生物膜与能量转换
生物膜与能量转换
生物体内的能量转换形式很多,例如,光能转换为化学能——光合作用; 化学能转换为化学能——线粒体氧化磷酸化,化学能转换为电能——电鳗生电,化学能转换为光能——萤火虫发光等等。生物体内重要的能量转换过程都与膜结构密切有关。例如,光合作用的主要过程是在植物细胞叶绿体的类囊体膜进行的。有细胞‘动力站’之称的线粒体内膜则是进行氧化磷酸化的主要场所。脊椎动物光感受器膜将光能转换,并经过一系列复杂的过程使之变为电脉冲传给大脑。原核细胞的氧化磷酸化过程在细胞质膜或其内陷的膜结构中进行。嗜盐菌的质膜既能将光能转换为化学能,也能进行氧化磷酸化。
线粒体内膜与氧化磷酸化 一切异养生物能量都来自食物。食物通过一系列的降解转化,最后通过呼吸作用把内含的能量转化为可利用的形式加以贮存或供各种活动的需要。
线粒体是细胞进行呼吸作用的重要细胞器。它有内、外两层膜。内膜向线粒体的中心腔内陷形成嵴(图1)。线粒体含有催化三羧酸循环、脂肪酸氧化、氨基酸分解、电子传递、偶联磷酸化等有关的酶和辅酶共约100种。它们有序地构成高度组织化的系统来进行各种反应。催化三羧酸循环、脂肪酸氧化、氨基酸分解的有关酶和酶系分布在基质中。很多脱氢酶、电子传递链和偶联磷酸化的酶系都位于内膜。生物氧化释放能量主要发生在线粒体内膜进行的电子传递过程。
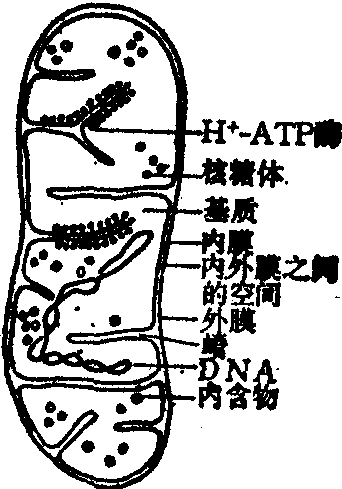
图1 线粒体模式图
1. 电子传递链:细胞内代谢通过三羧酸循环产生还原型烟酰胺腺嘌呤二核苷酸(NADH)。NADH以及一些直接以异咯嗪为辅基的脱氢酶的底物(如琥珀酸)最后通过电子传递链而被氧氧化。在此过程中释放出自由能。电子传递链是线粒体内膜的主要组份。经过四五十年的研究,虽然对电子传递链的组份及其序列还存在着不同的看法,各种来源的线粒体也有一定的差异,但下列电子传递链简图是为大多数人所接受的:

Green和Hatefi成功地将呼吸链分成四个复合物Ⅰ,Ⅱ,Ⅲ,Ⅳ,在一定条件下拆离的复合物可以进行重组,并基本上恢复活力,它们的序列如下图2所示。
电子传递链在线粒体内膜两侧的分布是不对称的。例如,琥珀酸脱氢酶,NADH脱氢酶位于基质一侧。细胞色素c、细胞色素c1位于胞质一侧。细胞色素氧化酶则跨膜分布。
2. 氧化磷酸化:底物通过电子传递链氧化所释放的能量转换为可供利用的能源物质三磷酸腺苷 (ATP),称为氧化磷酸化。
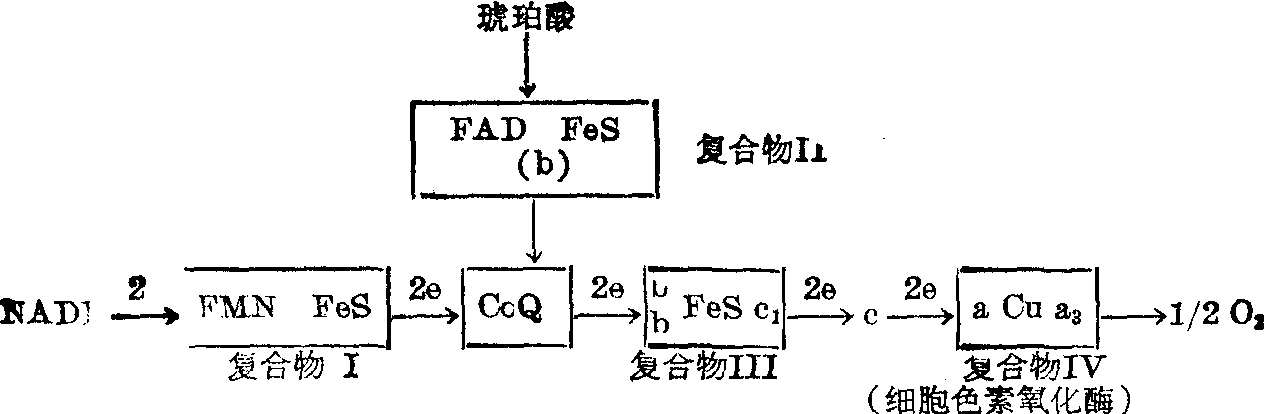
图2复合物Ⅰ~Ⅳ在电子传递链的序列
FMN,黄素单核苷酸; FAD,黄素腺嘌呤二核苷酸; FeS,铁硫中心。
一对电子(或氢)自琥珀酸传递到O2,每消耗一原子氧,可形成2个ATP分子,自NADH传递到O2,则可合成3个ATP分子。每消耗1g原子氧所产生的ATP摩尔数可以P/O比值表示之,称为氧化磷酸化效率。
