痛觉tòng jué
身体组织因受破坏或受强烈的刺激所产生的感觉。1909年周树人《人生象学·本论》:“痛觉: 凡知觉神经所在地,一受强撄,莫不感此。”1919年汤尔和译《诊断学》下卷:“第三原基终于大脑后正中回转之皮质中枢,此道以传搬温觉、痛觉及触觉为主。”
疼痛
疼(生~;刺~) 痛(痛楚;楚痛;疾痛;肉~;骨~;跳~;发~;负~)
感觉疼痛:痛觉
痛觉和痒觉:痛痒(~相关)
难受或疼痛的感觉:火辣
感到痛苦或疼痛的地方:痛处
酸麻疼痛:辛(辛楚)
酸疼发麻:酸麻
酸疼痛楚:酸楚
又酸又痛:痠痛 酸痛
痛时且觉酸软:痠痛 酸痛
酸疼无力:酸软
人身肌肉过度疲劳或因病引起的酸痛无力的感觉:痠
酸痛之极:酸削
因受伤而疼痛:伤痛
头部疼痛:头疼(~脑热)
因思念而引起的头痛:首疾
头脑眩晕胀痛:头昏脑胀 头昏脑涨 头昏脑闷 昏头胀脑 昏头涨脑
心脏所在部位感觉疼痛:心痛
刺激皮肉而感到疼痛:刺痛
肌肤因受淹渍刺激而疼痛:刺人
高温与皮肤接触感觉疼痛:烫(~手)
因被火烧或鞭打等而产生疼痛的感觉:火辣辣 辣豁豁
烧灼一样的疼痛:灼痛
燥热灼痛:火焦火辣
轻微疼痛的感觉:酸溜溜
隐隐约约的疼痛:隐病 隐伤
感到疼痛:作痛
剩余的疼痛:遗痛 遗恫
大痛之后未全部消失的痛楚:余痛
非常疼痛:疼得要命 痛不可言 痛不可忍 火烧火燎
疼痛之甚:痛煞煞
彻心疼痛:十指连心
疼痛到极点:彻心彻骨
极度疼痛:死去活来
发疼,很疼:生疼
剧烈的疼痛:剧痛
由于某些病症引起的内脏剧烈阵发性疼痛:绞痛
隐隐微痛的样子:阴阴
胀痛的样子:岑岑
疼痛难忍的样子:龇牙咧嘴 龇牙裂嘴 呲牙咧嘴 傞牙俫嘴 露嘴龇牙 雌牙咧嘴 雌牙露嘴 雌牙扭嘴
酸痛剧烈的样子:酸嘶
(疾病、创伤引起的 难受的感觉:疼痛)
另见:痛苦 难受
感觉到
着(~慌) 悟 觉得 觉道 感到
感知,意识到:觉(听~;嗅~;味~;温~;冷~;触~;肤~;色~;痛~;幻~)
以此感到骄傲和自豪:引以自豪
容易感觉到:显然
深切地感觉到:痛感
深深感到:深感
(客观事物的个别特征在头脑中引起的直接反应:感觉)
痛觉pain sense
伤害性刺激作用于痛感受器所引起的一种感觉。常伴有情绪活动和防卫反应。对机体具有保护意义。痛感受器一般认为是游离神经末梢。任何刺激只要达到一定的强度成为伤害性刺激时,都可引起痛觉,故痛觉不是由单纯一种刺激引起的感觉,且伴有痛反应。
痛觉tongjue
伤害性刺激作用于皮肤及身体其他部位时所引起的一种强烈感觉。痛觉感受器一般是游离的神经末梢,分布于皮肤及全身各处。引起皮肤痛觉的刺激主要是过强的机械力、过冷、过热及强的物理、化学因素等。皮肤内感受器呈点状分布,当损伤性刺激作用于皮肤时,可先后出现两种性质不同的痛觉,即快痛和慢痛。快痛尖锐,定位清楚,刺激除去即消失。慢痛为 “烧灼痛”,定位不明,过后才能感觉到。内脏痛可由机械牵拉、痉挛、缺血和炎症等引起,由交感神经传入。痛觉一般缓慢、持续、定位不精确。痛觉常伴有不愉快的情绪活动和防卫反应。
痛觉tongjue
对于引起身体疼痛的刺激物的感觉。包括皮肤痛觉、深部痛觉和内脏痛觉等。痛觉感受器游离于神经末梢,传导纤维有两种:AS纤维和C纤维,兴奋传至大脑皮层引起痛觉。一些研究认为,痛觉可以适应,而且在一定条件下可出现完全的适应。痛觉感受性因身体部位不同而异,背部与颊部最敏感,四肢前端最不敏感。痛觉感受性还因性别、经验、耐受力等的不同而不同。此外,一种痛觉能抑制另一种痛觉。痛觉是有机体内部的警报系统,对避开危险、保护有机体的生存有重要意义。无痛觉的人就等于缺少了一种天然保护屏障。
