发生遗传学
又称“发育遗传学”。遗传学的分支学科。研究生物的基因型如何转化成表现型。主要利用影响发育的突变型,并结合实验胚胎学、细胞生物学和分子生物学的方法、从不同水平分析基因和性状发育之间的关系,以阐明基因控制发育的机理。
发生遗传学
发生遗传学是发生学与遗传学相互渗透产生的一门边缘学科,研究基因如何表达成为生物性状的规律; 换句话说,是研究生物的基因型如何转化成表型。这个过程很复杂,是通过一系列生化反应实现的,但了解的还很少。
在多细胞生物中,一个完整的个体是由亿万个细胞所组成的,而这些细胞又都是由一个受精卵发育而成的。因此,一个个体中不同细胞的基因型应该都是相同的。可是具有相同基因型的细胞在不同时间内和不同空间内又能分化成为构造不同、功能有别的组织。从原理上说,这是由于部分基因有选择地表达所造成的。
一个个体中,各细胞的基因型既然相同,为什么有的细胞中某一基因能够表达,而在另一些细胞中却不能表达;不同的基因为什么只在不同时间或不同空间得到表达?这些问题的实质是基因调控的问题,也就是基因表达的条件问题。现代生物学基本的论点是: 任何基因只有在一定条件的作用下,才被激活,从而得到表达。否则,就处于被封闭状态,而不能发生作用。所谓一定的“时间”或“空间”可能就在于提供某些条件来激活某些基因,使之得到表达的机会。当然,当一个基因表达之后,其产物也可以成为激活其他基因的条件,这就是所谓基因相互作用机理之一。所以,从原则上来说:一个个体就是基因型和条件相互作用的产物(基因型+条件→表型)。虽然原核生物和真核生物(包括动物、植物和人)都有发生遗传学的问题,但是,因为真核生物(特别是高等动物)的性状发育时间较长,变化较多,研究困难较大;而另一方面,由于原核生物,结构简单,生活周期短,新的理化技术易于应用,所以,原核生物很早就被人们选为研究材料,从而在分子水平的研究中,发现了一系列发生遗传学的规律: 如操纵子学说以及基因表达的正控制和负控制等概念(见“细菌代谢的遗传控制”)。
激活或调节基因的条件,可以来自细胞的内部,也可以来自细胞的外部。下面试以原核生物大肠杆菌乳糖操纵子中的半乳糖苷酶基因的表达与调控为例予以说明。大肠杆菌的乳糖操纵子内含有两大类基因。一类是结构基因,如β半乳糖苷酶基因、透酶基因和乙酰化基因等。这类基因活化后可以产生相应的酶。不过这些结构基因的活化却受到细胞内DNA水平上另一类基因——调控基因的调节和控制,如操纵基因、启动(基因)和调节基因等等。可是乳糖操纵子中结构基因的表达,除受到细胞内调控基因的调控外,同时也受到细胞外的环境(培养基)中乳糖含量的影响。