整个电子传递链有三个氧化磷酸化偶联部位,第一偶联部位介于NADH→GoQ之间,此外,CoQ→细胞色素c和细胞色素c→O2之间分别各有一个偶联部位。
3. 三磷酸腺苷酶复合体(H+-ATP酶)催化电子传递链的磷酸化: 在电子显微镜下用负染法可以观察到线粒体内膜的内表面有许多直径为90A的球状颗粒,这就是电子传递过程中催化合成ATP的ATP酶复合体(图3)。它由三部分组成:
❶可溶性ATP酶(F1),分子量约360,000,至少由五种亚单位组成(α,β,γ,δ,ε),F1只有与ATP酶复合体其他部分结合后才表现出对寡霉素的敏感性;
❷赋予寡霉素敏感蛋白质(OSCP),分子量约18,000;
❸疏水蛋白(F0),含四种亚单位,分子量约70,000,具有质子通道的功能。此外,还有一个分子量约为10,000的多肽,它是ATP酶的天然抑制剂,专一地抑制F1的水解活力,可能在正常生理条件下起着调节酶活的作用。
H
 -ATP酶的分布很广泛,除线粒体内膜外,叶绿体类囊体膜、细菌质膜等都有分布,它们的组成与结构基本相似。
-ATP酶的分布很广泛,除线粒体内膜外,叶绿体类囊体膜、细菌质膜等都有分布,它们的组成与结构基本相似。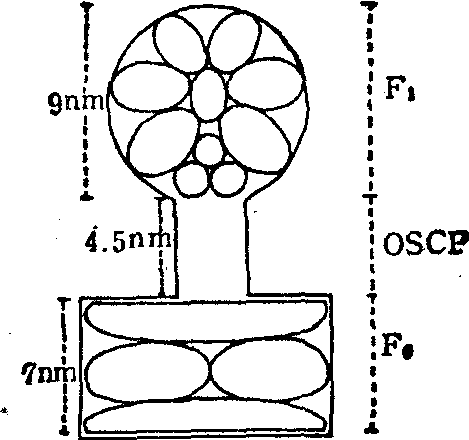
图3 ATP酶复合体各组份的可能排列
4. 氧化磷酸化的偶联机理:
(1) 化学假说: 1953年荷兰Slater提出,主要参照在糖酵解过程中
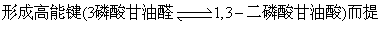 出的。根据这个假说,电子传递时高能磷酸键的形成是由于化学能的直接转换。电子传递过程释放的自由能直接转入ATP的前体而后再生成ATP。化学假说可以下式来表示:
出的。根据这个假说,电子传递时高能磷酸键的形成是由于化学能的直接转换。电子传递过程释放的自由能直接转入ATP的前体而后再生成ATP。化学假说可以下式来表示: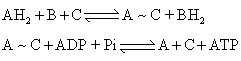
化学假说在氧化磷酸化研究的早期曾占很重要的位置。但主要的问题在于迄今始终未能分离出含高能键的中间递体。
(2)构象假说: 认为电子传递释出的能量首先不是贮存在共价键而是积贮于蛋白质肽键的立体结构之中。构象假说可用下列反应来表示:
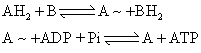
(3)化学渗透假说: 英国Mitchcll于1961年提出。这个假说的特点是突出膜的结构。Mitchell假设呼吸在线粒体内膜上构成三个回路 (图4),电子与氢的交替传递使质子发生定向转移,从膜的一侧运送到另一侧。由于膜对H+是不通透的,因而使膜内外形成了质子动力势或称质子电化学梯度(⊿
 H+), 它包括膜电位(⊿ψ),和膜内外的pH差(⊿pH)。所以可以将呼吸链看作是质子泵,而嵌在线粒体内膜上的H+-ATP 酶则利用以质子电化学梯度形式贮存的能量使ADP和Pi合成ATP,从而使质子发生逆向回流,质子梯度也随之消失。因此,化学渗透假说可简单概括如下式:
