有丝分裂mitosis
出现染色体和纺锤体的细胞分裂形式。亦称间接分裂。主要表现在细胞核发生一系列可见的形态学变化,故又称核分裂。是真核生物体细胞增殖的基本形式。有丝分裂时细胞停止执行其正常的生理功能。由于在有丝分裂中经过复制的DNA平均分配到两个子细胞内,使遗传物质得以在细胞世代间稳定相传。
过程 习惯上人为地分为前期、前中期、中期、后期和末期。
前期 从分裂开始到核膜消失为止。开始时,细胞停止功能活动,变为圆形,核略膨大。在此期间主要发生染色体形成、分裂极确立、核仁解体和核膜消失等过程。
染色体形成 在染色体凝聚因子作用下,已复制的DNA双链进一步螺旋化形成染色丝,随后缩短变粗,形成一定数目和形状的染色体。每条染色体由一对紧靠在一起的姐妹染色单体组成,两者于着丝粒处相连。
分裂极的确立 动物细胞分裂极的确立与中心粒的活动有关。在细胞分裂间期,已复制的两对中心粒开始向相对的两极移动,并分别向其周围发出星射线;在两对中心粒之间靠近核膜处开始形成纺锤体微管;随着核膜破裂,两对中心粒移向细胞两极,纺锤体最终即在此形成。高等植物细胞没有中心粒,但也能形成纺锤体,分裂极即由纺锤体形成的方向确定。
核仁解体 核仁逐渐变小并解体,构成核仁的部分物质转移到染色体上,以后被分配到子细胞中,再参加新核仁的形成。
核膜消失 核膜解体成小泡分散于细胞质中,但在低等真核细胞生物中则可见到分裂过程中核膜不破坏的现象。
前中期 主要过程是形成纺锤体和染色体排列在赤道面上。
纺锤体形成 纺锤体与染色体运动密切相关,没有纺锤体,染色体就不能排列于赤道面上,两条染色单体也不能移向两极。纺锤体微管组成4种丝状结构:❶连续丝(极间丝)是连系两极间的纺锤丝;
❷染色体丝(着丝点丝或牵引丝)是着丝点与一个极相连系的纺锤丝;
❸中间丝是细胞分裂后期出现在两组子染色体之间的纺锤丝,不与两极或着丝点相连;
❹星体丝是从两极的中心体放射出的丝状结构,只存在于动物和低等植物细胞内。
染色体排列于赤道面 牵引力平衡假说认为染色体是依靠两极牵引力的平衡排列在赤道面上。
中期 从染色体排列于赤道面上到两条染色单体开始移向两极的过程。有时也把前中期包含在中期内。此时染色体高度聚缩,形态和数目均能分辨清楚,若用秋水仙素处理,纺锤体微管解聚,细胞分裂停止,此时最适于进行染色体组型等的研究。在此时期,若从细胞一极观察,染色体着丝点朝向中央,而染色体臂呈辐射状朝向周围。由星体、纺锤体和染色体共同组成的复合装置称有丝分裂器,此时已完全形成。
后期 每对姐妹染色单体分开并移向两极的过程。染色单体通常是先从着丝粒处分开,随后两个染色单体的臂逐渐分离,当它们完全分开形成两个子染色体后,便分别向两极移动,结果在细胞的两极各有一组数目完全相等的染色体。染色体移向两极可能是由于牵引丝缩短和连续丝延长的结果。
末期 从子染色体到达两极至两个新细胞形成的过程。染色体到达两极后,首先发生膨胀、解旋而轮廓消失,形成大块的染色质; 在染色质周围由内质网和原先崩解的核膜小泡重新组成新的核膜; 核仁相继出现,子核即告形成。动物细胞的胞质分裂是以缢缩和起沟方式完成的,一般认为缢缩是由于细胞质周缘有一微丝组成的“收缩环”紧缩引起的,微管参与起沟的过程。
有丝分裂过程中,某些主要细胞器如线粒体、叶绿体、鞭毛与纤毛等都能进行自我复制。细胞质代谢也发生明显变化,如分裂前期,呼吸作用开始明显下降,到中期达到最低点,后期又逐渐回升。核酸合成作用在分裂期间几乎全部停止,蛋白质合成也降到最低水平。
有丝分裂过程中各期持续的时间虽然在不同物种有所差别,但一般是前期最长,末期次之,中期和后期都很短,前期历时约比中期、后期长4~6倍 (图1、2)。
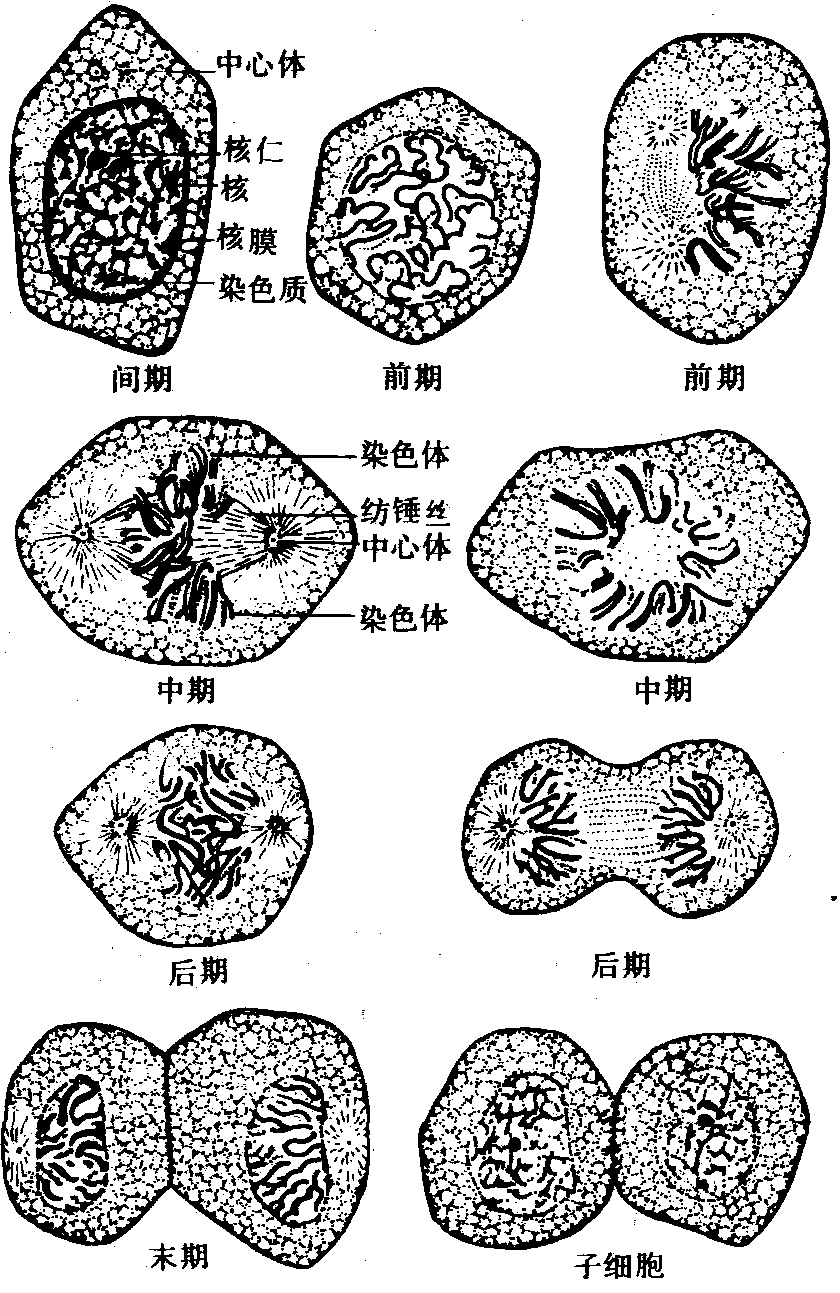
图 1 动物细胞有丝分裂模式图
