物质代谢的调控
生物体的整体,或其器官组织和细胞,都必须有适宜的环境,方能使生命所需的物质代谢正常进行。一旦外环境有所改变,体内的内环境也必定有所反映,随之物质代谢就会有相应的调节控制,使代谢途径正确无误,速度适宜,营养物质的供应和代谢产物的应用及消除得当,才能维持生命的正常继续。物质代谢主要是在细胞内进行,而外界的改变首先影响生物整体,随之而来的则是内环境的改变,这就使细胞受到影响,致使物质代谢中的化学反应必须有所调节。因此,物质代谢的调控是在生物体的整体、细胞及分子水平上起作用的。
物质代谢在整体水平的调控 一个生物体与其所生活的环境有着密切关系,其体内的代谢必定因环境的不同而异,尤其受营养素的供应影响,更为明显。爱斯基摩人生活于寒冷北极地带,主要只能食用兽、鱼等动物食品,其能量来源自然是脂肪及蛋白质,因此其体内的脂肪及蛋白质的分解代谢必然旺盛。而居住于较温和区域的人群,以含淀粉多的谷物为主食,体内能量的产生就多来自糖的分解代谢。即使生活在同一地区,因食性不同或供应短缺,往往会造成一种或几种营养素的不足,例如旧中国食米区因食用久磨精米而缺乏维生素B1。维生素B1以焦磷酸硫胺素的形式,是丙酮酸脱氢酶及α酮戊二酸脱氢酶的一种辅酶,故当维生素B1缺乏时,丙酮酸不能脱去羧基而进入三羧酸循环彻底氧化,因而堆集,以致引起脚气病的一些症状。再者,食性的改变也能影响促进化学反应的酶在量方面的增减; 例如婴幼儿以奶为最主要的食品,故其胃粘膜分泌凝乳酶较多,而不食用或少食用奶的成年人胃中这种酶则几近缺如,因此有些成年人在食用牛奶后,不能将其很好凝结而消化,以致腹泻。
在特殊情况下,如饥饿或应激,体内必须调节物质代谢,以应变故。某些病理状态可使病人不能进食,若不及时补充营养素,尤其是葡萄糖,当即造成体内一种饥饿状态。饥饿时,体内物质代谢必定发生变化,且随时间的延长而逐渐改变,这可从其血糖水平的变化看出一梗概(表1)。在饥饿期间,体外营养素的供应断绝,但体内不能无葡萄糖; 虽然脑组织可以适应或多或少的利用酮体,而红细胞所需的ATP则完全依靠血糖在细胞内的分解,所以其它器官组织中糖的分解代谢逐渐降低,而蛋白质的分解加强以增加糖异生作用; 同时三羧酸循环不能不运行以产生ATP,因此大量动用脂肪进行分解,使体内能量需求得到满足。这些物质代谢的改变都是由某些激素来调节的。现将饥饿中一些代谢参数列于表2。从表2可以看到血中胰岛素水平逐渐下降,胰高糖素有所提高; 还可以看到酮体明显增多,也可看到氨基酸水平下降。这无疑是脂肪分解及糖异生作用等增加的结果。
