寄主—病原物互作interaction between host and pathogen
在病理过程中寄主和病原物相互影响和决定发病表型及其过程。相互作用可以在群体、组织和细胞以及分子水平上发生。互作类型与生化特征有关并受基因型组合调控。
互作类型 寄主—病原物互作分为亲和性互作和非亲和性互作两种。它们受到寄主和病原物基因型的调控。在寄主抗病/感病等位基因和病原物无毒性/毒性等位基因互作中,寄主抗病基因和病原物无毒基因都以显性为主要表现形式。在自然情况下,植物的抗病表型是普遍的,而感病表型仅在特定情况下发生。对病原物来说,一种病原物能侵染的植物在整个植物界中总是少数,多数是它的非寄主植物,对其侵染表现抗性反应。
亲和性互作 病原物侵染感病寄主并引起发病,这时寄主和病原物的关系为亲和性互作(campatible)。在此关系中病原物具有逃避寄主识别和钝化或破坏寄主防卫反应系统的能力。
基本亲和性互作 指寄主和病原物在种的水平上亲和,即一种病原物对某一寄主植物具有种的专化性。与此相对的一个概念叫基本不亲和性互作导致非寄主抗性。它表示一种植物对大多数病原菌,由于不是其寄主而不受侵染。从进化角度看,一种病原物能成功地侵染寄主植物是进化的结果,使其获得在一特定寄主植物上生长和完成生活史的能力。这些能力包括从寄主植物获得必要的营养、克服植物预存性抑菌物质和侵染后积累起来的植物保卫素。对植物保卫素的特异性抑制作用,以及产生毒素杀死或麻醉植物使防卫基因不能表达。在理论上,这种适应性变化的来源可能是自由生活的腐生菌和其他植物上的病原菌;适应性变化的机制可能有适应、突变和近缘杂交。从寄主角度看,这种亲和性互作也发生诱导防卫反应,使在一定程度上起限制病斑无限扩展的作用,寄主植物不至于完全溃烂和最终死亡。这种在基本亲和性中所表现的抗病性称为基础抗性。
具有小种特异性的亲和性互作 在具有小种特异性的病原物和寄主的相互关系中,亲和性的发生有三种情况:一是在多数情况下寄主的抗病表型决定于寄主抗病基因和病原物无毒基因的显性互作,即病原物无毒基因产物与寄主抗病基因产物相互识别的结果,诱导寄主产生防卫反应机制而表现抗病。因此,在病原物无毒基因和寄主抗病基因两者之一发生突变,上述的识别就不会发生,病原物因而逃避识别而引起寄主发病。二是在小种专化性表现为一个小种能特异性克服寄主中某一特定抗病基因的情况下,在马铃薯晚疫病系统中寄主植物所表现的抗病性,是病菌产生的非特异性激发子与寄主细胞膜表面受体结合并诱导植物保卫素合成的结果。但这种结合可以被病菌小种产生的特异性抑制子所阻碍,因而寄主从抗病反应转变成感病反应。三是以毒素为致病因子的病害,亲和性决定于寄主细胞膜(包括原生质膜和线粒体膜)上毒素的特异性敏感受体。
非亲和性互作 病原物遇上抗病寄主或非寄主植物,病害一般不会发生,这种关系为非亲和性(incompatible)互作,这时寄主植物表现对病原物的早期识别和随之产生的过敏反应或其他防卫机制。