总的说来,原核生物操纵子和基因调控对分子遗传学所提出的新概念是把基因分为结构基因和调控基因两大类。
虽然原核生物和真核生物都有发生遗传学的问题,可是目前研究的重点集中在真核生物。真核生物和原核生物在基因表达的问题上有许多不同,但是其基本不同之一在于原核生物的转录与转译几乎是同时进行的; 而在真核生物的细胞中,这两个过程无论是在时间上或空间上都是分开进行的:转录是在细胞核内进行,转译是当核内转录出mRNA并运到细胞质后,在核糖体上进行的。所以,真核细胞的调控过程,基本上可以分为:
❶转录前的调控和
❷转录后的调控。此外,目前还发现,已经转译好的多肽链(一级结构)在形成蛋白质的二级或三级结构过程中也可以受到某些细胞内部或外部条件的调控,甚至在蛋白质分子中四聚体的组装过程中也还有机会受到调控。此外,在高等动物中,调控基因在质和量以及相互协调、相互作用上,比原核生物大为扩增和更为复杂。这就给研究这些动物的基因调控,设下极大障碍。直到目前为止,在高等动物中还未发现一种统一的、可以指导一切的基因调控的基本规律; 只是在若干独立系统中能深入到分子水平。下面试举两个例子予以说明。例如,在脊椎动物发育的细胞分化中,对检验基因功能常涉及到蛋白质合成的是乳酸脱氢酶(LDH)。LDH在葡萄糖无氧分解中起重要作用。把小鼠的骨胳肌制成匀浆,然后进行电泳分析和特殊染色,可以分离出五种不同的LDH同功酶。经过分离和化学分析,证明这些酶是由分子量一样的α,β两种亚基所构成的四聚体。这两种亚基是由相应的A和B两个结构基因在不同时空条件的调控下合成的。这两种亚基在四聚体里的数量随机组合上,可能有如下五种区带,各区带有不同的电荷,能被电泳分解成为五种不同的同功酶(图1)。
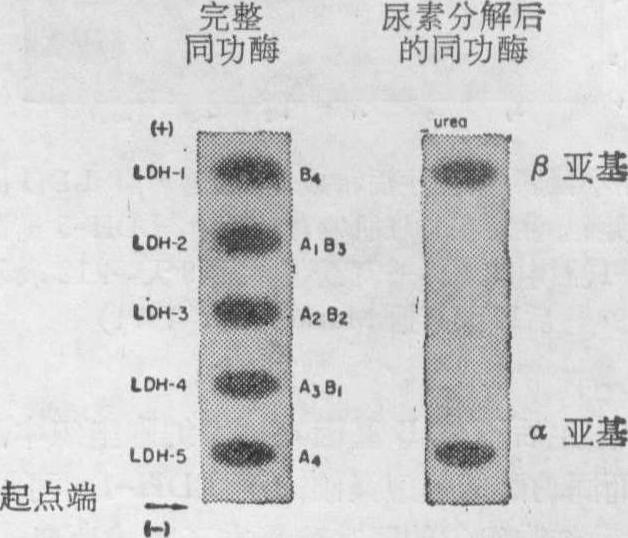
图1 表示小鼠骨胳肌的五种LDH同功酶的构型,
左边为完整的酶区带;右边为经尿素处理后再进行电泳分析的结果,即只有LDH-1(B4)和LDH-5(A4)区带出现(据Markert等,1971)
以上五种同功酶,确实是由α和β两种亚基随机组合的产物,可将LDH-1和LDH-5分离出来先进行冷冻后再溶解把其亚基拆散,然后使LDH分子在适当的盐溶液中重新随机组合,将会发现不是两种,而是五种电泳区带以二项式定律分布的比例显示出来(图2)。
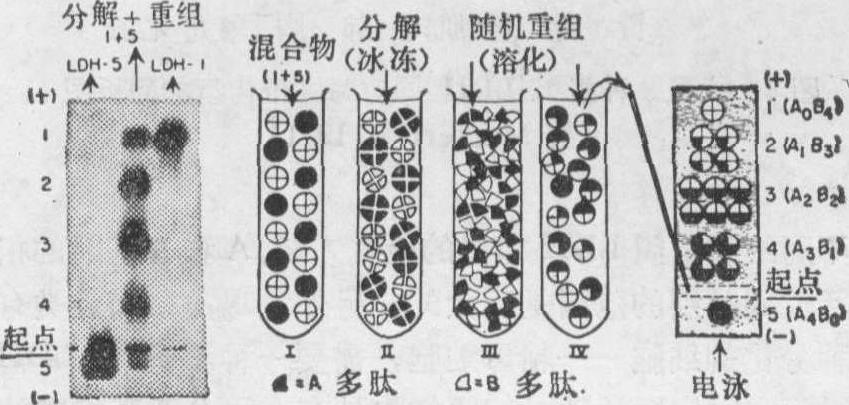
图2 表示LDH-1与LDH-5混合物的分解和
重组模式图。显示五种LDH同功酶的二项式定律的区带分布(据Markert等,1971)