应激就是机体对外界异常刺激的适应。这样的刺激可包括创伤、某些外科手术、肾衰竭、烧伤、感染、受冻及强烈情绪激动等。出现任何一种这样的情况时,体内总的表现有血液中皮质醇、胰高糖素、儿茶酚胺及生长激素等升高; 胰岛素的分泌虽然不受影响,体内的抗胰岛素性明显存在; 结果脂肪分解增加,故许多组织内脂肪的氧化代替了葡萄糖的氧化,因而加速了酮体的生成,使体内能量代谢发生改变; 同时蛋白质代谢亦发生异常,总的看来,分解增加,合成减少,使绝大多数的氨基酸在肝内脱去氨基,将生糖氨基酸的碳骨架经糖异生途径生成糖,并将氨基合成为尿素,因此造成患者体内氮的负平衡,这可能与皮质醇在应激者体内水平大量升高有关。
表1 血糖在饥饿中的变化
| 饥饿持续时间(h) | 血糖中葡萄糖(g/24h) | |
| 转换率 | 氧 化 | |
| 7~14 38~74 120~792 | 152 120 87 | 96 70 46 |
表2 饥饿时物质代谢的一些参数
| 饥饿持续时间 | |||
| 过夜 | 3d | 28~42d | |
| 胰岛素(mU/L) 胰高血糖素(ng/L) 葡萄糖(mmol/L) 乙酰乙酸+β | 37 75 4.8 | 20 140 3.8 | 17 110 3.8 |
| 羟基丁酸(mmol/L) 丙氨酸(μmol/L) 亮氨酸(μmol/L) | 0.1 344 112 | 1.8 321 152 | 7.4 140 72 |
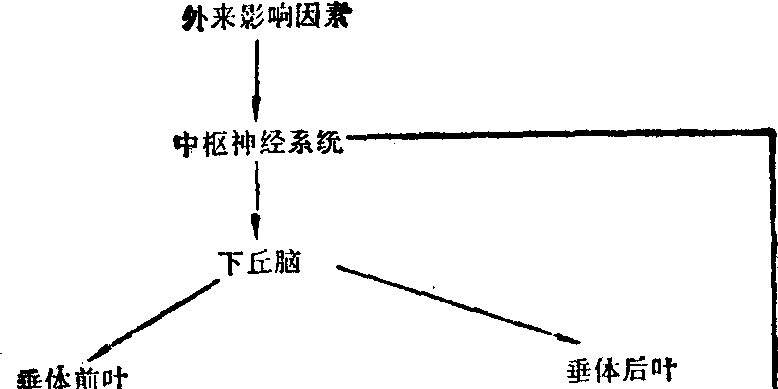
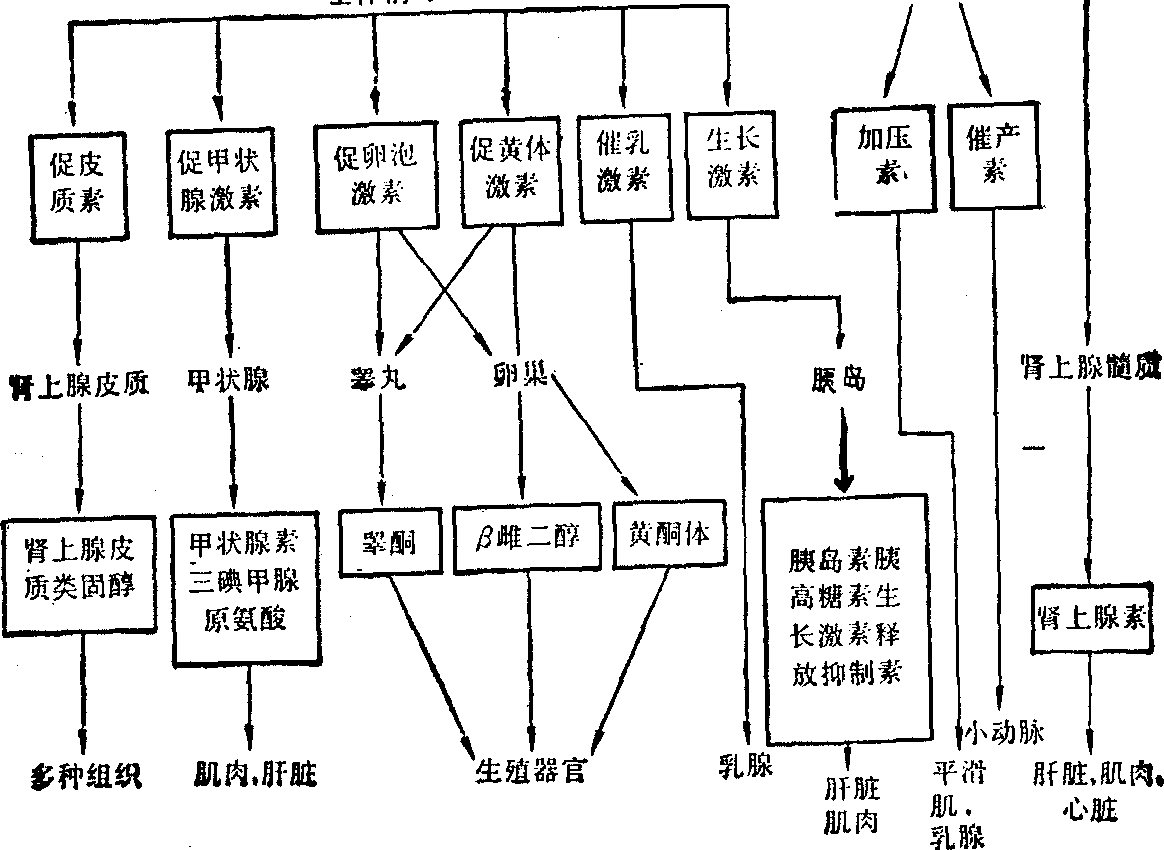
图1 调节体系示意图
纵上所述,可见机体总是通过神经体液来调节体内的物质代谢,以应付所遭遇的异常情况。
物质代谢在细胞水平的调节 (1) 作用于细胞的调节体系: 体内的物质代谢是受机体所在环境的影响。外来的刺激因素首先影响神经,然后传导到内分泌腺,以分泌激素,经血流而达到各种组织细胞并调节其物质代谢。激素是调节细胞中代谢的物质。各种激素虽然与全身的细胞都有接触,但只对其靶组织中的靶细胞起作用。现将这种调节体系中较重要者用图表示,其各环节间的关系如上图:
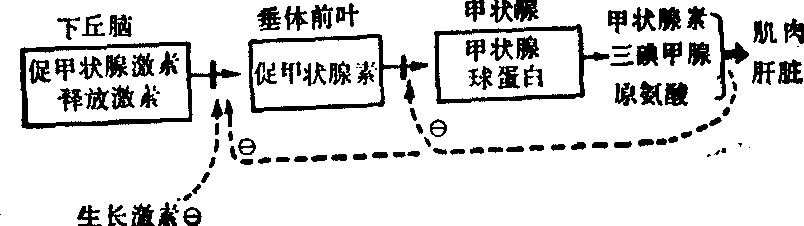
图2 反馈抑制示例Ө代表抑制