动植物细胞有丝分裂的区别 所有真核细胞的有丝分裂过程基本相似,但在动物和植物之间、高等植物与低等植物之间都存在某些差异,高等植物细胞因无中心体,故缺乏星体。多数植物靠染色体丝的缩短使子染色体向两极移动。植物细胞的胞质分裂开始于早末期,起始赤道面处的微管数量增加,并向周围扩展形成桶状的成膜体。与此同时,来源于高尔基体和内质网的一些小泡和颗粒成分沿赤道面排列、融合而成细胞板。新形成的细胞板两侧继续积累多糖,发育成细胞壁。
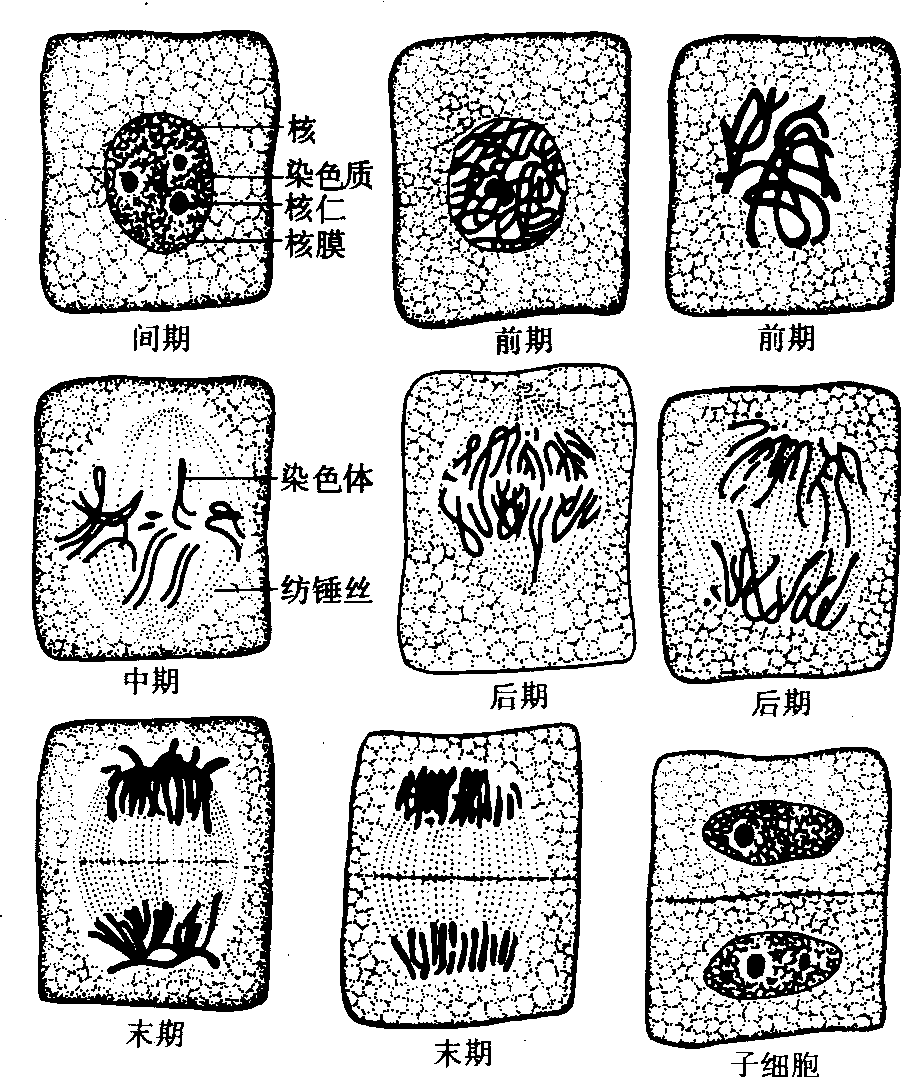
图 2 植物细胞有丝分裂模式图
异常的有丝分裂 正常而完整的有丝分裂应具有DNA复制、染色线复制、染色单体复制、染色体复制、核复制和细胞复制的一系列顺序进程。如果其中一个或多个程序受到抑制,将导致细胞核、染色体组成和染色体形态等出现种种变异。如果胞质分裂过程被抑制,将形成双核细胞,当多次受到抑制时,则形成多核细胞,如动物的横纹肌纤维和小鼠的肝细胞、高等植物的绒毡层细胞等。当核复制和细胞复制均受到抑制时,有丝分裂前期、中期和后期都在核膜内进行,即核内有丝分裂,其结果造成核的多倍性,如某些昆虫细胞和花药绒毡层细胞。有丝分裂中核物质的减少除减数分裂外,也存在体细胞的减数分裂过程,国内学者在高等植物中证明体细胞减数分裂后形成的子细胞是单倍性的。体细胞减数分裂有二种类型: 一是染色体数目减半,在蚊的上皮细胞和蚊培养细胞中均可见到; 另一类型是染色体数目没有变化,但染色体的大小减半,此现象发生在具有多线染色体的细胞内。
影响细胞有丝分裂的因素 某些激素、酶类、环磷腺苷、多种化学药品、X射线、紫外线、温度等对细胞都有一定的调控作用。如秋水仙素能使微管解聚或阻止微管的聚合过程,因而能抑制纺锤体的形成。以致染色体不能移向两极,细胞分裂停止于中期,或组成一个双倍性的核。
有丝分裂mitosis
又称间接分裂或核分裂。通过形成纺锤体,使复制后的染色体平均分配到每个子细胞的一种细胞分裂过程,是真核细胞繁殖的一种基本方式。这种分裂是一连续的复杂的动态过程,包括核、染色体和纺锤体、细胞器的一系列变化以及染色体平均分配到每个子细胞,从而使复制好的遗传物质准确分开,得以在细胞间世代相传。通常将有丝分裂过程划分为前期、中期、后期和末期4个时期。在前期,染色体出现,分裂极确立,核仁解体,核被膜消失。在中期,染色体排列在赤道面上。在后期,染色体的两条姐妹染色单体分开并移向两极。在末期,子核形成,胞质分裂形成两个新细胞。
有丝分裂yousifenlie
真核细胞的一种最基本的分裂方式,其核分裂过程发生染色体的一系列动态变化,并伴有纺锤丝的出现,胞质分裂后遗传物质平均分配给两个子细胞。根据染色体的动态变化可划分为前期、前中期、中期、后期和末期五个分期。从染色质集缩成染色体到核膜解体为前期。此时期染色质螺旋化形成的每个染色体,含有一个着丝粒区域和纵向并列的两条染色单体。动物细胞在间期完成复制的两对中心粒分开,移向相对的两极,两对中心粒之间近核膜处开始形成纺锤体微管。植物细胞没有中心粒,纺锤体微管由近细胞核两极部位的细胞质中发出。前期的后半期核仁解体,核仁线分散在核液中,一些核仁物质粘附于染色体上。前期末核膜解体成小泡,分散于细胞质中。从核膜解体到染色体排列在赤道面上为前中期。核膜解体后,由两极发出的两组纺锤体微管进入核区,在赤道区互相接触形成侧支架桥,变形而成为纺锤体。某些极微管(即染色体丝)分别与每个染色体的一条染色单体的着丝点连接,在两侧染色体丝的均衡牵引下,全部染色体的着丝粒区整齐地排列在赤道面上,染色体臂则伸向赤道面的两侧。从染色体排列在赤道面上到子染色体开始向两极移动为中期。中期持续时间很短。中期染色体集缩成典型的形态结构,由于两极牵引力达到平衡状态,所以染色体也处于一种不断摆动的平衡状态。从子染色体开始向两极移动到移至两极为后期。此时期的两条染色单体的着丝粒彼此分开,染色体丝缩短产生的拉力以及连续丝延长产生的推力,共同使两组子染色体分别移动到达两极。