生物膜与信息传递
信息传递过程包括信息分子的产生、识别,接受和传递。细胞表面接受细胞外的信息,并将它传至细胞内部,因而起着传递信息的中介作用。细胞识别,细胞免疫,神经传导,激素作用,凝集素作用都牵涉到细胞表面的信息传递功能。
受体 主要是指存在于细胞膜表面的一种或一类分子,通常是嵌入膜内的内在蛋白质(糖蛋白或脂蛋白),少数是脂质或糖脂分子。它们能识别、结合专一的生物活性物质(或称配体),生成的复合物能激活和启动一系列变化,从而导致该活性物质的最终生物效应。也有些受体不分布于细胞膜表面而位于胞浆或胞器内。受体能高度选择性地与特定的分子可逆地结合。这种结合不仅具有很强的亲和力,而且具有严格的立体化学结构特异性。这种结合能被饱和,并被专一的拮抗剂所阻断,这种物质称为“受体拮抗剂”,与受体结合引起药理效应的物质称为“受体激动剂”。
1. 受体的数量和分布:一种细胞膜可以含有几种不同的受体,如脂肪细胞膜上含有肾上腺素、胰高血糖素、胰岛素等近10种激素受体。它们的数目互不相同。同一受体在不同细胞膜上的受体数目也不相同。一般讲,每一个细胞膜上含有103~104个受体,乙酰胆碱的受体的密度和数量较大,分别为104~105个/μm2和1011个/细胞。受体的数目在正常生理条件下是恒定的。随着细胞生理状态的变化(如生长速度,分化程度,细胞周期等)和环境的影响,受体的数目也会发生一定的改变。
2. 受体的分离与结构: 由于细胞内受体含量极微,有些受体稳定性又差,因此受体的分离、纯化一直比较困难。迄今只有从Torpedo电鱼或电鳗的电器官中分离的乙酰胆碱的烟碱胆碱能受体和从正常人胎盘中分离的胰岛素受体已经得到纯品,从而对它们的结构才有较多的了解。
乙酰胆碱的烟碱胆碱能受体各亚基的氨基酸序列已经搞清,受体的mRNA已在无细胞体系及细胞中翻译成功。这种受体由四种共五个亚基组成,分子量为250~270×103。电子显微镜下呈玫瑰花瓣状结构,直径为80~90,中间有直径为25的小孔,可能是离子通道。
胰岛素受体的Stock半径为68~72A,分子量为30~100万,它主要由三种亚基: α(分子量约135,000),β(分子量约95,000)和γ(分子量约45,000)组成。α亚基为结合亚基,亚基之间靠—S—S—相连,亚基内部还有—S—S—结构。
3. 受体假说: 1978~1980年间,英国生理学家Lang-ley通过对神经系统药物和递质作用机理的研究,提出受体假说,企图阐明药理作用的原始反应。其主要论点是:当内、外源性生物活性物质到达机体各部分器官、组织时,在绝大多数情况下,都先与细胞膜表面的相应受体结合,从而引发一系列的反应,最终产生各种生理效应、药理作用和毒理反应,可用下式简单表示:
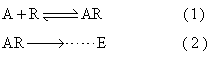
A为激动剂,R为受体,E表示效应。这一假说在发展过程中,先后提出过“受体占领假说”,“结合速率假说”和“诱导契合假说”等。最近又发展为“移动受体假说”。