对一种动物的每种类型的细胞组织,同功酶的构型具有高度的特异性。这是由α和β亚基的有无和数量的多少构成的,同时也是在动物个体发育不同时期中,A或B基因具有不同活性的结果。例如发育中的小鼠卵子只有LDH-1,可见在这一阶段只有B基因起作用。随着胚胎发育的进行,B基因的活性渐被抑制,而A基因却被激活。到了小鼠胚胎发育的第9天,A基因比B基因更加活跃起来,这就使在分化的细胞组织中,同功酶的构型偏向于LDH-5(图3)。随同胚胎进一步地发育,特别是在
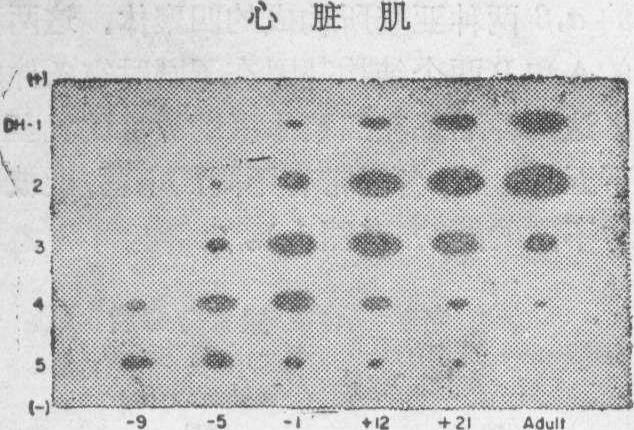
图3 在小鼠胚胎、出生后和成体心脏组织里LDH同功酶构型的变换。注意在心脏肌发育过程中,LDH-5渐渐偏向于LDH-1(图中横座标—9表示出生前9天,+12表示出生后12天)(据Markert等,1971)
出生时和出生后不久,B基因在许多细胞组织中,再度活动起来,而同功酶的构型又倾向于LDH-1。可见,在动物体内各种细胞组织的同功酶具有不同的构型,恰恰是反映着在它们发生过程中的不同阶段,A和B基因合成其相应亚基比例不同的结果(图4 )。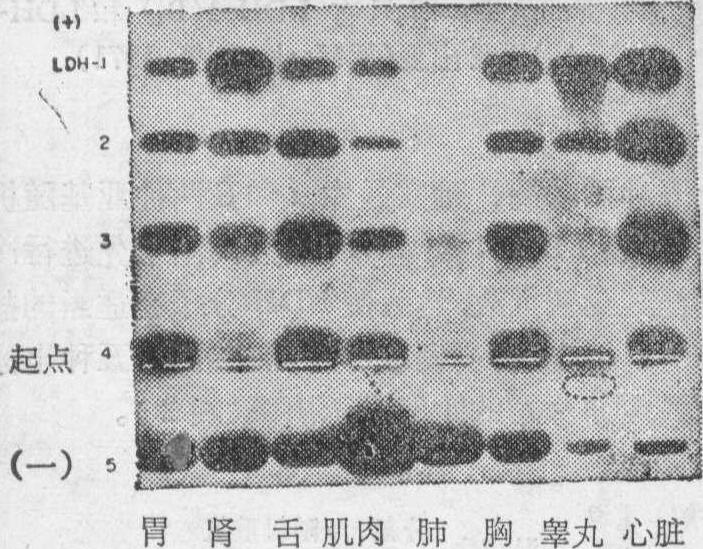
图4 大鼠8种组织LDH同功酶区带构型的酶析图(据Markert等,1971)
不同细胞组织LDH构型的检查表明,A和B基因的相对活性间差别的范围是巨大的。另外,鸟类和哺乳类有一种分化的细胞——精母细胞,需要一种不同的LDH来进行它的代谢活动。精原细胞具有α和β亚基组成的LDH同功酶。然而,在精母细胞开始分化时,A和B基因即被抑制,而第三种叫做C基因的却合成不同形式的LDH。由此产生的C四聚体,是一种由α和β亚基所组成的五种不同LDH截然不同的同功酶。C基因提供着一个特定时间上和特殊细胞中基因专一功能的例子,它在动物体内除初级精母细胞外,在一切细胞中都是不活动的。
在许多被检查过的鱼类,在它们的视网膜和某些神经细胞中,还发现一种额外的LDH同功酶。这一LDH的多肽是由一个E基因所编码,它只出现在成体细胞。E基因也只在少数几种细胞发育的一定的阶段内开始表达。在E基因内编码的多肽,形成E4四聚体,并也同α和β亚基共同聚合,造成在这类细胞中同功酶构型的多样性(图5)。
不同LDH基因有选择性的激活,最终要归因于细胞质的当时当地的理化成分,而这一成分反过来又受调控LDH亚基合成基因的影响。在正常情况下,LDH的β基因几乎在所有的小鼠细胞里都是活动的。