非寄主抗病性中的互作 这是自然界存在最广泛的非亲和性互作类型。一种植物对非自身病原菌的抗性可以从营养角度、预存性抗病机制(包括结构性的和化学性的)和主动防卫机制等方面找到依据。从寄主和病原物的基因型进行分析,虽然也有可能存在基因对基因关系,但不会是一种植物对每一种潜在的病原物都有一个抗病基因。
小种专化性抗病性中的互作 以识别为前提和主动防卫反应为特征。识别发生在激发子与受体之间,主动防卫反应是由识别作用诱发的一系列按时序发生的生理过程。在小种专化性抗病性中,寄主中的每一个抗病基因都与一个无毒基因互作。在这里寄主抗病表型通常是由单基因控制的,而病菌的无毒表型则可以由单基因或寡基因所控制。在对毒素为致病因子的病害抗性中,寄主的作用可以表现为降解毒素或对毒素的损伤具有修复机制。
互作类型的发育联系 1976美国爱伦堡(A.H.Ellingboe)等认为植物品种的小种专化性抗性是从基本亲和性发展而来。1984英国卡罗(J.C.Callow),阐述了四种互作类型的进化联系即: 1.在种的水平上的不亲和性(非寄主抗病性);2.在种的水平上的基本亲和性;3.品种抗病性;4.病原物小种与寄主植物品种的亲和性。(图1)。
非寄主抗性是由于病菌产生的激发子(E)与寄主感受蛋白(S)识别后激发产生诱导抗性的结果。
基本亲和性是由于病菌产生抑制子(Su)与激发子(E)竞争感受蛋白(S),使激发诱导抗性的过程阻断后发生的。
小种专化性抗性 是在病害的选择压力下通过突变或者育种使寄主获得了抗病基因(R),其编码产物R′是一种新的感受蛋白,对病原物表面特异性信号分子(A′)具有识别作用。R′与A′互作产生对具有无毒基因A的小种专化性抗性。
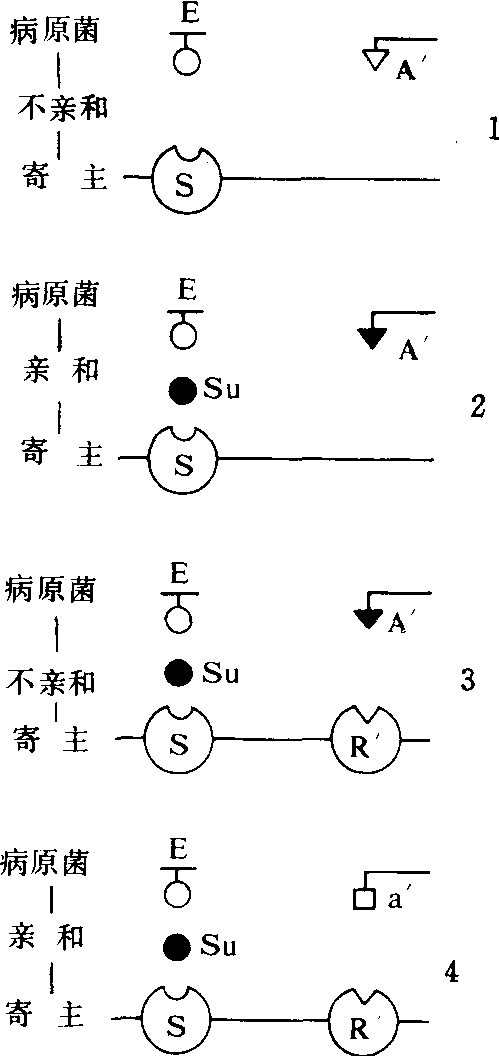
图1 寄主—病原物不同互作类型的发育联系
E:激发子 S:感受蛋白 A′:无毒基因产物 R′:抗病基因产物 Su:抑制子 a′:无毒基因突变后的产物
(仿Callow,J.C.1987)
小种专化性互作的亲和反应 在小种专化性抗性中,病菌的无毒基因处于选择压力之下,突变后的产物a′不被寄主识别,从而产生亲和性互作。无毒基因是对病菌不利的,在进化中经常被选择掉,使原有品种的抗病性丧失。
生化、遗传特征 寄主—病原物的相互作用可以分为识别和表达两个阶段。识别作用的结果决定功能表达。
识别 指寄主和病原物相互作用早期发生的过程。大多数互作模式都是针对这一阶段提出的。
锁钥关系 1982年美国孔恩(N. T,Keen)提出用以比喻寄主—病原物关系的。钥匙上的缺刻,有些决定基本亲和性,有些决定互作特异性。缺刻的排列组合决定互作关系多样性。