激素作用 激素由内分泌细胞产生,作为“化学信息”通过体液或细胞外液运送至靶细胞后产生专一性的效应来调节各种物质代谢和生理功能。有些激素 (如甾体激素、甲状腺素)须进入细胞与胞质内的受体相结合然后起作用。但很多激素作用时并不需要进入细胞而仅与细胞膜表面的受体相结合,导致其构象变化,继而再使膜上的腺苷酸环化酶活化使细胞内含的ATP合成环化腺苷酸(cAMP),后者引起一系列酶反应而产生生理效应。因此将激素称为第一信使,而将cAMP称为第二信使。例如,肾上腺素是由肾上腺髓质分泌通过血液输送至肝细胞产生效应。待它们与肝细胞表面受体相结合后,能使膜上腺苷酸环化酶活化产生cAMP,后者又使蛋白激酶等一系列酶蛋白相继活化,最终使糖原分解成葡萄糖从而使血糖浓度升高。其他一些激素,如促肾上腺皮质激素(ACTH)、促卵泡激素(FSH)等在作用时也都分别与它们的靶细胞受体相结合,导致腺苷酸环化酶的激活而产生cAMP。各种激素在细胞膜上具有各自专一的受体。1977年Jacobs和Cuatrecases假设腺苷酸环化酶并不始终与激素受体相偶联,换言之,激素受体不止一种,但细胞膜上的腺苷酸环化酶却是共用的。待激素分子与膜受体相结合形成复合体后,它们在膜上进行侧向移动,一旦它们与腺苷酸环化酶相偶联,才能导致后者的构象变化而被激活。这称之为移动受体假说。
近年来发现有些激素(如胰岛素)在作用时首先先与靶细胞膜的受体相结合,然后胰岛素、受体复合物通过吞噬作用一起进入细胞内部,继而与溶酶体融合,或与其他细胞器膜相结合再产生生理效应。
神经传递 高等动物的神经系统是由数目庞大的彼此连结的神经细胞——神经元组成,人脑内约含1011神经元。神经细胞通过突触与邻近神经细胞相联系。当神经细胞受刺激而兴奋时,受刺激的那一部分膜对Na+、K+的通透性大大增加,产生去极化现象而形成动作电位,电脉冲沿着神经细胞继续传递,当传至细胞末端时,或继续传给下一个神经细胞,或到达靶细胞(如肌肉)引起反应。神经细胞与神经细胞之间或神经细胞与肌肉组织之间有一小间隙,电脉冲在正常情况下是不能通过这一间隙的。这需要释出一种称为神经传递质的化学物质,它们可以扩散、越过间隙,并与下一个细胞膜上的受体相结合,从而产生新的电脉冲继续传递下去或使靶细胞引起反应。
神经传递质有很多种,例如γ-氨基丁酸、5-羟色胺、谷氨酸、乙酰胆碱(ACH)等等,其中对乙酰胆碱的研究历史最长,其作用机理也比较清楚。当神经脉冲传至突触前膜时,突触细胞质内大量的储存泡通过与突触前膜相融合而将所含的乙酰胆碱释放至距离为0.5nm的突触隙,之后乙酰胆碱与突触后膜上的受体相结合,导致后者的构象变化使突触后的神经细胞膜的Na+、K+通道开放,发生去极化,从而使神经兴奋继续传递下去(图)。在突触膜上还分布有乙酰胆碱酯酶,它可使乙酰胆碱迅速分解,从而使乙酰胆碱受体回复原来构象,恢复原有的离子运送屏障,重新回至静止状态。
释放的乙酰胆碱如果作用于靶组织(如肌肉),就会引起反应(如收缩)。当乙酰胆碱与肌肉细胞膜受体结合后,会引起它的构象发生变化,打开离子通道,使膜对Na+、K+的通透性瞬时增加,导致膜的去极化,使肌肉收缩。一个与乙酰胆碱结合后的受体可导致5×104离子通过。