痛觉tongjue
身体组织因受破坏或强烈刺激所产生的感觉(见图)。人体的各组织器官中,都分布有一些特殊的游离神经末梢,若受过强的机械力、过冷、过热以及其它各种强的物理或化学因素作用时,都能引起痛觉。这种特殊游离神经末梢在皮肤中分布的部位称为痛点。据估计,每1平方厘米皮肤表面约有100个痛点。在皮肤表面痛点数达100万个。近年来,科学家发现,当组织损伤或者发炎时,组织细胞或者血细胞中都会渗出一些物质,如组织胺、5-羟色胺等。这些物体有引起痛疼的作用,称为“致痛物质”。
痛觉
痛觉algesthesia
系一种浅感觉。由周围神经支配,分布于全身所有器官与组织,其中以皮肤、粘膜为最敏感。检查时用大头针等尖具刺扎皮肤,要对称、同等强度对比进行,以了解敏感程度。
痛觉
辨别对机体具有伤害性的刺激的感觉。无专一的适宜刺激,机械的、物理的、化学的、温度的、放射能的以及电的各种刺激,达到对有机体起破坏作用时,都会引起皮下游离神经末梢的兴奋,在脑中产生痛觉。
痛觉
疼痛是人们普遍的经验,但至今尚无满意的科学定义。Aristotle认为疼痛是和愉快相对立的感觉。一般认为疼痛包括痛感觉和痛反应二方面,并伴有强烈的情绪成份,表现为一系列躯体和内脏反应,甚至个体主动逃避或防御等复杂动作。von Frey (1895)最早提出疼痛感受器是游离神经末梢。后来有人检查身体上对痛最敏感的部位,如鼓膜、角膜、牙髓,这些区域只有游离神经末梢存在,进一步支持游离神经末梢是疼痛感受器的想法。1970年林可胜提出,终止在血管旁的无髓游离神经末梢分支,可能就是疼痛的化学感受器。最近,Iggo,Burgess等发现皮肤中有两种特异性的伤害感受器。一种是由Aδ纤维传导的机械-伤害感受器;另一种是由C纤维传导的对伤害性热(45℃以上)、化学、机械刺激都发生反应的多型伤害感受器。后者是一种比较原始的感受器,在皮肤、内脏、肌肉中都有,它们可能就是特异性的疼痛感受器。
疼痛的传入纤维有二类,一类是Aδ纤维,可能和传导快痛有关;另一类是C纤维,可能和传导慢痛或定位不清的烧灼痛有关。 1973年以来,Torebj rk和Hallin等用钨丝微电极经皮插入神经干的方法,记录了清醒人外周神经中单个Aδ纤维和C纤维的活动,进一步证明Aδ纤维的活动和刺痛有关,而C纤维的放电则引起持续的慢性痛和烧灼痛。
rk和Hallin等用钨丝微电极经皮插入神经干的方法,记录了清醒人外周神经中单个Aδ纤维和C纤维的活动,进一步证明Aδ纤维的活动和刺痛有关,而C纤维的放电则引起持续的慢性痛和烧灼痛。
关于疼痛的传导束,经典的观点认为是通过前外侧索上行的。前外侧索或称脊丘系统,包括三个部分,即新脊丘束、旧脊丘束和古脊丘束,或称脊网束。有人认为快痛主要由旧脊丘束和一部分新脊丘束传导,慢痛主要由旧脊丘束和脊网束传导。Melzaek等将旧脊丘束、脊网束和脊髓中脑系统等合称傍中央上行系统,它们突触较多,主要传导弥散性痛。除了上述脊丘系统的传导束外,现在有人认为可能背索、脊颈束和脊髓固有系统等其他传导束在痛觉传导上亦起一定作用(图)。
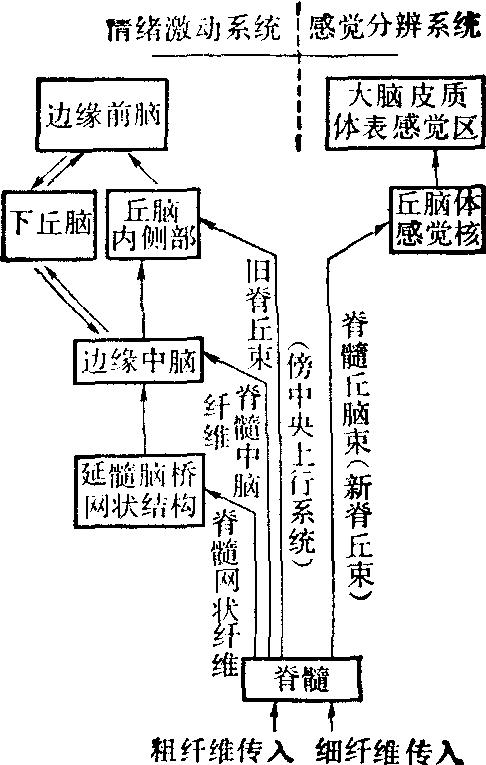
脊髓痛觉传导束示意图与痛觉有关的各级中枢 有脊髓、脑干、丘脑、大脑皮质、边缘系统和基底节。
