存在依赖核基因κ,基因型κκ的个体,虽有Kappa颗粒,经几次分裂后也自行消失。Kappa颗粒的存在与否,并不影响草履虫本身的正常生存和生长。可以认为这种颗粒也是一种由微生物感染而建立的内共生体。
质粒的遗传 质粒是存在于细菌和酵母菌细胞中的小型环状双链DNA分子,是除染色体以外的独立复制单位。质粒DNA载有与复制和转移有关的基因,还编码使宿主细胞具有某些性状的基因。例如大肠杆菌的致育性、抗药性和产生大肠杆菌素的基因等。有些质粒在细胞中游离存在,有些可与宿主细胞染色体整合 过去认为质粒仅代表前者,而附加体代表既能整合又能游离存在的分子,现在两者常作同义语混用,为了简便不管能否整合统称质粒。质粒的存在或消失与宿主细胞染色体无关,在正常条件下不影响细菌的生长和繁殖。质粒的数目与复制特点有关,根据质粒在细胞中的拷贝数可分为两种类型:一种是拷贝数少,每个细胞中只含1—4个,这种质粒称为严紧型质粒。严紧型质粒的分子量较高。另一种质粒分子量较低,拷贝数多,在染色体的复制停止后,仍能继续复制,每个细胞中约含10—20个,这种类型称为松弛型质粒。像这些分子量低、拷贝数多、能够扩增、有被某些限制性内切酶切割的专一切点、带有某些易于选择的遗传标记的质粒DNA,常用作基因工程中的运载体。
细菌质粒 主要有以下三种。
(1) F因子: F因子是发现较早的一种质粒,具有这种质粒的细菌可以进行“有性”生殖。F+细菌为“雄性”,F-细胞为“雌性”。“雄性”细菌的表面生长性伞毛,通过接合生殖F因子可以转移到“雌性”细胞,使之变为“雄性”细胞。F因子在细胞中既可游离存在又可与染色体整合,成为高频重组体(HFR)。质粒DNA由103—105碱基对组成,有性伞毛和其他有关转移的基因编码。另外,还包括一些插入顺序(IS)。这些顺序与大肠杆菌染色体上相应的顺序联会和交换,使F因子与细菌染色体整合形成HFR。
(2) R因子:是携带抗一种或多种药物基因的质粒,包括抗氯霉素、新霉素、青霉素、链霉素、磺胺和四环素等。这种抗药性质粒由抗性转移因子和抗性因子两部分组成,控制抗性的转移。后者编码不同的抗生素钝化酶使宿主细胞对抗生素产生抗性,但它们不能单独转移,必须是完整的R因子质粒才能转移。R因子的转移不仅发生在种间,也可发生在属间,例如大肠杆菌与痢疾志贺菌之间R因子的转移。R因子包含1.5×104—105碱基对。每细胞中含1—3个拷贝。
(3) Col因子: 是使某些肠道细菌能够产生大肠杆菌素的质粒。大肠杆菌素是一种有高度特异性的蛋白质抗体。对同种的其他菌株或有关菌种有杀伤作用。因这些细菌细胞壁表面有特异性受体部位,故为敏感型;反之无特异性受体者具抗性。大肠杆菌素的种类很多,有各自的杀伤作用。其中大肠杆菌素K的杀伤作用主要是抑制DNA复制、转录和多肽的合成。有些Col因子质粒通过接合生殖转移,也有些不具转移功能。Col因子DNA大约含碱基对6 000—21 000之间。现在基因工程中广泛利用ColEJ质粒为基因载体。
细胞质遗传
cytoplasm heredity
- 来盟坎是什么意思
- 来知德是什么意思
- 来知德是什么意思
- 来祖荫是什么意思
- 来祖荫是什么意思
- 来福灵是什么意思
- 来福阿德染料水是什么意思
- 来禽馆帖是什么意思
- 来禽馆集是什么意思
- 来秀是什么意思
- 来米多舒是什么意思
- 来者不善是什么意思
- 来者不善,善者不来是什么意思
- 来者不善,善者不来是什么意思
- 来者不拒是什么意思
- 来者勿拒是什么意思
- 来者可追是什么意思
- 来而不往非礼也是什么意思
- 来而不往非礼也是什么意思
- 来自是什么意思
- 来自地狱的报告是什么意思
- 来自大洋彼岸的报告是什么意思
- 来自大自然的寓言是什么意思
- 来自太空的挑战是什么意思
- 来自异国的孩子是什么意思
- 来自异国的孩子是什么意思
- 来自异国的孩子是什么意思
- 来自神秘世界的报告是什么意思
- 来自竞争的繁荣是什么意思
- 来自远方的信息——通讯史话是什么意思
- 来自青年的报告是什么意思
- 来自非洲是什么意思
- 来自音像世界的警示是什么意思
- 来航鸡是什么意思
- 来苏丹是什么意思
- 来苏丹是什么意思
- 来苏丹是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏散是什么意思
- 来苏汤是什么意思
- 来苏汤是什么意思
- 来苏膏是什么意思
- 来苯他明是什么意思
- 来裕恂是什么意思
- 来访和重访登记表是什么意思
- 来访者中心是什么意思
- 来访者中心疗法:其实践、含义与理论是什么意思
- 来访问题是什么意思
- 来踪去迹是什么意思
- 来辛马卡斯是什么意思
- 来辟是什么意思
- 来阿入赤是什么意思
- 来鹤亭诗是什么意思