激素的调节也受反馈抑制的控制,例如下丘脑可分泌促甲状腺激素释放激素,促垂体前叶释放促甲状腺激素,以促进甲状腺合成甲状腺球蛋白,作为合成甲状腺素及可以抑制垂体前叶释放促甲状腺激素,也可以抑制下丘脑分泌促甲状腺激素释放激素。此外下丘脑及胰脏分泌的生长激素释放抑制素也抑制促甲状腺激素释放激素的分泌。这一反馈抑制的例子可图示如上页:
(2) 激素的受体: 激素具有高度特异性,只作用于靶细胞,这就意味着靶细胞必定拥有使激素能识别的特殊结构; 这种结构就是激素的受体。激素的受体有的是在细胞的表面上,也有的是在胞液中,在细胞表面者为水溶性的肽及胺类激素的受体,而存在于胞液者则是易于通过质膜的脂溶性类固醇类激素的受体。它们可能都是一些特定的糖蛋白,对于激素分子都具有高度的特异性及亲和力。一种激素能选择性地识别其特定的受体,并与其结合成复合物。这种复合物即会导致“第二信使”的生成或释放。第二信使将激素与受体的复合物所携带的信息传递给特定的酶或分子体系,使激素所负的使命得以实现,这就是细胞内的第二信使调节特定酶促反应或使特定基因能够表达。

(3) 环磷酸腺苷的作用: 在许多情况下,第二信使实际即是环3′,5′-磷酸腺苷,简写为cAMP,其生成过程可图示如下:
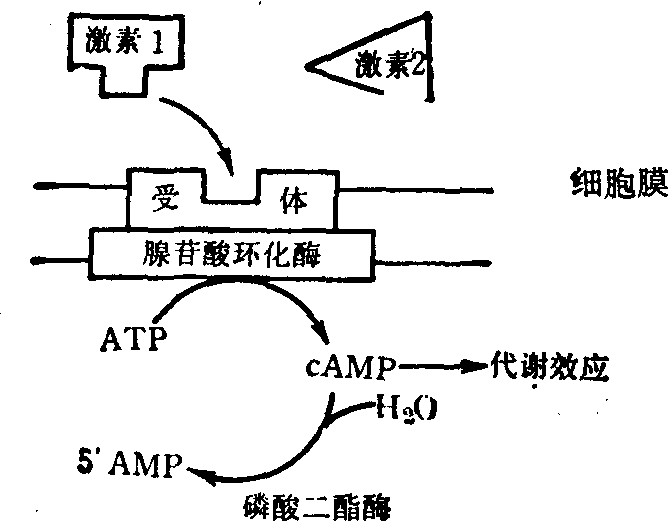
图3 cAMP的生成和水解
cAMP如何触发特定酶促反应的问题,可用糖原生成对糖原分解的作用来说明。在肌肉活动中肾上腺素与细胞膜上的受体结合而激发cAMP的生成;而cAMP当即触发糖原分解的级联反应(图4),同时抑制糖原的生成。当肌肉活动减少时,磷酸己糖因无需用以生产ATP而累积,即会反过来将糖原合成酶b激活为糖原合成酶a,以促进糖原的生成,同时抑制磷酸化酶b转变为磷酸化酶a。
cAMP也可影响胞核,其作用是通过激活特定蛋白质激酶,促使非组蛋白磷酸化而从与DNA的结合部位上脱落下来,从而使被阻遏的基因得以表达。

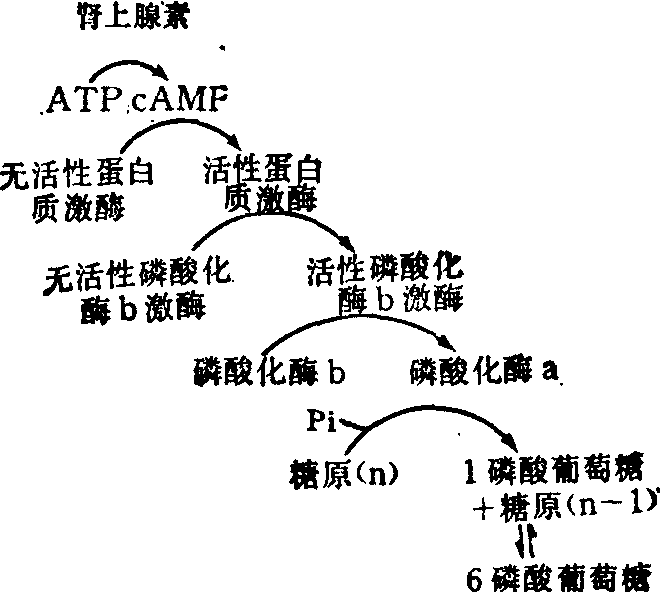
图4 糖原分解的级联反应
(4)性激素对靶细胞的作用: 雌激素及雄激素均为脂溶性类固醇物质,能透过质膜而进入细胞; 其受体存在于胞液中,且不以cAMP为第二信使。例如,雌二醇在进入子宫及乳腺的靶细胞后,在胞液中与一特定4s的蛋白质(即受体)结合,经转变成5s的化合物(即第二信使)后而进入胞核,并作用于染色体而促进mRNA的生成,以合成特定的蛋白质。这一过程可用雌二醇对鸡输卵管靶细胞的作用来说明:
睾酮也有类似的作用,但必须先转变成二氢睾酮后,才能与其特定蛋白质结合。