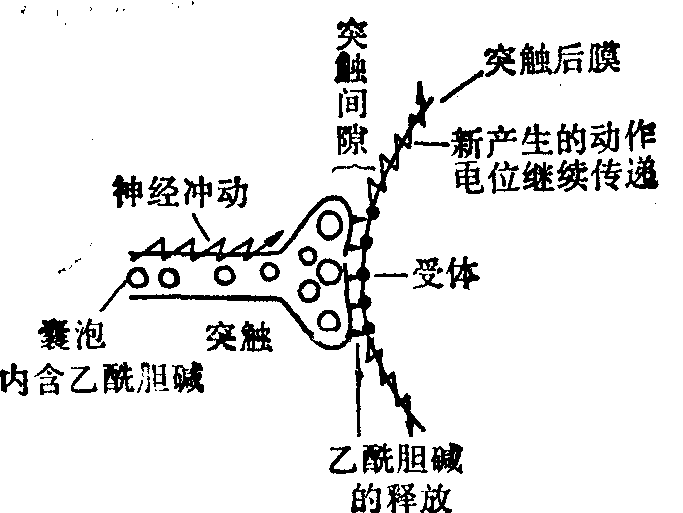
乙酰胆碱的释放及其与受体的结合(自J.B.Finean)
毒素作用 很多毒素也是通过与细胞膜上的受体相结合后才产生效应的。以霍乱毒素为例,它是霍乱弧菌产生的外毒素,分子量为84,000,由A、B二种亚单位组成。A亚单位有两条肽链A1和A2,由一对二硫键相连接。亚单位B与细胞膜上的受体相结合。亚单位A1则具有激活膜上腺苷酸环化酶的作用。
霍乱毒素的受体是一种神经节苷脂,毒素与它结合后,可能会发生如下的系列反应,首先引发受体的构象变化,接着亚单位A1在激活腺苷酸环化酶的过程中将烟酰胺腺嘌呤二核苷酸(NAD) 中的腺二磷核糖转移到细胞膜中的一种蛋白质上。在正常情况下,鸟三磷通过与这种蛋白质相结合使腺苷酸环化酶激活,待鸟三磷被鸟三磷酶水解,激活作用即停止。但是,如果鸟三磷结合在含有腺二磷核糖的蛋白质上就不易被水解,因而延长了腺苷酸环化酶作用的时间,产生的生物效应的持续时间也比较长。
霍乱毒素的作用与激素有几点不同:
❶激素的启动时间很短,霍乱毒素常常要延迟1~3刻钟才开始;
❷激素去除后,效应很快消失,而霍乱毒素在去除后生物效应还可持续几小时到几天;
❸一种激素的受体只在少数几种细胞膜中存在,而霍乱毒素的受体在很多细胞膜上都有分布。
凝集素的作用 凝集素是从各种植物或低等动物组织中分离而得的一类特殊蛋白质,能与动物细胞膜表面的受体发生特异性结合并产生一系列的生理效应。由于它们通过与细胞表面寡糖结构决定簇的交互作用,诱导细胞发生凝集,因而称为凝集素。因大多从豆类中分离而得,又称植物凝集素。最早发现它们能诱导红细胞发生凝集而用于临床血型分类,故又有血凝素或植物血凝素之称,后者如今已作为从红肾豆中分离的一种凝集素的专门名称(PHA)。目前已报道过的凝集素多达500种,不仅从豆类,而且可以从担子菌、软体动物、甚至蟹类组织中分离得。不但用于临床血型分类,也用于下列研究:
❶正常细胞和肿瘤细胞表面的结构;
❷分离细胞膜糖蛋白、寡糖及粘多糖;
❸丝裂作用;
❹作为抗原-抗体反应的白、寡糖及粘多糖;
❸丝裂作用;
❹作为抗原-抗体反应的一种模式等等。
虽然凝集素的种类很多,但目前研究得较透彻,并肯定具有严格专一性作用者不过十余种,如伴刀豆素A(ConA)、大豆凝集素(SBA)、香菇凝集素(LEA-A,-B)、金白蘑菇凝集素(ABA-A)、莲子凝集素(LA-A,-B,-C)、利马豆凝集素(LBA)、植物血凝素(PHA-L,-H)、商陆凝集素(PWA)、豌豆凝集素(PA)、蓖麻凝集素(RCA-Ⅰ,-Ⅱ)及麦胚凝集素(WGA)等。
凝集素受体 与凝集素特异结合的受体位于膜表面,具有复杂的寡糖链结构,它们拥有特殊的联结、侧链及多肽附着物,在这些寡糖链间还存在非共价键,如氢键的相互作用。凝集素与受体寡糖链的结合部位,以往多认为是终端的糖残基,现在证实还能与多糖核心部位相结合,甚至更为重要。曾发现核心顺序 (D-甘露糖)3-D-N-乙酰-葡萄糖胺-D-N-乙酰-葡萄糖胺是多种凝集素的结合部位。看来,不同凝集素的情况也不尽相同,例如,RCA仅与终端残基相互反应,Con A及LEA则与终端及核心都有作用。PHA也与外侧链D-半乳糖残基和核心的D-甘露糖残基都能结合。LEA可与受体的不同部位结合,它们都含有N-乙酰-D-葡萄糖胺与D-甘露糖残基的结构。