脊髓 Rexed根据脊髓背角灰质细胞的大小、方向、分布等特点将脊髓背角分为六层。和疼痛有关的神经元主要分布于第Ⅰ层、第Ⅳ层和第Ⅴ层。Mayer认为Ⅳ和Ⅴ层中的广动力学细胞在灵长类可以产生痛觉,但第Ⅰ层细胞在痛觉中的作用尚不太肯定。
脑干 经脊髓前外侧索上行的纤维大部终止在脑干网状结构中。延髓网状结构中的巨细胞核和痛觉有密切关系。在不麻醉的猫,延脑网状结构内侧部大多数神经元对痛刺激起反应。在动脉注射缓激肽后,引起动物强烈疼痛反应时,许多网状巨细胞核中的神经元就发生强烈兴奋。此外从延髓内侧网状结构中有二条和痛觉有关的下行性抑制系统: 一是缝际脊髓系统,起自脑干缝际核,主要是缝大核,为5-羟色胺能神经元组成,主要分布于同侧背角第Ⅰ层、第Ⅱ层等。另一是背侧网脊系统,它来自延髓腹内侧的缝际旁系统,所终止部位也与缝际脊髓系统相似,它的神经介质不明,可能是去甲肾上腺素能纤维。上述两条道路都通过背外侧索下行。中脑中央灰质是痛觉通路中的一个重要环节。因为脊丘束和脑干网状结构有许多纤维投射于此。电刺激中脑中央灰质在猫和鼠都有镇痛作用。这可能是激活了脑干中的下行抑制系统所致。
丘脑 经前外侧索上行的痛觉信号,也和其他感觉一样,首先集中到丘脑,其中和痛觉有关的核团主要有三群。(1) 丘脑腹侧基底复合体(VB):它包括腹后外侧核(VPL)和腹后内侧核(VPM)二部分。新脊丘束直接投射到VB的较尾部,有人认为它主要投射到VPL,并和痛觉的定位和辨认有关。
(2) 后核群(PO): PO区的细胞大约60%对痛刺激起反应。刺激停止后,有明显的后作用。PO区细胞感受野很大。其中有些细胞对电刺激牙髓亦起反应。静脉注射吗啡能抑制这种反应。由于旧脊丘束投射到PO,故它和痛觉的关系就特别引人注目。
(3) 内侧核群: 前外侧索上行的大量无髓纤维终止在内侧核群,主要是束傍核(Pf)、中央外侧核(CL)和中央中核(CM)等。有人认为束傍核和中央外侧核只对伤害性刺激起反应,而吗啡能抑制这种反应。Perl等发现CM中有对伤害刺激起反应的单位。刺激人Pf核及其附近可加剧病人痛觉症状,毁损此区可缓解疼痛。
大脑皮质 从丘脑VB主要投射到大脑皮质第Ⅰ体感区(SⅠ),部分投射到第Ⅱ体感区(SⅡ)。在猴的SⅠ细胞,对皮肤刺激发生反应的细胞中只有2%对伤害刺激起反应。刺激人的SⅠ也只有2.5%引起刺痛。有人报告切除SⅠ后引起痛觉缺失,相反亦有报告切除后出现痛觉过敏的。有人报道刺激猫、鼠和人的牙髓可在大脑皮质记录到诱发电位。
SⅡ和痛觉的关系比SⅠ更为密切。在SⅡ,对皮肤起反应的细胞中有38%对伤害刺激也起反应。在SⅡ尾部有些神经元有较大的二侧感受野,只对伤害刺激起反应。损伤猫的SⅡ会增加痛阈。SⅡ还接受丘脑后核群的投射。有人认为PO-SⅡ区径路对痛觉信号具有整合作用。
边缘系统和基底节 海马是边缘系统的一个重要部位,刺激猫海马背部时,能抑制内侧核群的细胞放电和提高痛阈。边缘系统中其他部位亦和痛觉有关,如直接刺激人的杏仁核可引起痛觉,而电刺激膈区、视前区均可导致痛阈增加,甚至缓解痛。近年来许多作者在边缘系统及其有关结构如扣带回、下丘脑等都记录到痛敏细胞,它们可能和痛的情绪成份有关。
此外,最近有许多实验证明电刺激动物或人尾核具有明显的镇痛作用。
电刺激脑和吗啡的镇痛效应 近几年来,发现电刺激脑干中某些部位能产生明显的镇痛作用。1969年Reynold报道局部电刺激大鼠中脑灰质背侧部时引起明显镇痛作用,其有效区域包括位于脑室周围灰质和导水管周围灰质附近。1962年邹岗等发现兔脑室内注射微量吗啡有强烈的镇痛作用。后来,一般认为注射微量吗啡镇痛作用最强的部位分布于下丘脑内侧部,底丘脑和中脑导水管周围灰质等处。