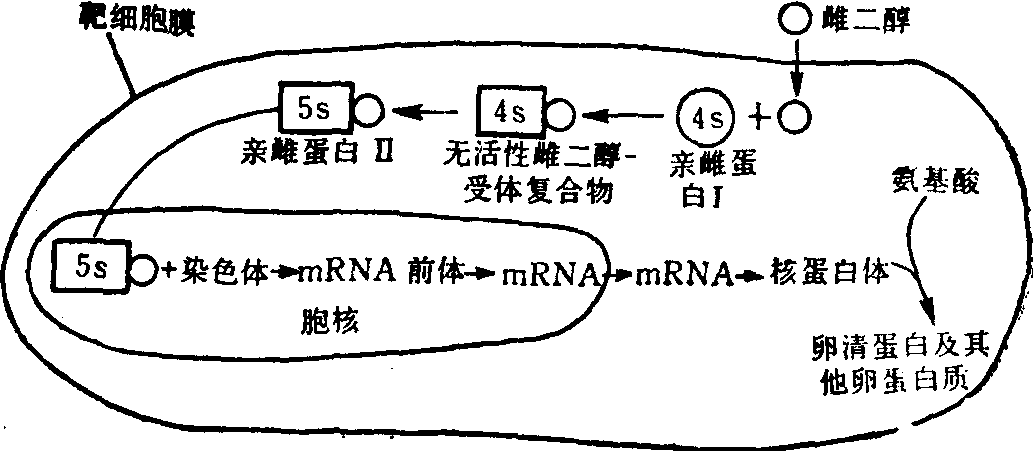
图5 雌二醇的作用机理示意图
亲雌蛋白I(estrophilin I)为雌激素受体;
亲雌蛋白II(estrophilin II)为活性雌二醇受体复合物
物质代谢在分子水平的调节 物质代谢中的各个化学反应主要由酶促成,所以酶分子的结构改变及合成、降解速度的增减乃是调节代谢最直接的因素。酶分子结构改变甚为快速,其发生以分秒计。而合成及降解则较为缓慢,需数小时才能实现。结构的改变包括分子的变构及修饰,与机体的需要及内环境有较大关系,至于合成及降解,尤其是合成则涉及基因的作用。
(1) 通过酶分子变构的调节: 一个代谢途径的发生或消失及进行速度的增减,取决于其调节酶的活性。而酶的活性,在温度、pH、作用物及辅助因子等恒定的情况下,又取决于其分子的结构。调节酶具有变构的性质,故常称为变构酶。变构酶往往位于代谢途径的开端处或其附近,其所促进的多为一重要而不可逆的反应,其分子结构可为一些激动或抑制效应分子所改变。变构酶的分子具有一些特定的结构部位,可与某些小分子结合,如与作用物、终末产物、代谢中间产物或甚至其它代谢途径的产物结合; 一经结合,酶分子的构象即会发生改变而影响其活性; 结合不同的分子,产生不同的结果。在分解代谢中,例如在糖酵解中,ATP是磷酸果糖激酶的作用物,也是其变构调节剂。当体内需要进行糖酵解以供给能量时,ATP与磷酸果糖激酶结合而促进酵解的进行,但在ATP水平升高到超过需要时,它又有抑制磷酸果糖激酶的作用,从而使酵解减慢或停止。柠檬酸盐也是磷酸果糖激酶的抑制剂,有降低酵解速度的作用。
变构酶可与多种小分子物质结合,但与代谢途径的终末产物结合是其特有的性质。当终末产物超过需要时,即与酶分子结合而改变其构象,因而抑制其活性; 这就是反馈抑制。变构反馈抑制的例子很多,其最早发现者为细菌中促进L苏氨酸转变为L异亮氨酸的酶体系。这一体系由五种酶组成,以苏氨酸脱水酶为这一转变开端的酶。苏氨酸脱水酶可被转变过程的终末产物,异亮氨酸所抑制; 这当然是异亮氨酸生成过多时发生。异亮氨酸是苏氨酸脱水酶的特异抑制剂,并不抑制体系中其它酶,同时其它的中间产物对苏氨酸脱水酶也无抑制作用。异亮氨酸与苏氨酸脱水酶以非共价键与酶分子形成可逆性结合,故在需要时异亮氨酸又可脱离结合部位,恢复酶的活性。由此可见,反馈抑制显然是通过酶分子的变构来调节代谢。