从子染色体移至两极到形成两个子细胞为末期。移至两极的染色体解螺旋化复原成染色质,在每组染色体周围聚集的核膜成分融合成为核膜。随着子细胞核的形成,核内出现新的核仁,从而核分裂完成。在核分裂结束后,或者染色体解螺旋和形成核膜的同时进行胞质分裂,甚至有时在中后期就已经开始胞质分裂。动物和低等植物细胞的胞质分裂以缢缩和起沟方式进行,在空间分布上无拘束的细胞,其胞质的周边有一个由微丝组成的收缩环,它的紧缩使细胞产生缢束,最后在缢束处起沟而使细胞质一分为二。紧密排列的细胞靠逐渐起沟过程进行胞质分裂,即纺锤体赤道面周边的纺锤丝开始聚集稠密物质并逐渐布满整个赤道面而形成中体,中体与沟底细胞膜连接,使裂沟不断加深,而中体变得越来越小,直至胞质分裂完成前消失。植物细胞的胞质分裂靠细胞板的形成,此时期纺锤体两极的纺锤丝消失,中间区的纺锤丝向四周扩展形成桶状的成膜体,随后来自于内质网和高尔基体的小泡及颗粒等成分沿赤道面排列,融合而成细胞板,把胞质一分为二。然后在细胞板两侧积累多糖,发育成细胞壁。
总之,在间期进行一次复制的DNA,通过有丝分裂平均分配到两个子细胞中去,使子细胞与亲代细胞中遗传物质保持恒定,从而表现出与亲代相似的遗传性。
有丝分裂mitochysis
系细胞分裂中最常见的一种分裂方式。细胞增殖靠有丝分裂。一个细胞周期包括间期和有丝分裂期。有丝分裂期是一个连续的细胞变化过程。全过程共分为4期,即前期、中期、后期和末期。各期之间并无截然的界限,是由1个期逐渐过渡到下个期。此种分裂的特点主要是染色体呈一系列丝状分裂的过程,至染色体进入细胞的两极,核膜重新形成,新的细胞核完成为止。有丝分裂主要见于体细胞的分裂。
有丝分裂
细胞分裂方式之 一。即“体细胞分裂”。一般分4个时期:前期、中期、后期和末期。前期:中心体分裂并移向两极。染色体由于螺旋化而能被观察到,着丝粒以外都纵裂为二,成两个染色单体,最后核仁和核膜消失。中期:染色体移向纺锤体的中间,排列在赤道板上。后期:着丝粒分裂,染色单体在纺锤丝牵引下分别移向两极。末期:纺锤体消失,核膜和核仁出现,接着细胞质分裂。动物细胞通过分裂间隔的形成,植物细胞通过细胞板的形成,终于出现两个具有相同细胞核和大致上相等细胞质的子细胞。
有丝分裂mitosis
是最常见的一种细胞分裂方式。由于在分裂过程中有纺垂丝出现,故称有丝分裂。在分裂间期,染色体进行自我复制;在分裂过程中,复制后的染色体又均等地分配至两个子细胞中。有丝分裂的分裂间期较长,分裂期较短。分裂期又可分为前、中、后、末四个期。
有丝分裂
真核生物一般通过染色体的复制进行细胞分裂和增殖,这种分裂方式称有丝分裂,简称丝裂。每种生物细胞中都含有一定数目、形态的染色体,在有丝分裂过程中,通过染色体的复制和分离,把同样数目、同样形态的染色体分配到分裂后的两个子细胞中,使子细胞都具有完全和母细胞相同的染色体。
细胞从一次分裂结束到下一次分裂结束的周期称为细胞周期。细胞周期包括G1、S、G2和M四期:M为丝裂期;G1、S和G2期组成两次细胞分裂之间的间期。在间期发生DNA双螺旋的复制,而DNA为染色体的主要成分,因此DNA双螺旋的复制就是染色体复制的开始。
G1期(DNA合成前期) 此期中每条染色体只有一条DNA双螺旋,由两股DNA链联合组成,呈松散的细丝状。
S期(DNA合成期) 此期每条染色体中的DNA双螺旋分开成两股,每一股DNA链合成另一股互补DNA链,组成一个新的DNA双螺旋(半保留复制),这样就形成了两个DNA双螺旋(见“DNA复制”)。1957年Taylor用放射自显影技术予以证明。他先把细胞放在含有3H标记的胸腺嘧啶核苷的培养基中培养,细胞分裂一次,即染色体复制一次后,每条染色体中一股DNA链所合成的另一股互补DNA链都含有3H,因3H有放射性,能使覆盖于其上的胶膜中的银离子还原而显影,故全部染色体都有放射性。再把细胞放入不含3H标记的胸腺嘧啶核苷的培养基中培养,细胞第二次分裂,即染色体第二次复制后,这时每条染色体中的一股由一条含3H的DNA链和一条不含3H的DNA链合成互补链,因而仍有放射性;而另一股由不含3H的两条DNA链合成一条DNA互补链,因而没有放射性。所以第二次复制后只有一半的染色体有放射性,从而证明DNA的半保留复制(图1)。