电刺激脑镇痛和脑内注射微量吗啡镇痛开始是从不同角度来研究痛觉机理的,但随着研究的深入,发现二者有很多相似之处:
❶二种镇痛的有效脑区相似,主要都分布于室周灰质和导水管周围灰质。
❷二种镇痛均与脑干缝际核群有关。特别和中缝背核和中缝大核有密切关系。
❸二者都与5-羟色胺递质系统的参与有关,这类纤维经背外侧索下行,抑制背角第Ⅴ层神经元对伤害刺激引起的放电,而达到镇痛作用。
❹二种镇痛作用在相当程度上都可被纳洛酮所对抗。
现在认为,电刺激脑的镇痛作用可能是通过释放内啡呔而实现的。而吗啡有拟内啡呔作用。二者都作用于导水管周围灰质,可能和吗啡受体相结合而兴奋中缝核群,特别是兴奋中缝大核等5-羟色胺能神经元,经下行抑制系统由背外侧索下行抑制背角第Ⅴ层细胞的活动而镇痛。由于钠洛酮只能部分对抗这种镇痛作用,因此脑内可能还有另一条下行抑制系统参与抑制痛觉。可能从延髓网状巨细胞核下行的去甲肾上腺素系统在电刺激镇痛和吗啡镇痛中亦起着相当作用。
吗啡受体与内啡呔 1973年Terenius等证明吗啡受体的存在。1975年,Haghes首先从脑中发现能和吗啡受体结合的二种活性物质,即甲-硫氨酸脑啡呔和亮氨酸脑啡呔,它们都是五肽。脑啡呔的意思是脑内具有吗啡样作用的肽,除了这些五肽外,后来还陆续发现一些分子量较大的吗啡样活性物质,它们主要从脑垂体中提出的。现在统称为内啡呔。Goldstein将内啡呔定义为“有吗啡样活性的内源性物质”,而脑啡呔仅指内啡呔中具有五肽的物质。到目前为止已发现有9种内啡呔:
❶甲-硫氨酸脑啡呔;
❷亮-氨酸脑啡呔;
❸α-内啡呔;
❹β-内啡呔;
❺γ-内啡呔;
❻垂体吗啡肽;
❼止痛素;
❽强啡呔;
❾脑新肽等。
一般认为脑啡呔可能是真正的神经递质,而分子大一些的内啡呔,特别是β内啡呔等垂体肽,因为它们亦分布在血液中,而且性质比较稳定,所以很象激素。
吗啡受体和脑啡呔的分布 吗啡受体在灰质内多,白质中少。在人及猴,杏仁核中分布浓度较高,其次为下丘脑、丘脑内侧、尾状核头部、中脑中央灰质;海马、嗅三角及大脑皮质的某些部分。此外吗啡受体还分布在脊髓罗氏胶样质区,而这些部位正是躯体和内脏C纤维传入的部位,表明有相当多的受体分布在初级传入纤维的末梢上。
脑啡呔在脑内分布很难正确判定,而且分布不均匀,但它和吗啡受体的分布有相当平行的关系。在猴脑的尾状核、苍白球、下丘脑前端等处脑啡呔浓度很高,其次是杏仁核、中脑导水管周围灰质以及第四脑室底等部。根据吗啡受体、脑啡呔的分布情况和痛觉传导通路,电刺激镇痛和吗啡微量注射镇痛的有效脑区的关系看来,吗啡受体和内啡呔系统在疼痛的控制上具有极为重要的作用。
内啡呔的镇痛作用 一般认为脑啡呔在整体动物中的镇痛作用很弱,给动物脑室内注射很大剂量才出现微弱的镇痛作用,而且只能持续几分钟。但Takagi报道将5μg的甲-硫脑啡呔微量注射到鼠的延髓巨细胞核 (NRGC)内能产生较强的镇痛作用,并可持续30分钟,应用钠洛酮2mg/kg可对抗上述镇痛作用。Takagi认为脑啡呔微量注入巨细胞核出现明显镇痛作用,表明巨细胞核是脑啡呔产生镇痛作用的重要作用点。内啡呔中,镇痛作用最强的是β-内啡呔,据Loh等(1976)报道,β内啡呔在大鼠中其镇痛作用可比吗啡强18~33倍,而甲硫脑啡呔在同一实验中却仅有很弱的镇痛作用。β内啡呔无论静脉注射或脑内注射,都能产生明显的镇痛作用。