H+), 它包括膜电位(⊿ψ),和膜内外的pH差(⊿pH)。所以可以将呼吸链看作是质子泵,而嵌在线粒体内膜上的H+-ATP 酶则利用以质子电化学梯度形式贮存的能量使ADP和Pi合成ATP,从而使质子发生逆向回流,质子梯度也随之消失。因此,化学渗透假说可简单概括如下式: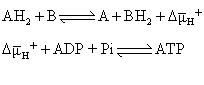
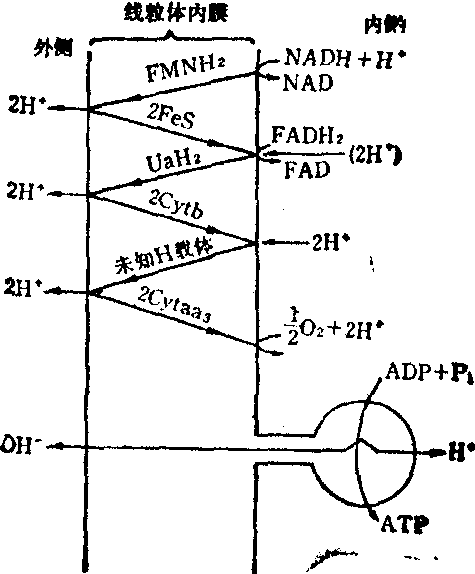
图4化学渗透假说示意图
UQ,泛醌(辅酶Q); Cytaa3,细胞色素c氧化酶。
Mitchell假说的贡献在于对氧化磷酸化的偶联机理研究并没有受传统的化学概念所束缚,而提出偶联过程涉及一个物理现象,也即电子传递所释放的能量转化形成膜两侧的化学梯度(⊿ pH)和膜电位。它们可以通过ATP酶复合体的催化导致ATP的合成,这些假设迄今为止已被大量实验所证实,除线粒体内膜的氧化磷酸化外,
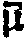 1+也是叶绿体类囊体膜、好氧细菌质膜、 嗜盐菌紫膜进行能量转换过程的中间形式。因此,化学渗透假说有力地推动了生物能转换机理的研究,Mitchell也因此获得了1978年诺贝尔化学奖。但近年来陆续报道与Mitchell假说相矛盾的实验结果,一些科学家又提出了有关能量转换的新假说。对于ATP酶复合体合成ATP的细节迄今仍了解很少。
1+也是叶绿体类囊体膜、好氧细菌质膜、 嗜盐菌紫膜进行能量转换过程的中间形式。因此,化学渗透假说有力地推动了生物能转换机理的研究,Mitchell也因此获得了1978年诺贝尔化学奖。但近年来陆续报道与Mitchell假说相矛盾的实验结果,一些科学家又提出了有关能量转换的新假说。对于ATP酶复合体合成ATP的细节迄今仍了解很少。脊椎动物光感受器膜的能量转换 外界各种刺激 (或信息)作用于各种感受器后,都要先把它们分别转变成神经系统能进行加工处理和传递的电信号,因此也可以把感受器看成是一种“换能器”。对视觉系统来说,当光子作用于视细胞时由视细胞外段——光感受器膜将光信号转变成感受器电位,然后通过突触,传递到双极细胞、神经节细胞,并变成脉冲形式的神经冲动经视神经最后到达大脑产生视觉。
脊椎动物的视网膜中有二种视细胞: 视杆细胞和视锥细胞。视杆细胞的外段含有几百个互相重叠的扁平圆盘体,它们具有双层膜结构,圆盘膜厚75~80A,视紫红质镶嵌其中,它们约占膜蛋白的87%。视锥细胞与视杆细胞的结构很相似。视紫红质由视蛋白和11-顺-视黄醛组成,含348个氨基酸残基,分子量为39,048,整个分子由7条跨膜多肽组成。分子中的11-顺-视黄醛的侧链含有四个双键,其中三个呈反式第四个(11,12位)呈顺式。视黄醛通过质子化的希夫碱键与视蛋白赖氨酸残基相结合。视黄醛是维生素A的衍生物 (即维生素A侧链末端—CH2OH被—CHO替代)。
视觉过程的分子基础简述如下:
