生化反应机制的示踪研究
核素示踪技术是研究生化反应机制极为有用的手段。本条主要简述其中几个主要方面。
生化反应的可逆性 用放射性核素示踪法观察生化反应的可逆性,反应系统中前身物和产物的浓度都接近生理浓度,这对结论的可靠性非常重要。实验中加入少量带标记的产物,测前身物的放射性,即可判断反应的可逆性。
作结论时必须确定被测物的放射性是由于标记产物转化生成了标记前身物,并非由于其他因素,如示踪物吸附于非标记分子上,示踪物通过其他化学键连至非标记分子上等。所以对放射性物质的分离定性工作十分重要。如果证实某一反应确属可逆反应,还可根据反应两侧的放射性分布观察反应速度。
用上述方法对许多生化反应进行的观察表明,在生物体系中除有些是明显的不可逆反应外,大多数反应可归纳为以下几种:
❶明显可逆反应,正向和逆向由同一酶催化,条件也相似;
❷有些表面看来不可逆的反应实际上是可逆的,只是其中一个方向占明显优势,使另一个方向从定量的角度容易被掩盖;
❸有些表面看来是可逆的反应,实际上是不可逆反应,其逆向反应由另外的酶催化,而且往往所需要的其他条件(能量、金属离子等)也不同。
键的形成及打开 生物体系中有不少复分解反应,由于涉及同一种原子(或基团),其键的形成或打开的部位常并非一目了然(见图1)。第一种可能性如向上的箭头所示,第二种可能性如向下的箭头所示。要弄清何者正确,只需将其中一个反应物加标记,标记的部位在“X”上,然后分离产物测放射性。若示踪物为C—X*—D,产物中C—X*—B带标记,则知箭头向上的反应是正确的。反之,若A—X*—D带标记,则箭头向下者是正确的。
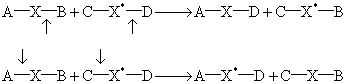
图1 复分解反应的两种可能性
典型例子是酯化反应及其逆过程(酯的水解)。键的打开或形成可以在酯的C—O间也可在O—R′间,一般方法不能区别。为此,用H218O代替普通水,使水分子上氧原子可和酯分子上的氧区别。结果18O集中在羧酸的分子上。所以酯分子上键的打开和形成是在C—O之间(见图2)。
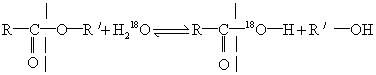
图2 酯化反应及其逆过程中键的打开和形成