疼痛学说 关于疼痛的学说,长期以来一直存在着剧烈的争论,至今没有完全解决,比较重要的痛觉学说有特异学说、型学说与闸门学说等。
特异学说 认为痛是由伤害刺激作用于特殊的感受器(游离神经末梢),经Aδ和C纤维,经脊丘系统,传导到丘脑内侧核群,而形成痛觉。这个学说是在Müller的感觉器官特殊神经能量定律的基础上提出的。后来vonFrey根据皮肤感觉的点状分布和各种感受器的形态分布特点而提出疼痛的特异学说 。当时他只涉及到痛觉感受器的特异性。后来其他学者进一步发展到寻找从感受器传向中枢的特殊传入纤维和疼痛的特殊传导束和有关的特异性痛中枢。特异学说历史较长,有较丰富的实验根据,并有较多的临床支持; 但亦有一些缺点,特别是1950年以来Weddel等认为人角膜和耳廓上只有游离神经末梢,却能感受触、冷、热、痛等各种不同性质的感觉。这些事实表明游离神经末梢并非是疼痛的特异性感受器,而是具有更为广泛功能的感受装置。此外,亦有一些临床资料不支持特异学说,如在外科作前外侧索切断术后,并不能持久地消除疼痛,常有复发现象。而且幻肢痛、烧灼痛、中枢性痛等临床现象,亦不支持疼痛是一种固定的、直线的、点对点的神经系统连系。
型学说 Goldscheide (1894)首先提出了刺激强度和中枢整合是疼痛的决定因素。他认为来自皮肤的感觉输入,在背角细胞中的综合可能会产生一种特殊的构型,它可能和疼痛有关。过强地刺激各种感受器,或在病理条件下由于加强对非伤害性刺激的综合作用,当细胞输出超过临界水平时,就会引起痛觉。Weddel (1955)和Sinclair认为,所有的皮肤感觉性质是由神经冲动在空间和时间上的构型所产生的,而不是由不同性质的感受器和特殊传导通路引起的。皮肤上各种感受器在功能上没有很大的差别。各种刺激引起不同感觉,只是因为神经纤维中传导的冲动在时间和空间上呈现不同的组合型式。过强刺激引起大量的传入冲动,在中枢就产生了痛。型学说能较多地解释临床现象,但提法过于笼统,实验事实较少,目前还未能形成一个严谨统一的理论基础。
闸门学说 闸门学说系Melzack和Wall (1965) 所提出,是吸收了特异学说和型学说的优点,综合而成的一种新的痛觉学说。它认为刺激皮肤产生的传入神经冲动,被传导到脊髓的三个系统中去:
❶背角中的胶样质细胞;
❷经背索纤维投射到脑;
❸背角中的第一级中枢传递细胞(T细胞)。
Melzack认为胶样质细胞 (SG)位于背角第Ⅱ、Ⅲ层中。它的作用类似闸门,能调节外周传入冲动与T细胞之间的突触传递。T细胞位于第Ⅴ层中,接受从皮肤、内脏、肌肉中的细纤维传来的冲动,其活动亦受到脑的下行性纤维的影响。它们对各式各样的刺激强度都起反应,刺激越强,放电频率就越高。
SG细胞是闸门的关键,它以突触前抑制的方式调节T细胞的放电水平。粗纤维的传入冲动兴奋SG,细纤维的传入冲动抑制SG。当粗纤维的传入冲动兴奋T细胞时,同时亦兴奋SG。SG的兴奋可使粗、细传入纤维的末梢都去极化(突触前抑制),表现为负的背根电位。从而抑制了T细胞的放电。相反,当细纤维传入冲动时,则抑制SG,从而解除了对粗、细纤维的突触前抑制,而出现容易化(表现为正的背根电位),这就导致T细胞持续放电。如果细纤维接连传入冲动,使SG持续受到抑制,T细胞的放电会一再加强,当T细胞的输出超过临界水平,就会激活作用系统而引起痛感觉和痛反应。综上所述,粗纤维的传入活动通过突触前抑制而关闭闸门,有止痛作用。细纤维的传入活动通过突触前容易化而开放闸门,能诱发痛。