G2期(DNA合成后期) 此期中两条DNA双螺旋已分别和组蛋白结合,形成两条染色单体,借着丝粒连接在一起。这样从G1期的一条染色体复制成两条染色单体,也称为姊妹染色单体。1973年,Latt在含有5溴脱氧尿嘧啶核苷(BUdR)的培养基中培养细胞,由于BUdR渗入染色体DNA以后可降低其螺旋化程度,当用Giemsa法染色以后,与未渗入BUdR者相比,染色较浅。当经过两次分裂以后,一条染色体的两条姊妹染色单体中,一条含有双股BUdR的DNA,另一条含有单股BUdR的DNA,前者较后者着色浅。所以也直接证明了DNA的半保留
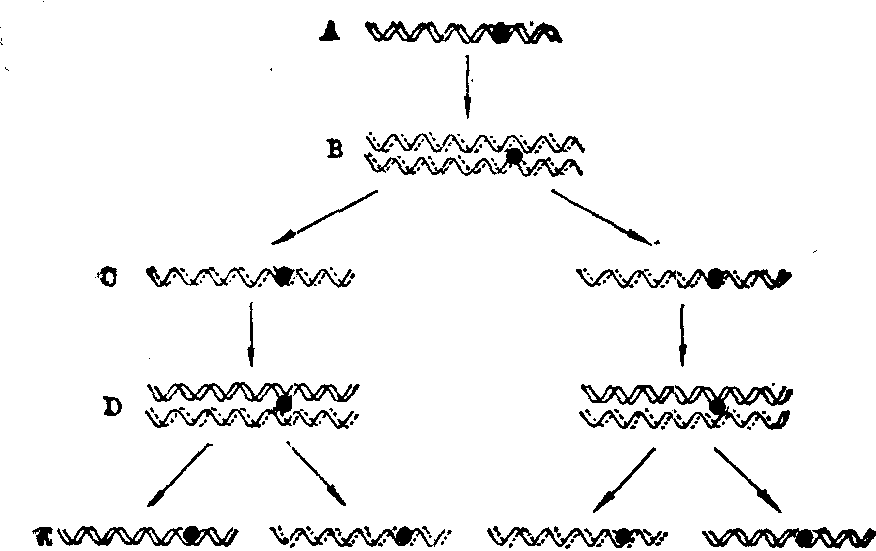
图1 DNA的半保留复制
A,B. 在含有3H标记的胸腺嘧啶核苷培养基中培养。其所复制的染色体中的互补DNA链均含有3H,所以全部染色体都有放射性
C,D,E. 在不含
 H标记的胸腺嘧啶核苷培养基中培养。其所复制的染色体中的互补DNA链均无放射性。因此只有一半的染色体有放射性
H标记的胸腺嘧啶核苷培养基中培养。其所复制的染色体中的互补DNA链均无放射性。因此只有一半的染色体有放射性A为染色体的两股DNA链:B为复制的中期染色体,虚线示含3H的互补DNA链; C为分裂后期的染色体;D为复制后的中期染色体;E为分裂后期的染色体
复制。实验结果又表明,某些姊妹染色单体深浅相间,说明姊妹染色单体之间发生了交换,这种现象就叫姊妹染色单体交换。

图2 姊妹染色单体交换
经特殊处理后,两条姊妹染色单体染色深浅不同,从各条染色单体的深染和浅染节段的排列,可证明某些姊妹染色单体的一些节段之间曾发生交换M期(丝裂期) 根据此期染色体的形态变化特点,又可分为四期:
❶前期:早前期的每一条染色体实际上是由两条染色单体密切结合而组成的。染色体开始螺旋化,因而逐渐缩短,成为光学显微镜可以看到的染色线,其上的染色粒清晰可见。随着染色体螺旋化程度增加,染色体更加缩短。在前期开始或更早时期,中心粒形成两个,并移到细胞核的两边,每个中心粒周围可见到辐射状的星射线,并在两个中心粒之间形成由微管构成的纺锤体。此时,核仁、核膜逐渐消失,染色体更缩短变粗,向赤道面移动。
❷中期: 染色体的着丝粒排列在纺锤体两极之间的赤道面上,形成赤道板。着丝粒各与一条由肌动球蛋白构成的牵引丝相连,牵引丝分别通向一极。姊妹染色单体开始分离。
❸后期:染色单体分开而成子染色体,着丝粒也已经分裂,由于牵引丝的作用,子染色体开始向细胞两极移动,着丝粒在前,两臂向后,呈“V”形“L”形或棒状。
❹末期:染色体到达细胞的两极后,逐渐解旋,最后舒展松散,不易分辨,在两团染色质周围,分别形成核膜,核仁重新出现,形成两个子核。细胞质接着分裂。在细胞表面,细胞膜沿赤道收缩,逐步加深,横缢而成为两个子细胞。
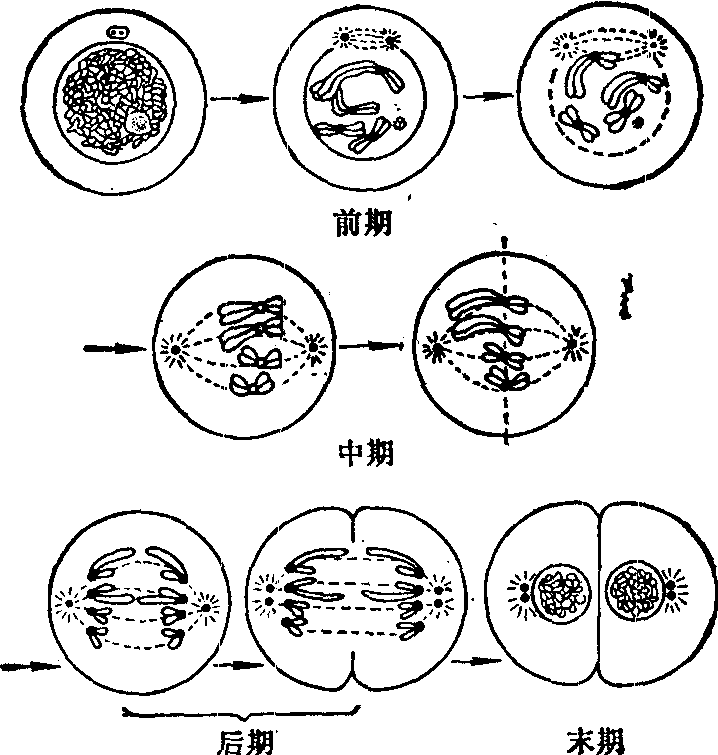
图3 细胞有丝分裂过程
经过有丝分裂形成的两个子细胞,含有与母细胞完全相同的染色体。多细胞生物个体的生命的开始都是一个合子——受精卵,经过许多次的细胞分裂,组织分化和整合,成长为一个个体,一个个体的全部细胞中的染色体在数目和形态上都是基本相同的,都同样含有该个体的全部遗传信息。所以有丝分裂过程保证了细胞核中每一条染色体和其全部遗传信息的准确复制和平均分配于子细胞。
有丝分裂
细胞分裂包括核分裂及胞质分裂两个过程,一般核分裂后随之发生胞质分裂。高等真核生物中核分裂主要是以有丝分裂方式进行,有丝分裂又称间接分裂。增殖细胞经过间期的G1、S、G2各期,重要大分子特别是载有遗传信息的DNA复制加倍,然后通过有丝分裂,精确地将其平均分配到两个子细胞中。有丝分裂保证了携带遗传信息的染色体一代代地以相同的数目传递下去,从而维持了遗传的稳定性。