(2) 通过酶分子修饰的调节: 酶分子含有多种基团,有的可以去除或加上,也有的可以交互改变; 通过这种修饰作用以影响酶的活性,如加上或去掉磷酸及硫氢基与二硫键的互相转变等都是对酶分子修饰的常见例子。糖原的合成与分解就是通过糖原合成酶与糖原磷酸化酶的磷酸化与脱磷酸作用来调节的。糖原合成酶以I及D两种型式存在;I型为去掉磷酸并具有活性者,但经蛋白质激酶的磷酸化作用则成为无活性的D型。糖原分解中的磷酸化酶则以a及b两种形式存在,其本身的磷酸化或脱磷酸在活性上则与糖原合成酶的正好相反;磷酸化酶a是磷酸化型,在受磷酸化酶磷酸酶水解而脱去磷酸后,即成为无活性的b型。
(3) 通过基因的调节: 体内的酶与其它物质一样,也有其合成及分解代谢。通过酶蛋白的代谢,即可影响细胞内酶的含量,进而调节物质代谢的速度及存在。酶的半寿期甚短,如无合成,细胞内酶的含量即将逐渐下降。影响酶合成的因素有作用物、代谢产物、激素及药物等。这些因素都是通过基因来调节酶的合成。作用物影响酶合成的例子很多,例如食用低蛋白质高糖类膳食的人,其肝脏中分解氨基酸的酶的含量必定很低,因体内对无多大用处的酶,不必大量合成。一旦改食高蛋白质膳食时,一日之内肝脏中分解氨基酸的酶的含量必定增高。这表明,肝细胞内酶的合成当视膳食所供给营养素的性质而定,实际是作用物作用于基因而使所需要的酶得以合成; 一般称这种作用为酶的诱导,可图示如下:
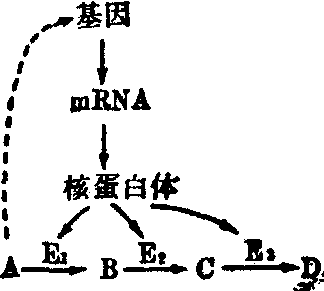
基因因作用物A的影响而生成A的代谢途径中所需要的酶E1、E2及E3等,将A依次转变为B、C及D。酶的诱导生成; 在细菌中亦是常见的,例如在含葡萄糖的培养基中生长的大肠杆菌不能使乳糖发酵,而在含乳糖或甲基半乳糖的培养基中生长者则能; 这是因为乳糖或甲基半乳糖作用于基因而诱导生成β半乳糖苷酶的结果。
代谢产物通过基因对酶蛋白合成的调节,最常见的是反馈抑制。当一代谢途径的终末产物超过需要时,即与调节酶结合而造成反馈抑制,致使这一代谢途径中酶的合成减少。
高等动物体内影响蛋白质合成的激素将信息依次传递给细胞的受体、第二信使及基因,然后由基因调节蛋白质的合成。例如,胰岛素对蛋白质有促进合成并抑制分解的作用,这就极有可能使糖酵解及脂肪酸合成等代谢途径中的酶在细胞内的含量有所增加。又如糖皮质激素促进肌肉蛋白质的分解,由此产生的氨基酸在肝细胞中通过糖异生作用而生成糖,所以这个激素的作用是导致氨基酸分解代谢中及糖异生作用中酶的合成。