此外,粗纤维传入冲动除可兴奋SG和T细胞外,还可经背索上传到高级的中枢控制系统,再反过来影响闸门控制系统。
作用系统是指接受T细胞发出冲动的较高级中枢结构。它包括二个部分,一个是感觉分辨系统,产生痛感觉的成分。另一个是情绪激动系统,主要产生痛的情绪部分。
闸门学说认为,有些中枢活动如焦虑、注意,可通过下行系统开放或关闭脊髓闸门来控制疼痛的感受。这种影响范围很广,有的可开放或关闭从身体任何部分来的传入冲动的闸门,另一些显然只涉及局限性的闸门活动。
支持闸门学说的证据有:
❶在临床上刺激粗纤维可以镇痛,表明粗纤维兴奋可关闭闸门而达到镇痛作用。
❷它能比较满意地解释临床多种疼痛症状的原因,如幻肢痛病人,机械刺激可触发幻肢痛,可能系空间综合造成T细胞的兴奋加强所致。
❸有的实验表明背角中确有相当于T细胞的神经元,它接受粗细纤维的会聚性输入,并对多种感觉刺激发生反应。但具有严格的特异性的即只对伤害刺激起反应的T细胞却极罕见。
尽管上述事实支持闸门学说,但它仍受到许多学者的反对。其根据有:
❶闸门学说的关键是刺激细纤维时产生突触前容易化而表现为正的背根电位。不少生理家曾致力于寻找这种正的背根电位,如Zimmerman等发现刺激皮神经中的Aδ和C纤维并不产生正的背根电位。Franz和Iggo亦得到类似的结果。为此他们认为:闸门学说提出的刺激细纤维能产生突触前容易化缺乏生理学基础。
❷Vylicky等考虑到电刺激皮神经不是自然痛刺激,而改用辐射热刺激猫爪。这种刺激主要能兴奋C纤维,但观察到的仍是负的背根电位。Burke等用类似的辐射热致痛的方法从未获得正的背根电位,这些事实都和闸门学说不符。
Wall和Melzack最近对闸门学说分别作了不同程度的修正。Wall认为,
❶某些细纤维(Aδ和C)只对伤害刺激起反应,而另一些阈值较低的粗纤维,若刺激达到伤害水平时,它们的放电频率就增加;
❷脊髓和三叉神经核中的痛敏神经元,亦可被传导非伤害性信号的外周神经容易化或抑制;
❸来自脑的下行控制系统可调节传递伤害信号的神经元的兴奋性。Melzack认为不仅粗纤维可以镇痛,在一定条件下,刺激细纤维可能亦有镇痛作用。这表明闸门学说本身还正在不断发展、补充和完整。
痛觉pain
由伤害有机体的各种刺激所引起的感觉。使有机体产生伤害的刺激可能是机械的、温度的、电的和化学的,只要这些刺激达到了破坏有机体的强度,都可能引起痛觉。它的感受器主要是分布在皮肤下和内脏器官上的游离神经末梢。身体的一切组织和器官都能产生痛觉。它的机制比较复杂,一般认为,游离神经末梢受到伤害后产生神经冲动,沿着脊髓丘脑侧囊到达丘脑,再投射到大脑皮层的中央后回,产生痛觉。它是高等动物和低等动物共同具有的一种最原始的感觉,其生物学意义在于它是危险的警报系统,引起机体的防卫反应。在身体的不同部位,痛点(痛觉最敏感的点)的分布不同,如脚掌、手掌、腘窝、鼻尖上的痛点较少,胸部、背部、脸颊的痛点较多。人的痛觉的感受性有较大的个别差异,受情绪、动机、经验、暗示、年龄等多种因素的影响。在战场上受重伤的士兵很少作痛的反应,只有1/3的伤兵需要止痛剂。在临床条件下,有35%的病人在应用安慰剂后可以减轻疼痛。
痛觉pain sensation