在人类发生遗传学深入到分子水平的,莫过于血红蛋白合成的研究了。合成血红蛋白的基因调控是一个相当复杂的过程,包括血红素合成与珠蛋白合成的关系以及由胎儿血红蛋白向成体血红蛋白转变等问题。
影响血红素生产的主要机
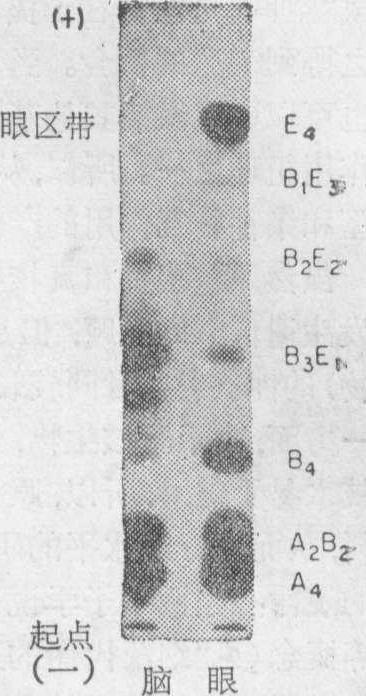
图5 表示在青花鱼(Sco-mber scombrus)眼和脑组织含有E亚基LDH同功酶的酶析图(据Markert等,1971)
理,也就是合成的第一步,是产生δ-氨基乙酰丙酸(ALA)。ALA的产生反过来又为ALA合成酶所制约。ALA合成酶是一种线粒体的酶,但却是由核基因编码。酶的合成虽在细胞质的核糖体上,但这一初级产物是没有活性的酶,而其激活却发生在线粒体内。所以,具有活性的酶促成血红素的合成是在线粒体内,然后才释放到细胞基质中。血红素的存在能影响血红蛋白生产的速度。在无细胞系统里,自由血红素的加入,增加了珠蛋白的合成;动物的实验也提供类似的结果。有迹象表明血红素存在的作用点是在血红蛋白合成的启动基因上面。本世纪五十年代末和六十年代初英国剑桥大学的研究者们,发展了纸上电泳和色层分析并把二者结合使用,进行血红蛋白氨基酸的排列顺序的鉴定。这就为研究血红蛋白的基因调控创造了有利条件。血红蛋白和许多其他蛋白质一样,正常具有四聚体的结构,其分子是由重复起来的两对不同的亚基所组成。每一个亚基又是由141或146个氨基酸合成的肽链,围绕着一个带有铁原子的血红素而摺叠起来。正常成人最常见的血红蛋白是Hb A,这是由两个α链和两个β链所组成的四聚体。极少数红细胞还具有一种浓度较低的血红蛋白,叫做HbA2,由两个α链和两个δ链所组成。HbA和HbA2的比例,保持在97.5%A和2.5%A2相当恒定的数量上。怀孕后3到6个月之间的胎儿的血红蛋白叫做Hb F,是由两个α链和两个γ链所组成。由Hb F向HbA的转变,涉及到正在分娩或即将分娩期间,γ链合成逐渐减少而至停止,并为β链的产生所代替。但α链的合成却在胎儿和成体期间继续不断地进行着。研究血红蛋白系统的有利条件之一,是血红蛋白的产生无例外地都在红细胞和其前身细胞以内。在早期人胚卵黄囊细胞里,最先出现的原始红细胞是在所谓“血岛”之中。这样的原始红细胞经常保留它们的细胞核。在后来的发育过程中,红细胞的生产区逐步向胚胎的内部移动,而其他组织如肝、原肾、淋巴结、脾和骨髓都先后曾经成为红细胞的发源地。