1. 感光过程的原初反应: 当光量子被视网膜视杆细胞外段圆盘膜吸收后,视紫红质中的视黄醛发生异构化(即11-顺-视黄醛经过一系列反应变成全-反-视黄醛)(图5)。这纯粹是光化学反应,整个过程的第一步反应在6皮秒(6ps,即6×10-12s)内完成,而其余步骤都可在暗中继续进行。

图5 视紫红质漂白的光化学过程括号中数字为该物质的吸收峰值
当11-顺-视黄醛转变成全-反-视黄醛后即与视蛋白解离(图6)。
在暗中,通过两个酶的催化,全-反-视黄醛又重新形成11-顺-视黄醛,然后,11-顺-视黄醛再与视蛋白结合重新形成视紫红质。

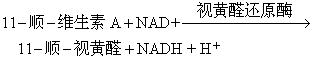
2. 光刺激的继发传递: 对于光信息如何转变为感受器电位的机理曾提出很多假说,其中Hagin等人所提出的离子假说比较流行。他们认为,在脊椎动物光感受器的细胞质内Ca2+的浓度远低于感受器外部及其内含圆盘膜中的Ca2+浓度。很可能,光作用于视紫红质分子形成全-反-视黄醛后,使之从与视蛋白的结合部位释放出来,随即与圆盘膜和质膜的一定组份的氨基形成希夫碱,从而增加对Ca2+的通透性,使光感受器外和圆盘膜中的Ca2+较多地进入细胞质内。它们与质膜结合后,使Na+通道关闭,从而产生超极化现象,形成感受器电位。这样Ca2+也就起到了把视紫红质分子光化学反应的信息从圆盘膜传递到视细胞外段的质膜的作用。假如一个视紫红质分子接受一个光子之后能使很多Ca2+进入光感受器的细胞质内,而每一个Ca2+又能阻止很多的Na+流入光感受器,这样就能从量子增益的角度来解释光感受器的光电转换过程中的巨大放大作用。
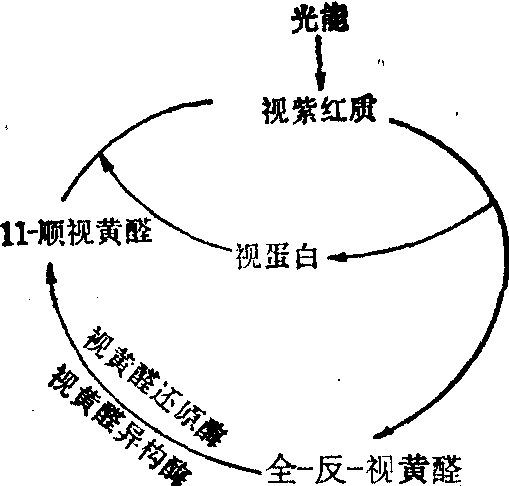
图6 视杆细胞的视循环
近年来又有一些研究者指出,在光刺激后,视杆细胞外段中的环化GMP(c-GMP)的含量会迅速下降,它可能起着调节视杆细胞外段质膜对离子通透性的作用。当视紫红质捕获光子后,脊椎动物视杆细胞外段质膜Na+通道被关闭也可能与此有关。
嗜盐菌紫色膜的能量转换 嗜盐菌是杆状细胞,直径约0.5μm,长度约5μm,最适于在4.3mol/L的NaCl(25%)溶液里生长(海水盐浓度一般为0.6mol/L)。在盐的浓度低于3mol/L时,嗜盐菌即难以存活。
1967年,美国Sto
 ckenius等发现嗜盐菌质膜含有一种细菌视紫质(菌紫质),也能将光能转化为化学能,这是一种无需叶绿体的光合作用,它使嗜盐菌在含氧低,光线强,渗透压大的环境中满足细菌进行新陈代谢及维持其细胞完整的需要。
ckenius等发现嗜盐菌质膜含有一种细菌视紫质(菌紫质),也能将光能转化为化学能,这是一种无需叶绿体的光合作用,它使嗜盐菌在含氧低,光线强,渗透压大的环境中满足细菌进行新陈代谢及维持其细胞完整的需要。嗜盐菌的细胞质膜呈暗红色,从中可分离一种红色和一种紫色膜部分。当O2供应充足时,大部分为红色膜,而在缺O2情况下,紫色部分可增至50%。