感觉的一种。有机体受到伤害性刺激所产生的感觉。痛觉种类很多,可分为皮肤痛,来自肌肉、肌腱和关节的深部痛和内脏痛。通常可伴有某种生理变化和不愉快的情绪反应。人的痛觉存在个别差异。有人痛感受性低,有人则高。痛觉与心理因素关系很大。在民族、性别、年龄方面也存在着一定的差异。影响痛觉的心理因素主要是注意力、态度、意志、个人经验、情绪等。痛觉是有机体内部的警戒系统,能引起防御性反应,具有保护作用,亦是疾病诊断的重要参考因素。
痛觉
机体对各种伤害性刺激的感觉。由体表和一些机体内部组织(如肌肉、肌腱、关节、内脏等)上的游离神经末梢受到刺激,产生兴奋,传入大脑皮层而引起。大脑皮质对痛觉可起调节作用。当刺激达到一定强度时,常伴有痛反应(如局部肌肉收缩、呼吸暂停或加快、出汗等)。痛觉包括皮肤痛,来自肌肉、肌腱、关节等的深部痛以及内脏痛等。痛觉的适应性很小。
- mudras是什么意思
- mudristakumudacandraprakarana是什么意思
- mudsill是什么意思
- mudskipper是什么意思
- mudslinging是什么意思
- mud snake是什么意思
- mud turtle是什么意思
- mudung是什么意思
- mudvalan是什么意思
- mud是什么意思
- muet是什么意思
- mufaddaliyat是什么意思
- muf dif是什么意思
- muff是什么意思
- muffetee是什么意思
- muffle是什么意思
- muffler是什么意思
- muf guil yinx de是什么意思
- mufti是什么意思
- mufti haji-el-husaini是什么意思
- mug是什么意思
- mu ga是什么意思
- mugabe,robert gabriel是什么意思
- mugdhabodha是什么意思
- mugger是什么意思
- mugging是什么意思
- muggy是什么意思
- mughal是什么意思
- mughal art and architecture是什么意思
- mughal-e-azam是什么意思
- mughal empire是什么意思
- mughal([ˈmu:gʌl],[muˈgʌl])dynasty是什么意思
- mughal,empire是什么意思
- mugwump是什么意思
- mugwumps是什么意思
- muhamd ibn-qasim是什么意思
- muhammad是什么意思
- muhammada是什么意思
- muhammad abdur rahman chaihtai是什么意思
- muhammad adil sha sur是什么意思
- muhammad ali是什么意思
- muhammadan是什么意思
- muhammad bin tughluk是什么意思
- muhammad jaisi是什么意思
- muhammad of ghur是什么意思
- muhammad rail sauda是什么意思
- muhammad reza shah pahlevi是什么意思
- muhammad riza是什么意思
- muhammad shah是什么意思
- muhammad sultan是什么意思
- muhammad tughlaq是什么意思
- muhammad ⅰ是什么意思
- muhammad ⅱ是什么意思
- muhammad ⅳ是什么意思
- muhammad ⅴ是什么意思
- muhammad ⅵ是什么意思
- muharram是什么意思
- mu hka是什么意思
- mu hkungri是什么意思
- muhkup是什么意思