胎儿和成体的红细胞都是来自相对未分化的干细胞。这样的细胞也都具有共同的干系,叫做成红细胞。这些细胞是变形虫样的,嗜碱性,但在细胞质内没有任何颗粒。纵然正在循环着的红细胞代表一种相对同质的细胞群,但在一个特别基因的多肽链产物上却极为不同。血红蛋白的合成发生在红细胞被释放到循环系统之前。这一事实对研究基因的表达,极不方便,因为基因活动转变机理中的关键步骤发生在红细胞的产地,而这一组织远非同源,因而成为一种比较难于掌握的实验材料。对于这个问题的解决办法有二:
❶把产生红细胞的组织在半固体的琼脂培养基上进行离体培养;
❷利用严重贫血动物做实验,因为在这样的动物体内,血红蛋白合成途径中的许多早期分化步骤,都发生在循环着的细胞里,而且这样的细胞可以在组织培养中进行实验和观察。
观察的结果证明,从胎儿到成体有四个基因:α、β、γ与δ牵连到正常血红蛋白的合成。各种血红蛋白的合成中的根本问题,是从胎儿到成体的转变。这两种血红蛋白的差别是在胎儿血红蛋白中,γ链代替了β链,因而使胎儿血红蛋白对碱稳定。究竟为什么胎儿需要一种不同于成体的血红蛋白,还是一个未解决的问题。诚然,在胎儿的循环系统里,血红蛋白对氧的亲和力较成体为高,可给予胎儿一种特定的生理优势,因为在它这样的系统里,氧气从母体循环到胎儿体内,必须通过胎盘这一关卡。联系到某些遗传的血红蛋白病的患者,如β型海洋性贫血(重型)是有特殊意义的。在这些贫血症者成体的循环系统中,经常存在着含有大量的胎儿血红蛋白的细胞(图6)。
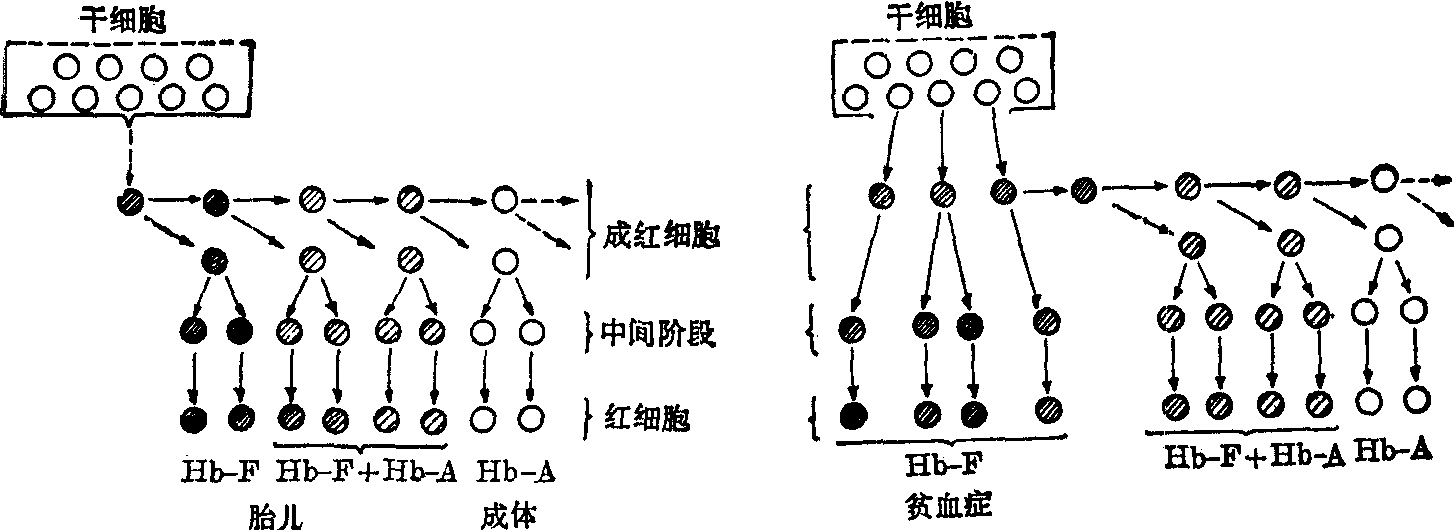
图6 表示从HbF到HbA在胎儿末期的转变(左)和严重海洋性贫血症产生HbF的机理(右),红细胞分化的模型。圆圈里画线多少的程度显示红细胞前体在分化不同阶段合成HbF的相对数量(据Baglioni,1962)。
从胎儿到成体血红蛋白成分的转变,可以用曲线图表示。从这样的图中可以看到α链自始至终维持着100%的生产,γ链开始时是和α相似的,但在胎儿出生前,即已逐渐下降,到了出生后6个月的时期,即由刚出生的50%下降到0%。与此同时,β链却有一个相应逐渐增生过程,并在婴儿出生后6个月的时期,完全代替了γ链。
根据上面罗列的事实,Ingram等人提出一个血红蛋白在发育中转变的假设。这一假设认为当一个干细胞的后代转入成红细胞时,血红蛋白便开始合成。这意味着一切合成血红蛋白的基因,都在干细胞中受到阻遏,它们的分别激活,是在成红细胞阶段。在这一分化阶段中,β、δ、γ三个基因当中哪一个被激活,取决于两个因素:一是干细胞所经历的分裂次数,一是在细胞分化前它在那个特殊环境停留时间的长短(图7)。