嗜盐菌在无氧条件下可以直接利用太阳能合成ATP。在黑暗条件下如果供应O2,也可通过呼吸作用形成ATP。嗜盐菌紫膜只含有一种蛋白质称为菌紫质,约占紫膜的75%,其余为脂质。与视紫红质相似,菌紫质由蛋白质和色素分子——视黄醛组成。每分子菌紫质含一分子视黄醛,它通过希夫碱基与蛋白质相连结。菌紫质的蛋白质部分的分子量为26,000,含235个氨基酸残基,一级结构已基本分析完成。分辨率为7A的电子衍射研究结果表明,每个跨膜分布的菌紫质分子含7条几乎平行的柱形多肽,它们呈α螺旋结构,长35~40A,相隔10A,首尾相接,垂直于膜平面。其一COOH末端暴露于细胞质一端,一NH2末端则在细胞外一侧。
菌紫质受光照后跃迁到激发态,然后将自由能逐步释放,依次转变为各种中间物,最后恢复至原来的基态。整个光化学过程极为复杂。在这样的能量转换过程中,紫膜起着质子泵的作用,把膜内一侧的质子泵到另一侧,从而产生质子电化学梯度(⊿
 H+),它可以通过嗜盐菌质膜的H+—ATP酶合成ATP。
H+),它可以通过嗜盐菌质膜的H+—ATP酶合成ATP。☚ 生物膜的分子结构 生物膜与物质运送 ☛
- 蒙自仁是什么意思
- 蒙自南湖是什么意思
- 蒙自木蓝是什么意思
- 蒙自木蓝是什么意思
- 蒙自木蓝是什么意思
- 蒙自虎耳草是什么意思
- 蒙自虎耳草是什么意思
- 蒙自虎耳草是什么意思
- 蒙自赤杨是什么意思
- 蒙自赤杨是什么意思
- 蒙自赤杨是什么意思
- 蒙舍是什么意思
- 蒙花皮是什么意思
- 蒙花皮是什么意思
- 蒙花皮是什么意思
- 蒙茏是什么意思
- 蒙茸是什么意思
- 蒙茸是什么意思
- 蒙药制剂学是什么意思
- 蒙药图鉴是什么意思
- 蒙药学是什么意思
- 蒙药学是什么意思
- 蒙药学是什么意思
- 蒙药志是什么意思
- 蒙药方剂是什么意思
- 蒙药方剂是什么意思
- 蒙药炮制学是什么意思
- 蒙莉莎时装有限公司是什么意思
- 蒙蒂是什么意思
- 蒙蒂是什么意思
- 蒙蒂是什么意思
- 蒙蒂斯是什么意思
- 蒙蒂菲奥里是什么意思
- 蒙蒙是什么意思
- 蒙蒙夕照开棠邑,叶叶风帆下建康。是什么意思
- 蒙蔽是什么意思
- 蒙藏《格萨尔》的关系是什么意思
- 蒙藏事务处是什么意思
- 蒙藏关系史研究是什么意思
- 蒙藏合壁智慧之源是什么意思
- 蒙藏周报是什么意思
- 蒙藏委员会是什么意思
- 蒙藏委员会建议整顿蒙古交通是什么意思
- 蒙藏民族关系史略是什么意思
- 蒙藏院是什么意思
- 蒙藏院是什么意思
- 蒙袂辑屦是什么意思
- 蒙袂辑屦是什么意思
- 蒙被是什么意思
- 蒙被子放屁——独吞是什么意思
- 蒙诏帖是什么意思
- 蒙诏帖是什么意思
- 蒙译修辞是什么意思
- 蒙语词汇学探索是什么意思
- 蒙语诠释是什么意思
- 蒙语语法研究是什么意思
- 蒙诺阿日是什么意思
- 蒙读唐诗鉴赏辞典是什么意思
- 蒙谤而晏如,不慑惧于可畏者,劲人也。是什么意思
- 蒙贝仪式是什么意思