有丝分裂是经过长期的生物进化过程而逐渐演变成的一种比较完善的分裂方式。原核生物是进行直接分裂的,如大肠杆菌(E.coli) 的DNA分子为简单的环状附着在细胞膜的一点上,DNA复制后,接触点的细胞膜生长,向内延伸形成隔膜,借此把两个DNA分子分配到两个子细胞中去。而真核细胞的DNA含量远比原核生物高。细胞在间期中,染色质分散在整个核区,适于DNA的复制和DNA的转录,但不利于遗传物质的均等分配。由于长期的演化产生了有丝分裂,在此过程中染色质浓集包装,成为数量一定的染色体,通过有丝分裂装置及一系列复杂、精巧的过程,把染色体平均分配到两个子细胞中去。
有丝分裂装置系指有丝分裂时产生的由微管及其结合蛋白所组成的星体和纺锤体。星体是指围绕中心粒向外辐射状发射的微管。纺锤体是由大量微管纵向排列组成的中部宽阔,两极收敛的细胞器,形状像纺锤,因而得名。纺锤体由连续牵丝(或称极间微管)、染色体牵丝(或称着丝点微管)及区间牵丝三类组成。连续牵丝由一极通向另一极,但绝大多数连续牵丝并非真正连续,而是来自两极的微管在赤道板处彼此相搭,侧面结合,有的是和两极均不接触的微管。染色体牵丝是指一端由极部发出,另一端结合到着丝点上的微管。区间牵丝是指在后期和末期时连接已经分向两极的染色体或子核的微管。有丝分裂装置在维持染色体的平衡、运动、分配中起着重要的作用,例如平均105次有丝分裂中只有一次产生误差。
有丝分裂过程 有丝分裂是一个核改组的连续过程,根据形态学特征人为地划分为前期、早中期、中期、后期和末期五个时期。
前期 标志前期开始的第一个特征是染色质不断浓集,实质上是染色质的螺旋化、折叠和包装过程。此时出现线状的纤维,有丝分裂因而得名。中前期时染色体纤维进一步缩短、变粗,已经能看到每条染色体是包含有两条染色单体的成双结构,并出现了主缢痕。主缢痕处染色体有一特化部位叫着丝粒,为重复序列的DNA,每一拷贝包含约300个碱基对,两条姐妹染色单体的DNA在此处互相传染,联系在一起,着丝粒本身不含有遗传信息。着丝点附着在主缢痕着丝粒上,过去认为主要是蛋白质性物质所组成,电镜下可见着丝点为板状或杯状的碟子,有三层结构,直径0.2—0.5μm。着丝点是染色体牵丝附着部位,牵丝可穿透各层和着丝粒相连。某些类群的植物、昆虫、藻类有弥漫性着丝粒,微管可沿染色体分布。着丝点是微管组织中心(MTOC)之一,体外实验证明,在适宜的条件下,微管蛋白可以着丝点为生发起点组装成微管,在体内情况下尚无明确的证据。
中心粒在G1期为一对互相垂直相邻的九组三联管构成的小圆柱体,直径0.25μm,长度不定,位于邻近核被膜的胞质中。在一对中心粒的周围是一团透明的、电子密度高的中心粒周围物质(PCM),这一复合物称之为中心体,实际上PCM具有MTOC的作用。一般在早或中S期时,两个中心粒稍有分离,以每个中心粒为模板,“复制”另一个中心粒(很可能PCM中蕴藏着重组中心粒的信息)。新复制的中心粒在S及M期逐渐延长。动物细胞在有丝分裂前期时,两对中心体连同其组装的星体沿核被膜呈弧线向细胞的两端移动。达到相对位置时决定了细胞分裂极。两极之间在靠近核被膜处形成初步的纺锤体。纺锤体的组装与去组装有一定的极性,极部为负极,此端微管蛋白亚单位的去组装大于组装,而另一端为正极,组装大于去组装。中心体体外组装纺锤体的实验证明,MTOC的功能在间期弱,中期很强,说明它有一个成熟过程。细胞进入前期时,由中心体所组装的布满细胞质的间期微管消失,代之以星体。
前期末时核仁消失,核被膜开始崩解。植物细胞没有星体,由许多微管组织纺锤体。