表面分子特征 1984年英国卡罗(J. A. Callow)以基因对基因关系为例阐明了寄主与病原物识别的分子内涵(图2)。寄主抗病性(R)/感病性(r)等位基因的产物是 一类位于细胞表面,具有位点结合构型的受体(recepter)。而病原物无毒性(A)/毒性(a)等位基因产物则是一类结构相同而识别末端不同的分子。只有在寄主抗病基因产物和病菌无毒基因产物功能互补的情况下(R/A),结合和识别才能发生,从而诱导产生寄主抗病性。在主动抗性模式中寄主参与抗病性的基因有两类: ❶与病菌无毒基因互补的植物抗病基因(R),抗病基因产物只起识别作用,因此称效应分子。
❷被识别作用激发的寄主防卫基因,这些基因产物或次生产物有抑菌或抗病作用,因此称作用分子。
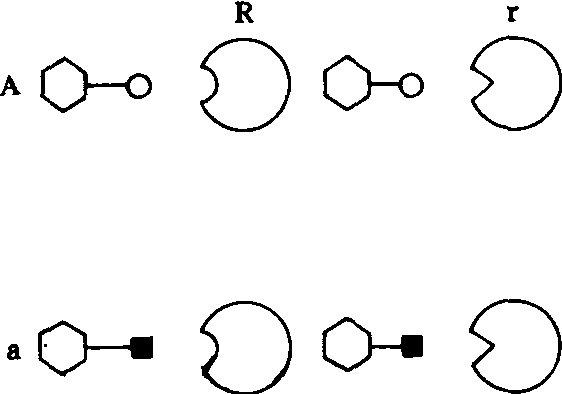
图2 寄主—病原物表面分子识别示意图
A:无毒基因显性 a:无毒基因隐性R:抗病基因显性 r:抗病基因隐性
(仿Callow,J.C. 1987)
功能表达 由识别作用激发诱导的防卫反应基因表达可以是新基因转录或原有蛋白的激活或聚合。
新基因的转录 在植物保卫素诱导合成中证明有新基因转录和翻译。植物接种非亲和小种后短时间内合成的polyA-mRNA(含多聚腺苷酸信使核糖核酸)要比对照高数倍,其中可以发现新型的mRNA,稍后出现新型蛋白质。如在接种后以转录和翻译抑制剂放线菌素(Actinomycin D)和灭瘟素(Blasticidin)处理,上述转录和翻译过程就会被阻,植物保卫素的合成受到抑制。
原有蛋白的激活或聚合 见于许多主动抗病反应过程如水解酶的产生,外缘凝结素的合成,病程相关蛋白的产生等。
表1 寄主—病原物互作中的病原物信号分子
| 信号分子类型 | 生理功能 |
| 花生四烯酸和二十碳五 烯酸 | 植保素合成 |
| 真菌糖肽 | 富含羟脯氨酸糖蛋白的积 累 |
| 真菌孢外糖蛋白 寡聚半乳糖醛酸DP= 2—6<12 | 决定亲和、不亲和反应 蛋白酶抑制剂Ⅰ和Ⅱ合成 |
| DP≥10 环状β-1,2-葡聚糖 | 植保素合成 增强豆科植物根毛感染和 结瘤 |
| 七-β-葡糖苷直链醇 (β-3,6葡聚糖) | 植保素合成 |
| 果胶降解物 脂多糖(LPS) | 决定亲和、不亲和反应 增加豆科植物根毛卷曲和 结瘤 |
| 荚膜多糖(CPS) 胞外多糖EPS EPS寡聚糖片断 | 增加根毛卷曲和结瘤 根毛卷曲 根毛卷曲和感染 |
表2 寄主—病原物互作中的植物信号分子
| 信号分子类型 | 生理功能 |
| 酚类化合物 | |
| 芳香族化合物 单环化合物(乙酰丁香 酮) | 诱导微生物分解代谢基因 |
| (α-乙酰丁香酮) | 诱导根癌土壤杆菌Vir基 因表达 |
| 黄酮类物质(淡黄/木樨 草苷,苯基苯乙烯酮,毛地 黄黄酮) | 诱导根瘤菌结瘤基因表达 |
| α-生育酚(维生素E) | 诱导Ustilago violacea寄 生菌丝 |
| 半乳糖醛酸类物质 | 诱导真菌Phytopthora spp.孢子萌发及囊化反应 |
| 大豆外缘凝结素SBL (soybean lectin) 环β-1,2-葡聚糖 | 诱导Bradyrhizo bium japonicum结瘤基因表达 诱导根癌土壤杆菌吸附寄 主及Vir基因表达 |