还有一些用普通方法不易观察到的反应,用示踪方法可间接测知其存在,甚至测知其化学键形成或断开的方式。用18O标记的无机磷酸盐研究氧化磷酸化过程就是一个较突出的例子。例如由α-酮戊二酸经琥珀酰辅酶A形成琥珀酸,表面看来无机磷酸盐并不直接参加反应,只是在形成一个高能磷酸键时发生偶联。然而,从分子水平来看,上述反应中能量传递的机制并未阐明。实际情况很可能是琥珀酰辅酶A脱去辅酶A时先形成一个含磷酸根的中间物,后者又很快水解释出磷酸根,使GDP变为GTP,形成高能键所需的能量可能就是通过这种方式从琥珀酰辅酶A分子带到GTP上。为此制备了18O标记的磷酸盐,加入上述反应系统后确能得到含18O的琥珀酸,也就是说很可能确实存在着一个含磷酸根的瞬时性中间物。所以,在不能直接分离中间物时,这样的示踪实验可以为反应的中间过程提供有价值的论证。
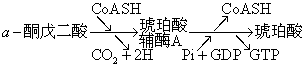
图3 α-酮戊二酸形成琥珀酸的过程
结构转变的途径 生物体内许多分子结构较复杂,而且其中许多原子是相同的。要弄清这些原子如何从简单分子中转变过来形成特殊的结构排列,常常需要针对不同目的设计一些专门的示踪实验。例如鲨烯环化有两种可能的方式(见图4)。以1-14C-乙酸为原料,生物合成鲨烯及胆固醇,用降解法测定14C在鲨烯及胆固醇分子中的分布,发现第二种转变方式和实验结果一致。若按第一种方式转变,胆固醇分子中C7及C5、C12及C13上的14C分布与鲨烯不一致。
按第二种方式形成的胆固醇,其C13上甲基从何而来又有两种可能性:
❶C8上的甲基移二位,C14上的甲基
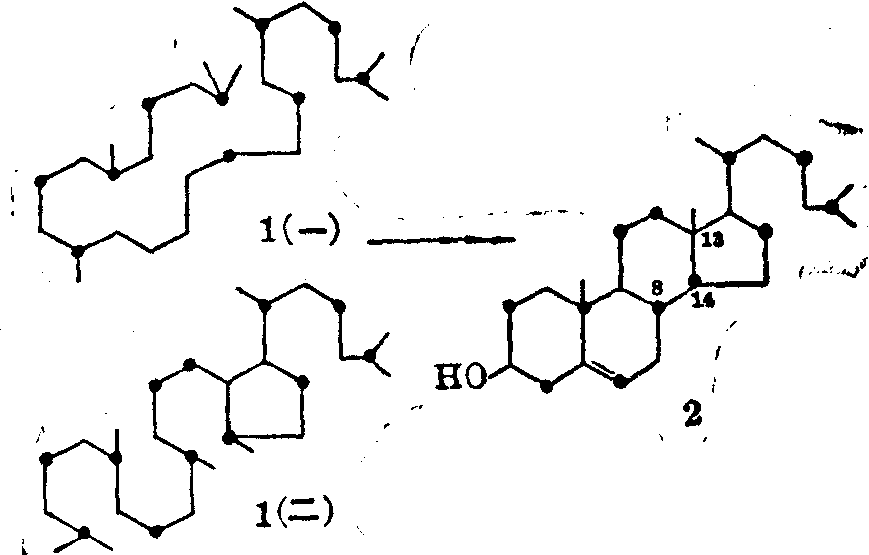
图4 鲨烯环化的两种可能途径
“●”表示由1-14C-乙酸生物合成的分子中实际测得的
14C的分布
❷C14甲基移至C12,C3上的甲基移至C14。为了阐明C12上甲基的来源,Maudgal等进行了以下实验(见图5): 以3-13C及4-13C标记的乙酰乙酸为原料,合成四种标记位置不同的鲨烯,以此混合物为前身物作参入试验,分出标记羊毛固醇。再以特殊的降解法(Kuhn-Roth方法)将所有与甲基相邻的碳氧化成醋酸,每一羊毛固醇得6分子醋酸,四种鲨烯得到24分子醋酸,再将醋酸分子转变为乙烯,用质谱仪分析其中13C的含量。结果发现每24个乙烯分子中有一个具有二个13C原子。从图可看出,若C8上甲基移两位,不可能出现这种含二个13C的乙烯分子,而若C8及C14各移一位,则每24个乙烯分子中应有一个分子含二个13C原子。由此证明后一种移位方式是正确的。
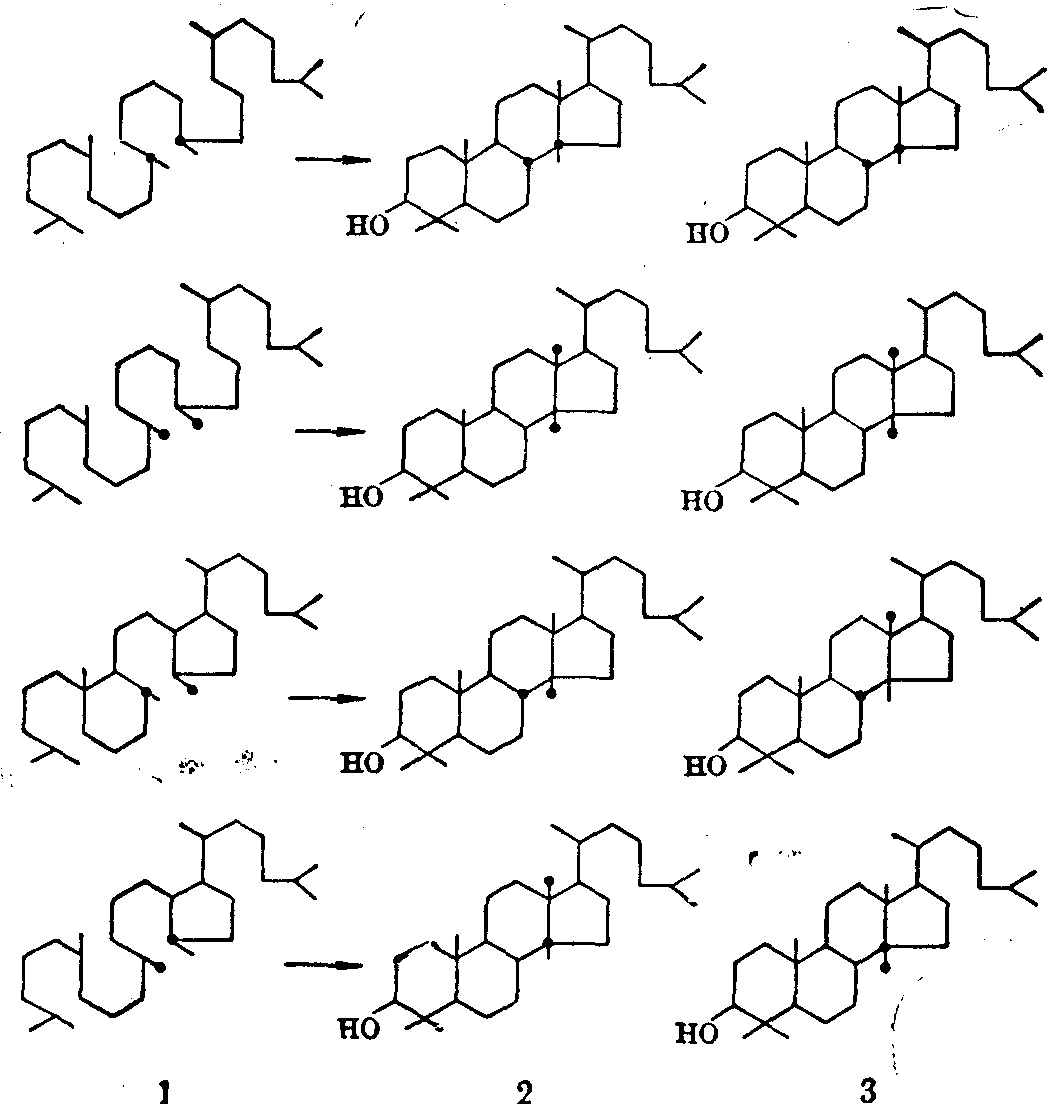