早中期 核被膜一旦崩解,则细胞进入早中期,此时核周围的纺锤体侵入细胞的中心区。一部分纺锤体微管的自由端最终结合到着丝点上,形成染色体丝。前中期的特征是染色体剧烈地活动,个别染色体激烈地旋转、振荡、往返于两极之间;纺锤体微管试图“捕获”染色体,并将其排列在中期板上。最终,一侧纺锤体牵丝的自由端“捕获”住一条染色体的一侧着丝点,接着另一侧的纺锤体牵丝的自由端捕获了该染色体的另一侧着丝点,这一过程是随机的。最后两侧相反方向力量的平衡使染色体排列在中期板上。前中期在哺乳类细胞中持续10—20分钟。
中期 染色体排列在赤道板上,细胞即进入中期,这时染色体牵丝作用于染色体上的力量持平。若用微束激光打断一侧染色体牵丝,则染色体被拉到相反极的一侧。纵向看,动物细胞染色体呈辐射状排列,植物细胞染色体则占据整个赤道面,小的染色体排在内,大的排在外缘。组成纺锤体的微管数目不等,酵母细胞可少到16条,高等植物可达5 000条,培养细胞上每一着丝点上可连34±5条微管。
后期 排列在中期板上的染色体,其姐妹染色单体借着丝粒联系在一起。后期伊始,几乎所有的姐妹染色单体同时分裂开,此时每条染色单体称之为染色体。此种分裂动力并非来自与两极相连的染色体牵丝的张力,因为在用秋水仙素处理破坏微管的情况下,两个单体也可以分裂开。着丝粒部位的分离,立即打破力的平衡,着丝点受到不断缩短的染色体牵丝的牵引而产生染色体的向极移动。着丝点在前,两臂拖后,视着丝点在染色体上的位置不同,呈v、j或i型。染色体向极运动的速度很慢,约为每分钟0.2—0.5μm。向极运动开始之后,接着便借助于极间牵丝的延长及滑动,把两极之间的距离推向更远。染色体的向极运动称之为后期A,两极间的距离增加称之为后期B。不同生物类型后期A和B所起作用不同,有的种类后期时两极间的距离全然未变,有的可达中期时两极间距离的15倍。
末期 后期结束时两组染色体完全分到两个子核中,核被膜片段重新包围两组染色体,组成完整的核被膜,形成两个间期核。同时染色体开始解螺旋,染色质分散于核中,核仁重新出现。在后期及末期时,两组染色体或子核之间有区间牵丝的形成。
胞质分裂 在动物细胞方面,虽然核分裂和胞质分裂经常是相继发生的,但是属于两个分离的过程,例如大多数昆虫卵,核可进行多次分裂而无胞质分裂,形成多核原生质团。某些菌类和藻类也是如此,多核细胞可长达数尺,以后胞质才分裂形成单核细胞。由中、晚后期开始,在细胞中部赤道板处胞质向下起沟,形成环状缢缩。虽然在此期中纺锤体逐渐瓦解消失,但在细胞中部微管反而增加,其中掺杂有浓密物质和囊状物,这一结构称为中体。在分裂末期,用微管抗体的间接免疫荧光法可以显示这一结构。缢缩逐渐加深直达中体,此时中体像一条系带联系两个子细胞。胞质分裂的机制被认为和肌肉收缩机制类似,为肌动蛋白和肌球蛋白分子的相互滑行所致。现已证明在细胞起沟处质膜下方有肌动蛋白与肌球蛋白成分的收缩环,后期时环已形成备用,此时若用电刺激可引起收缩。用肌球蛋白及肌动蛋白抗体处理,可阻断收缩。在分裂的海胆卵中已分离出具有ATP酶活性的肌动蛋白,说明胞质分裂和ATP分解供能有关。用抑制肌动蛋白的细胞松弛素B处理可抑制胞质分裂。细胞分裂也是一个细胞膜面积扩增的过程,后期细胞膜下贮存有大量的泡状细胞膜备用。用缩时显微电影拍摄活细胞,可见细胞在分裂时有激烈的发泡现象,和细胞膜的扩增有关(图1)。
植物细胞的胞质分裂和动物细胞不同。后期在纺锤体中央区域出现“成膜体”,在某些方面类似于动物细胞的中体。“成膜体”的形成首先始于赤道板处的细胞膜附近,电镜下可见此处分布有微管、电子密度物质和囊状物。附近的高尔基体逐渐向其间迁移是囊状物的来源。由于不断加入微管和囊状物,成膜体扩展到整个赤道板,其中的小囊也逐渐扩大,互相融合,最后形成一片连续的细胞膜。此时,成膜体遂转变成“细胞板”,细胞一分为二。“细胞板”最后导致细胞壁的产生(图2)。