不同的凝集素可以各有互不相同的专一性受体,如WGA和Con A的受体不一样。也可以具有共同的受体,如人红细胞血型糖蛋白既是Con A又是RCA的受体。多数情况下,同一凝集素在各种细胞上结合的受体寡糖键结构基本相同,如WGA在L1210、PV 3T3及PYBHK三种细胞具有相同的受体,但在淋巴细胞的受体结构却不一样。
在细胞内膜系统中,核膜也存在凝集素的结合部位,如Con A、RCA-1、WGA和PHA等。对线粒体膜与凝集素结合的问题也有过少数报告。
当凝集素与其专一膜受体结合后可以诱导细胞出现多种生理效应,如细胞发生凝集,特别是肿瘤或转化细胞可以发生高效价的凝集作用;诱导淋巴细胞发生丝裂现象;产生胰岛素样的生理作用以及对受精过程、细胞表面受体运动、膜透性、吞噬及细胞增殖等都可产生一定的影响。
- 是非只为多开口,烦恼皆因强出头是什么意思
- 是非只为多开口,祸乱都因硬出头是什么意思
- 是非只是个善恶是什么意思
- 是非曲直是什么意思
- 是非曲直是什么意思
- 是非曲直是什么意思
- 是非曲直是什么意思
- 是非有分,以法断之是什么意思
- 是非法是什么意思
- 是非终日有,不听自然无是什么意思
- 是非自有公论是什么意思
- 是非题是什么意思
- 昱是什么意思
- 昴是什么意思
- 昴降(昴星)是什么意思
- 昵是什么意思
- 昵是什么意思
- 昵呢儿女语,恩怨相尔汝。划然变轩昂,勇士赴敌场是什么意思
- 昼是什么意思
- 昼上人集是什么意思
- 昼光是什么意思
- 昼夜是什么意思
- 昼夜兼行是什么意思
- 昼夜兼行是什么意思
- 昼夜时间利用系数是什么意思
- 昼夜永·洞房记得是什么意思
- 昼夜班计划是什么意思
- 昼夜节律是什么意思
- 昼夜节律是什么意思
- 昼夜节律是什么意思
- 昼夜蔽日月,冬夏共霜雪。是什么意思
- 昼夜证象同异辨是什么意思
- 昼夜证象同异辨是什么意思
- 昼夜证象同异辨是什么意思
- 昼尔于茅,宵尔索綯是什么意思
- 昼帘绪论是什么意思
- 昼无事者夜不梦。是什么意思
- 昼星楼医案是什么意思
- 昼有所思,夜梦其事是什么意思
- 昼永是什么意思
- 昼热是什么意思
- 昼热是什么意思
- 昼锦堂图是什么意思
- 昼锦堂记碑是什么意思
- 昼锦还乡(昼锦游、昼锦)是什么意思
- 昼间人口是什么意思
- 显是什么意思
- 显是什么意思
- 显亲扬名是什么意思
- 显亲扬名是什么意思
- 显亲扬名是什么意思
- 显亲扬名是什么意思
- 显人才是什么意思
- 显仁后回南是什么意思
- 显仁皇太后(韦氏)是什么意思
- 显佑宫是什么意思
- 显像管是什么意思
- 显像管生产现场管理是什么意思
- 显克微支是什么意思
- 显克微支是什么意思