图5 胆固醇分子上第13位碳上甲基来源的实验
1.制得的标记鲨烯 2.C8上甲基移两位应得的标记羊毛固醇
3. C8及C14上甲基各移一位应得的标记羊毛固醇
上述实验也说明了稳定核素在代谢研究中有独特的地位。因为同一分子内部究竟含一个或二个标记原子,在上述实验中是无法用放射性测量技术加以判断的,只能通过质谱仪加以判断。
酶促反应动力学 生化反应有许多是多分子底物与多分子产物的反应。例如:
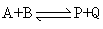
(1)顺序机制: 酶先与底物A生成复合物EA,再与底物B生成复合物EAB。然后顺序形成产物P和Q。
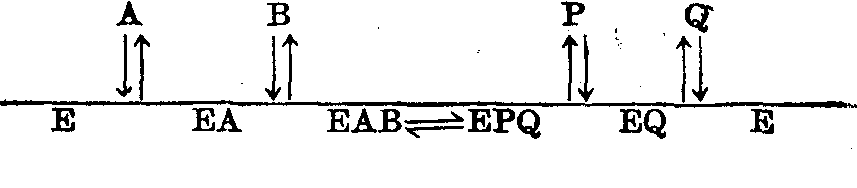
图6 酶促反应的顺序机制示意
(2)随机机制: 酶既可先与A结合,也可先与B结合,EAB形成后转变为EPQ,后者或先释出Q,或先释出P。
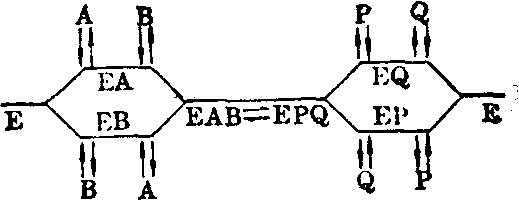
图7 酶促反应的随机机制示意
(3)乒乓机制: 在此类反应中酶可以存在二种
以上形式: 一种能与底物A结合,形成并释出P,另一种能与底物B结合,形成并释出Q。通常是两个过程相继发生,亦即酶先与A结合,释P后酶转变为另一形式,与B结合再释出Q。
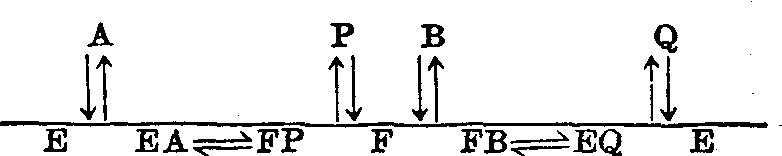
图8 酶促反应的乒乓机制示意
某一具体反应属于三种机制的那一种,可通过同位素交换法加以区分。实验在化学平衡的条件下进行,加入标记A*,观察标记原子在产物P中出现的量及速度。
在顺序机制中,没有B时反应不能完成,标记A*不能形成标记P*。但当B的浓度增大到使EA饱和(绝大部分形成EAB),EA的浓度降至极小,解离成E+A的量也极小,于是标记A*与A交换的可能性接近零,A*→P*受到完全抑制,亦即产物P中几乎测不到标记原子。
在随机机制中,没有B时反应也不能完成。但B浓度增高的影响与顺序机制不同。此处有两条通路形成EAB,虽然B浓度增大仍可使EA浓度降至极小,但另一通路却可使已经形成的EAB通过逆向反应解离成EB+A,所以加入标记A*后仍可与A交换,形成一定量的EA*B,从而使产物P带标记。标记P*的出现可与顺序机制相区别。
上述两种机制中A*→P*都需要B (及Q)的存在。乒乓机制则不然。没有B和Q的存在仍能见到A*→P*,这是乒乓机制与其他两者的区别。