核被膜的崩解与重组 核被膜上分布有核孔复合物,在核内膜之下有一薄层网状结构,称之为核纤层。目前已知哺乳动物细胞核纤层主要含有三种核纤层蛋白,分别称之为核纤层蛋白A、核纤层蛋白B和核纤层蛋白C,属中等纤维,混合组装成核纤层。核纤层的外侧和细胞膜、核孔复合物相结合,内侧和染色质上的一定位点相结合。此种相互作用可能对于在有丝分裂中维持和稳定染色质的高级结构有重要意义。细胞在间期生长过程中,核被膜面积不断扩增,故核纤层也相应地生长重组。核纤层蛋白的合成在整个间期中均在进行,这些亚单位或者直接加入到已有的纤维中间或端部,或者组成新的纤维。在核被膜崩解时,首先发生的是核纤层蛋白磷酸化,然后核纤层解聚,接着便是核被膜崩解,形成小囊泡,多残留于纺锤体的周围。一些核孔复合物则附着在染色体上。核纤层蛋白A、C分散于核质和胞质的混合溶胶中,而核纤层蛋白B和核被膜囊泡紧密结合。分裂末期,首先三种核纤层蛋白去磷酸化,重新聚合成核纤层,带有核纤层蛋白B的核被膜囊泡,在核纤层蛋白聚合的同时来到核的周围,最终联成完整的新核被膜,核孔复合物重新返回到核被膜上,因而核纤层蛋白的磷酸化与去磷酸化调控核被膜的崩解与重组。
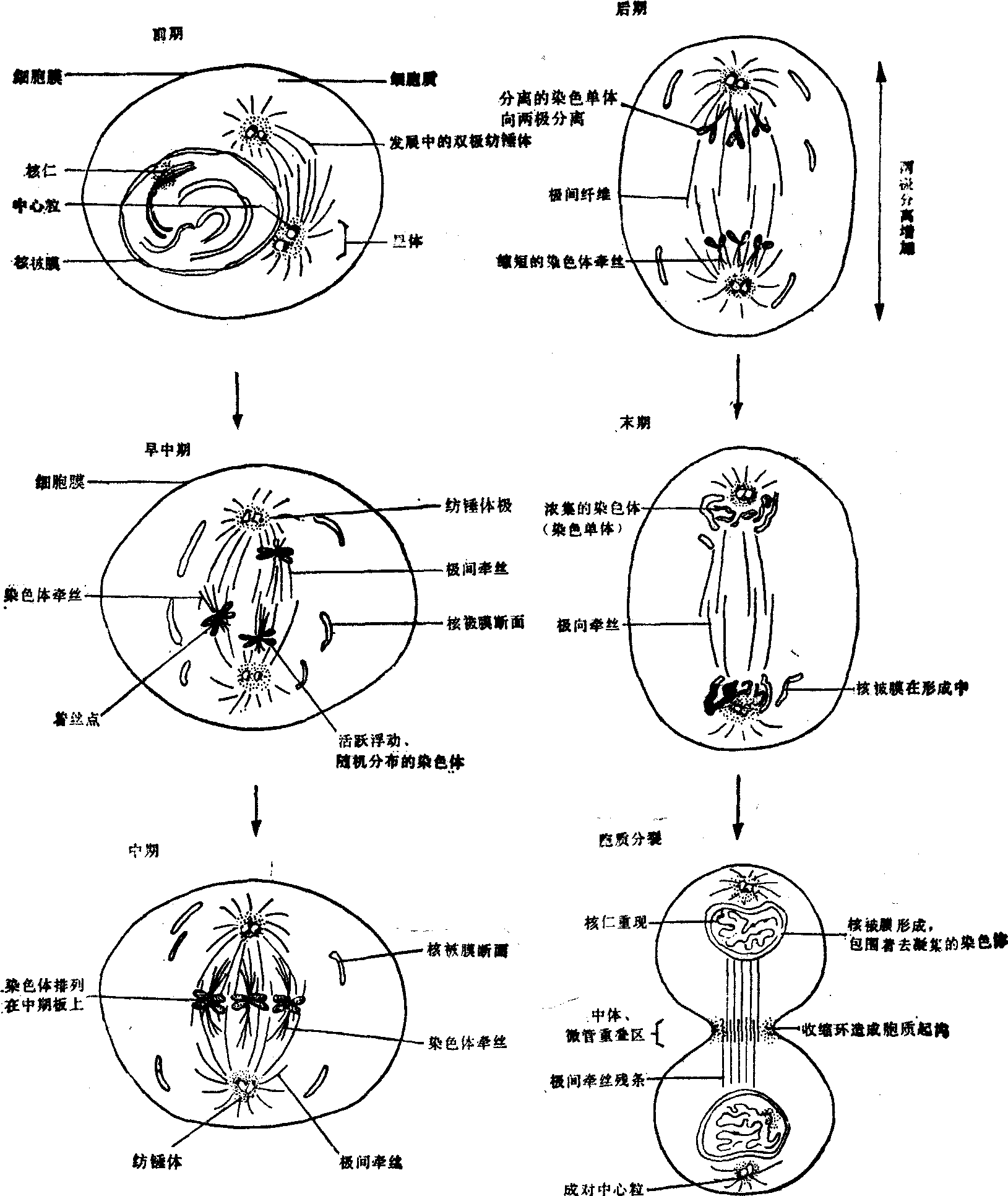
图1 有丝分裂过程
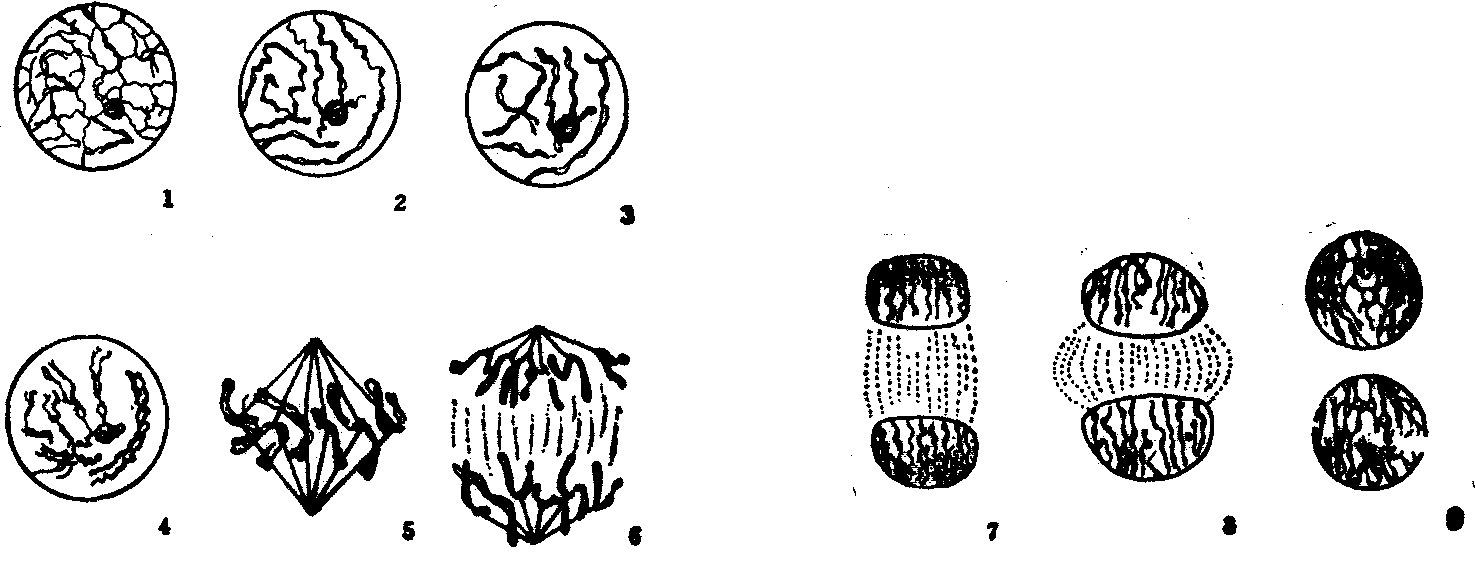
图2 植物有丝分裂各时期
1.前期开始; 2—4.前期; 5. 中期; 6.后期; 7—8.末期; 9.末期结束
着丝粒与着丝点 长期以来着丝粒与着丝点这两个术语是作为染色体上纺锤丝附着区域的同义语使用的。遗传学文献中多用着丝粒一词,细胞学家多用着丝点一词。后来在电镜下研究哺乳类染色体超微结构时发现,主缢痕两侧为一对三层结构的特化部位,认为是非染色质性物质的附加物,将此种结构,或最外一层称之为着丝点。将主缢痕内高重复序列的异染色质,易为碱性染料深染的区域称为着丝粒。但并非有丝分裂的各期,或各种生物的染色体均有此种分化的结构,如鸟类的大染色体上有此种碟状三层结构,而小染色体上却没有。
由于从硬皮病CREST综合征病人血清中发现了抗着丝点抗体,用间接免疫荧光法可以显示在中期染色体的主缢痕部位有一对亮的荧光斑点,即为着丝点。间期细胞的着丝点称为前着丝点,在G1期细胞核中为单个荧光斑点,S期逐渐出现成对的荧光点,到G2期则完全是成双的荧光斑点,从而证明着丝点在S期复制。
染色体的运动机制 关于染色体在细胞分裂中运动的机制有多种学说,多主张和有丝分裂器的活动密切相关。其中主要有动力平衡学说及滑动微管学说。
(1)动力平衡学说: 1967年有人用偏振光显微镜观察活细胞后,得出结论:后期染色体向两极移动是由于染色体牵丝在极部微管蛋白的亚单位的不断去组装,逐渐缩短,从而拉动染色体以每分钟1μm的速度向极移动。在偏振光显微镜下可见排列有序的微管出现双折光现象,后期时极部微管的双折光现象消失,说明着丝点微管极部的去组装所产生的力来拉动染色体。经计算,拉动一条染色体通过如此短的距离需力甚小,仅为10-13N,约需30个ATP分子。纺锤体上有ATP,极部含有更为丰富的ATP为之提供能量。而最近活体的荧光技术研究认为,后期时染色体牵丝在着丝点部位微管蛋白去组装大于极部而导致染色体拉向两极。
另一些人主张,染色体后期的向极运动的能量并不是来源于染色体微管,而是像肌肉收缩一样来源于肌球蛋白、肌动蛋白分子的相互作用。因为在纺锤体中发现有肌球蛋白、肌动蛋白分子,但用破坏肌动蛋白微丝的细胞松弛素B却不能阻止后期染色体的向极运动。
(2)滑动微管学说:此说认为后期染色体的向极运动力,来源于染色体牵丝和极间牵丝之间的相互滑行,像纤毛、鞭毛运动的机制一样。在纤毛、鞭毛上有动力蛋白横桥,可能在滑行中起作用。在有丝分裂细胞中可以提取出类动力蛋白,但在显微镜下却未发现纺锤体微管上有有规律排列的动力蛋白臂。滑行学说是建立在染色体牵丝和极间牵丝反向平行排列的基础上。实际上它们的极性是一致的,即为同向平行排列,因而难以用此说来解释染色体后期的向极运动。近期实验证明,若用纤毛的动力蛋白抗体处理细胞,后期B受到抑制,说明有一种类动力蛋白的蛋白后期B中起作用,而对后期A无影响。与此机制相似,前期内中心粒连同星体向两极移动与微管蛋白的亚单位加入两组星体的正极后星体不断延长,导致两组之间侧面结合的微管相互滑行而产生推动力这一现象有关(图3)。