| 奥品那冠瘿碱(Opines) | 诱导Ti质粒转移 |
| 植保素 | 诱导病原产生植保素甲基 化酶 |
| 病原酶底物或其降解物 | 诱导病原胞外酶合成 |
信号传递 寄主—病原物互作涉及到一系列相互联系的细胞化学反应。最先引发反应的物质即信号分子。反应中按时序发生的过程即信号传递。
信号类型 信号传递具有双向含义,寄主植物接受病原物的信号引发防卫反应,病原物接受寄主植物的信号可诱导产生降解酶或起动致病基因(表1、2)。
寄主接受的信号 有病原物细胞壁的组分,病原物产生的酶,病原物酶降解植物细胞壁产生的物质,寄主植物酶降解病原物细胞壁产生的物质等。
病原物接受的寄主信号 有植物受病原物酶降解产生的细胞壁组分的单体、寡聚体和其他成分。
信号传递 已提出三种模式:❶在主动抗性反应中,由激发子作为信号经跨膜感受蛋白传递。
❷在细胞壁修饰中,刺激胼胝质合成和积累的激发子如脱乙烯几丁质是通过嵌入细胞壁脂相后与膜磷脂顶部基因或与甾醇结合来传递的。
❸在过敏反应中包括受体活化磷酸脂酶C,产生第二信使、肌醇三磷酸盐和甘油二酯,进一步刺激钙离子流、活化蛋白质激酶和释放脂肪酸,从而活化交换反应,最终细胞死亡。这一模式的特点是离子通道反应位于受体下游,是由第二信使开放的。
- 朱雀、玄武是什么意思
- 朱雀丸是什么意思
- 朱雀丸是什么意思
- 朱雀丹是什么意思
- 朱雀桥边野草花,乌衣巷口夕阳斜。旧时王谢堂前燕,飞入寻常百姓家是什么意思
- 朱雀桥边野草花,乌衣巷口夕阳斜。旧时王谢堂前燕,飞入寻常百姓家。是什么意思
- 朱雀汤是什么意思
- 朱雀汤是什么意思
- 朱雀汤是什么意思
- 朱雀航是什么意思
- 朱雄丸是什么意思
- 朱雄丸是什么意思
- 朱雅清是什么意思
- 朱集汉是什么意思
- 朱集禧是什么意思
- 朱雍模是什么意思
- 朱雪琴是什么意思
- 朱雪芳是什么意思
- 朱雪龙是什么意思
- 朱雯是什么意思
- 朱雯是什么意思
- 朱雯是什么意思
- 朱雯·罗洪读书是什么意思
- 朱霁青是什么意思
- 朱霁青是什么意思
- 朱霁青是什么意思
- 朱霁青是什么意思
- 朱霁青是什么意思
- 朱震是什么意思
- 朱震是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨是什么意思
- 朱震亨的传说是什么意思
- 朱霖是什么意思
- 朱霖是什么意思
- 朱青冰是什么意思
- 朱靖华是什么意思
- 朱静是什么意思
- 朱静是什么意思
- 朱静是什么意思
- 朱顺佐是什么意思
- 朱顺泽是什么意思
- 朱颂伟是什么意思
- 朱颂周是什么意思
- 朱颖人是什么意思
- 朱颖铨是什么意思
- 朱颜是什么意思
- 朱颜是什么意思
- 朱颜绿鬓是什么意思
- 朱馥生是什么意思
- 朱驎是什么意思
- 朱马利耶夫是什么意思
- 朱马勒是什么意思
- 朱马格列季耶夫是什么意思