根据可逆反应化学平衡的原理还可导出上述不同情况下酶反应的速度方程,帮助进一步了解酶反应动力学。
- 1.收购办法是什么意思
- 1.收购政策是什么意思
- 1.收购调拨时期 (1949—1979年)是什么意思
- 1.改善了农业生产基础设施是什么意思
- 1.改良区划是什么意思
- 1.改造沙荒地是什么意思
- 1.改造私商是什么意思
- 1.改革开放前的村级机构是什么意思
- 1. 改革教学方法是什么意思
- 1.改革统一经营的体制实行户有户养是什么意思
- 1.改革耕作制度是什么意思
- 1.政党种类是什么意思
- 1.政区分布是什么意思
- 1.政府的大力支持保证了农村邮电通讯业的发展是什么意思
- 1.政治学是什么意思
- (1)政治工作是什么意思
- 1.政治思想工作是什么意思
- 1.政法机关的建立与加强是什么意思
- 1.政社合一,实行公社所有制是什么意思
- 1.政策因素是什么意思
- 1.政策措施是什么意思
- (1)教具是什么意思
- (1)教堂是什么意思
- 1.教学是什么意思
- (1)教学人员是什么意思
- (1)散文是什么意思
- 1.敬老院发展步伐加快,集中供养人数增加是什么意思
- (1)数学是什么意思
- 1.数学是什么意思
- 1.数量是什么意思
- 1.整体推进,重点突破是什么意思
- 1.整体计算比较是什么意思
- 1.整体计算比较。是什么意思
- 1.整半劳动力的组合状况是什么意思
- (1)整经与穿经是什么意思
- 1.整顿、恢复和稳步发展阶段是什么意思
- 1.整顿管理制度是什么意思
- 1.整顿队伍是什么意思
- 1.文义是什么意思
- (1)文件、函电等是什么意思
- 1.文化是什么意思
- 1.文化_(1)文化学是什么意思
- 1.文化馆是什么意思
- 1.文化(艺术)馆、站是什么意思
- (1)文字种类是什么意思
- (1)文学作品是什么意思
- (1)文学批评是什么意思
- 1.文学种类是什么意思
- 1.文物古迹是什么意思
- 1.文艺创作是什么意思
- 1.文艺学是什么意思
- 1.新中国成立初期是什么意思
- 1.新中国成立初期的法制宣传工作是什么意思
- 1.新中国成立初期缓慢增长是什么意思
- 1.新中国成立前是什么意思
- 1.新中国成立前中等农业教育发展概况是什么意思
- 1.新中国成立前农村文化状况是什么意思
- 1.新中国成立前甘肃高等农业教育发展情况是什么意思
- 1. 新乡市是什么意思
- (1)新技术、新产品开发是什么意思