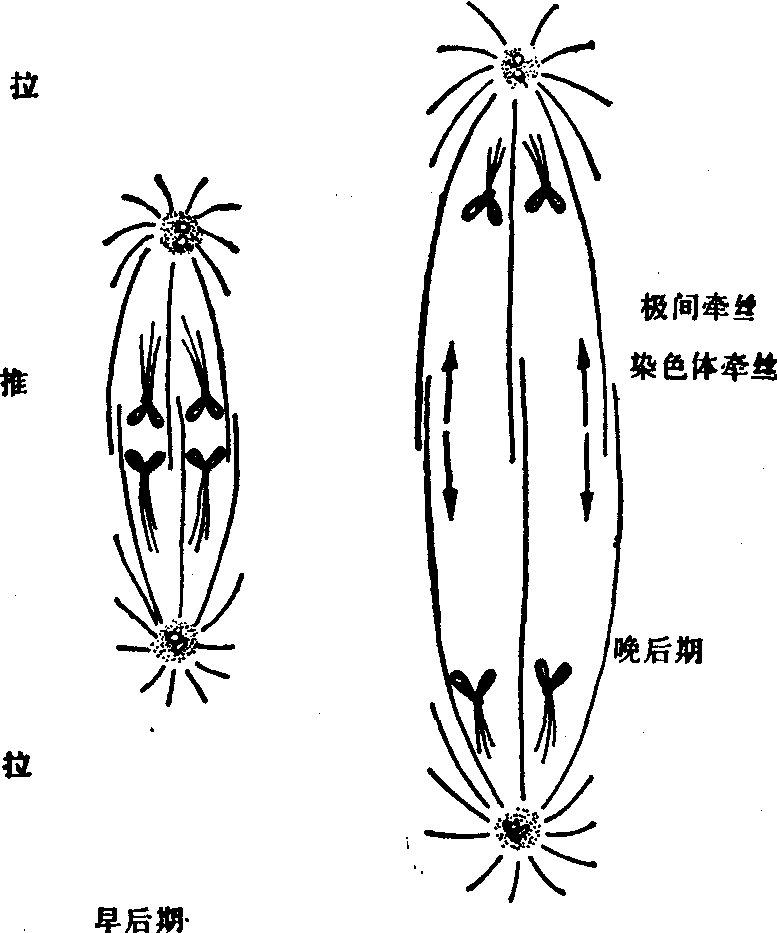
图3 作用于后期分离姐妹染色单体的推力和拉力
有丝分裂的控制 对控制有丝分裂的机制所知甚少,目前认为自由Ca2+的浓度是重要的调节因素。在体外实验中证明Ca2+是微管聚合的有力抑制剂,若将Ca2+显微注射到有丝分裂的细胞内,在注入Ca2+的附近可见双折光性暂时消失,但不久即恢复,说明胞质溶胶还有一种控制机构可以移去这些Ca2+。研究工作证明在分裂细胞中,纺锤体周围有大量表面的平滑膜囊,它可以按一定程序,定时地吸收或释放Ca2+,以调节微管的组装与去组装。大量工作进一步证明Ca2+ 是通过调钙蛋白而发挥作用的,它在分裂细胞中有相当高的浓度。
有丝分裂的原始类型 随着真核细胞的演化,有丝分裂的机构也随之复杂化,最后导致高等真核生物的有丝分裂机制的出现。许多低等真核生物的种类保持了一系列由原始到高级阶段的有丝分裂的中间过渡形式。例如涡鞭毛虫、眼虫等没有明显的染色体凝集和解凝集周期变化,或者只有一定程度的螺旋化和解螺旋。大量的低等真核生物在分裂时核被膜不破裂或仅部分破坏。最原始的涡鞭毛虫虽然产生了纺锤体,但只是成束地平行通过若干由核被膜围成的细胞质隧道,核被膜始终保持完整,此时纺锤体只是有助于确定分裂方向,通过两个子染色体与核被膜的结合点之间的生长,将两个子染色体分开。再高级一些的涡鞭毛虫只有中央一个隧道,两对中心体之间有纺锤体通过隧道,着丝点和内核被膜结合,靠核被膜生长,染色体被拉到接近两极,这时着丝点微管才同和着丝点隔着融合在一起的内、外核被膜相连接,拉着着丝点向两极移动。
有丝分裂的变异 高等真核生物某些种类的器官和组织,在自然状态下有丝分裂存在某些变异,常见以下类型:
❶胞质不分:形成二核或多核细胞,见于横纹肌细胞、体外培养细胞和某些植物雌配子的八核胚囊。
❷染色体后期不分开或进行核内有丝分裂(又叫内复制): 形成多倍体,见于哺乳动物肝细胞、某些昆虫和植物花药绒毡层细胞。
❸姐妹染色单体不分离:形成双分染色体,偶见于培养的人体细胞、植物花粉粒。
❹细胞周期中缺乏M期:核染色质反覆加倍而不分开,结果形成多线染色体,典型的例子是果蝇晚期幼虫的唾腺细胞。
❺体细胞减数分裂:可以产生单倍体,见于蚊子肠上皮细胞、玉米和水稻根尖细胞。
❻多极分裂:由纺锤体极部纵裂并转向,引起三极乃至更多极的核分裂。如果胞质也随之分开,便形成几个染色体数减少了的细胞。如果胞质未相应地分裂,则产生了一个多核细胞或数个核数目不等的细胞。多极分裂普遍存在于培养的癌细胞中。
有丝分裂
mitosis
有丝分裂
mitosis
有丝分裂
mitosis
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消丸是什么意思
- 内消乳疬方是什么意思
- 内消乳疬方是什么意思
- 内消化积丸是什么意思
- 内消升麻汤是什么意思
- 内消地胆散是什么意思
- 内消小豆散是什么意思
- 内消托里散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消散是什么意思
- 内消方是什么意思
- 内消方是什么意思
- 内消旋体是什么意思
- 内消昆布散是什么意思
- 内消沃雪汤是什么意思
- 内消沃雪汤是什么意思
- 内消沃雪汤是什么意思
- 内消法是什么意思
- 内消活关轻窍散是什么意思
- 内消浸酒是什么意思
- 内消漏芦煮散是什么意思
- 内消牡蛎丸是什么意思
- 内消牡蛎丸是什么意思
- 内消痔漏丸是什么意思
- 内消痔漏丸是什么意思