干细胞分裂的次数少,又停留在胎儿体内环境的所谓“青年”成红细胞里,γ基因得到转录,再由mRNA转译成γ链。在这过程中,细胞里一定还生产着对β和δ基因转录的阻遏物。随着干细胞继续不断的分裂和γ基因转录逐渐变慢,β和δ基因却相应地逐渐活跃起来。到了婴儿出生后6个月的时期,由于干细胞及成红细胞又经历了多次分裂,并处于与胎儿体内截然不同的环境之中,
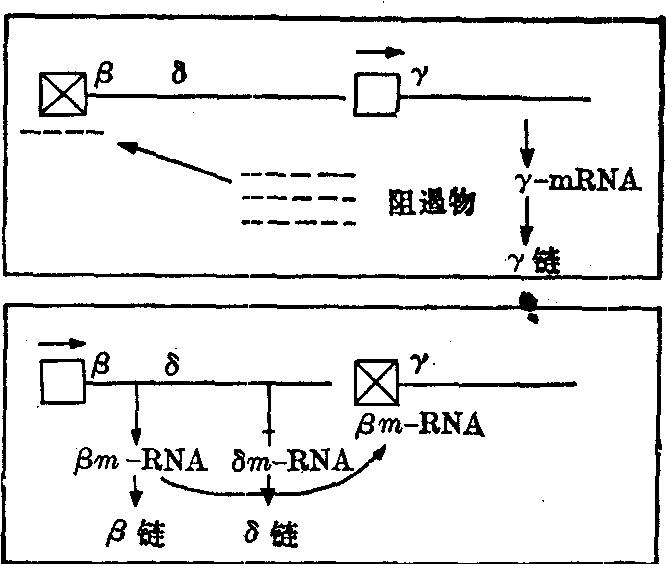
图7 表示胎儿末期从Hb-F到Hb-A转变控制机理的模型(据Ingram,1963)
所有的成红细胞都已达到“成熟”。成熟的成红细胞只能产生β和少量δ链,而在这时,γ基因却被阻遏。可以说人类红细胞中珠蛋白基因是在不同时间的序列中被激活而表达的。
Ingram的假说当然仍需要进一步的验证,可是他们提出了一个富有启发性的概念,对真核生物的发生遗传学研究做出了贡献。
发生遗传学
developmental genetics
- 金枝玉叶是什么意思
- 金枝玉叶是什么意思
- 金枝玉叶是什么意思
- 金枣丸是什么意思
- 金枣丹是什么意思
- 金枣丹是什么意思
- 金枣丹是什么意思
- 金枣儿是什么意思
- 金枣儿是什么意思
- 金枣儿是什么意思
- 金枣儿是什么意思
- 金枣化痰丸是什么意思
- 金枣化痰丸是什么意思
- 金枪丹是什么意思
- 金枪散是什么意思
- 金枪膏是什么意思
- 金枪药是什么意思
- 金枪铁扇散是什么意思
- 金枪鱼是什么意思
- 金枪鱼是什么意思
- 金枪鱼(Thunnus tonggol)是什么意思
- 金枪鱼围网渔业是什么意思
- 金枪鱼围网渔船是什么意思
- 金枪鱼延绳钓船是什么意思
- 金枪鱼渔业是什么意思
- 金枪鱼类和金枪鱼渔业是什么意思
- 金柑是什么意思
- 金柑是什么意思
- 金柜散是什么意思
- 金柱关之战是什么意思
- 金柱塔是什么意思
- 金柱演是什么意思
- 金柴胡是什么意思
- 金柴胡是什么意思
- 金树仁是什么意思
- 金树仁是什么意思
- 金树仁是什么意思
- 金树仁是什么意思
- 金树仁是什么意思
- 金树基是什么意思
- 金树泽是什么意思
- 金树滋是什么意思
- 金根是什么意思
- 金根是什么意思
- 金根宽是什么意思
- 金根车是什么意思
- 金桂月挂屏是什么意思
- 金桃姑娘是什么意思
- 金桃酒是什么意思
- 金桢轸是什么意思
- 金桥出口加工区是什么意思
- 金梁是什么意思
- 金梁是什么意思
- 金梁段子八吉祥帽儿是什么意思
- 金梅感冒片是什么意思
- 金梅牌兰花豆(片)是什么意思
- 金梅牌江苏老酒是什么意思
- 金梅生作品选集是什么意思
- 金梦庚是什么意思
- 金梨是什么意思