绝大多数药物及毒物都是异生物质,均能诱导生成分解它们的酶,以解除其不利于生命的作用。药物及毒物多在肝中进行生物转化; 促进生物转化的酶,常称为微粒体酶,都不是潜在的或已有酶前身物的激活,而是经异生物质的诱导而从新合成的。这无疑是通过基因的一种调节作用。就现在所知,药物及毒物诱导剂大致可分为苯巴比妥及多环烃两种类型,前者所能激起微粒体酶的种类远远大于后者。
纵观上述,物质代谢的调节虽然分为整体、细胞及分子三种水平来讨论,但实际上体内代谢的调节总是依次通过神经、激素及基因,最后落实到酶分子上来实现。
- 想从今、无暇斸蔷薇,锄罂粟。是什么意思
- 想从前我们俩分手是什么意思
- 想他好晴是什么意思
- 想他的帐是什么意思
- 想会到是什么意思
- 想会开是什么意思
- 想似是什么意思
- 想住是什么意思
- 想作是什么意思
- 想你想的肝肠断是什么意思
- 想你等6则是什么意思
- 想佳人、妆楼凝望,误几回、天际识归舟.是什么意思
- 想佳人、妆楼颙望,误几回、天际识归舟。是什么意思
- 想保暖住雪屋是什么意思
- 想倒是什么意思
- 想倒眠了是什么意思
- 想偏了心是什么意思
- 想偏心是什么意思
- 想做是什么意思
- 想做什么就做什么是什么意思
- 想做某件事的兴趣是什么意思
- 想做某件事的念头是什么意思
- 想停止而停不下来是什么意思
- 想偷是什么意思
- 想偷汉子又害羞是什么意思
- 想像是什么意思
- 想像、说话毫无拘束,漫无边际是什么意思
- 想像不断涌现是什么意思
- 想像中国的方法:历史·小说·叙事是什么意思
- 想像催眠是什么意思
- 想像力是什么意思
- 想像消魂,怨东风、孤衾独晓。是什么意思
- 想像若在眼,周流空复情。是什么意思
- 想像虬龙去,依稀羽客随。是什么意思
- 想像训练是什么意思
- 想像飞鸣态是什么意思
- 想像飞鸣态,如闻霄汉音是什么意思
- 想儿是什么意思
- 想充人物是什么意思
- 想充小白人儿是什么意思
- 想入非非是什么意思
- 想入非非;异想天开是什么意思
- 想入飞飞是什么意思
- 想八路是什么意思
- 想兹为襁褓,如鸟拾柴枝。是什么意思
- 想冤家想得魂飘荡是什么意思
- 想净反落脏是什么意思
- 想出了神是什么意思
- 想出办法是什么意思
- 想出挖出是什么意思
- 想出欠帐是什么意思
- 想出花头是什么意思
- 想分香旧恨,刘郎去後,一溪流水。是什么意思
- 想到是什么意思
- 想到“血洗”是什么意思
- 想到一块了是什么意思
- 想到一块儿是什么意思
- 想到一块儿了是什么意思
- 想到一点子上是什么意思
- 想到与得到是什么意思