内质网endoplasmic reticulum
细胞质内参与细胞蛋白质合成和类固醇激素、脂类、糖原代谢及具有其他多种功能的膜性管道细胞器。可分两类:膜上附有大量核糖体者; 称粗面内质网或有粒内质网(RER或GER); 膜上无核糖体附着者,称滑面内质网或无粒内质网(SER或AER)。二者彼此相连,但功能不同,在不同类型细胞中所占比例不同,且可随细胞功能状态的变化而相互转变。在靠近细胞核处,内质网膜可与核被膜外层相连续,内质网腔与核周隙相通; 靠近细胞膜处,可能与细胞膜内褶部分相通连。
发现史 有些学者早已发现细胞质中存在一种核外染色质或动质。电镜技术发展以后,人们对内质网才有所认识。1945年,波特(K. R. Porter) 等观察组织培养的鸡胚成纤维细胞时,首先看到一些交织成网的膜性管道,分布在细胞核周围的细胞内质中。其后已证实内质网可分布在细胞的各个部分。所谓核外染色质,实际上是结合在粗面内质网膜上的核糖体,因其量大而密集,在常规染色的切片标本上呈现类似染色质的嗜碱性斑块; 动质一词,则表达了其含量随细胞生理状态而变化的特性。
化学组成 内质网膜比细胞膜薄,厚约5~6nm,其化学组成资料是对通过蔗糖密度梯度离心法从细胞匀浆中分离出内质网膜碎片围成的微粒体进行生物化学分析获得的。内质网膜的脂类物质中,不饱和磷酯含量较高,胆固醇含量较低,与细胞膜比较有明显的差异,细胞膜中饱和磷酯的含量较高,胆固醇的含量比内质网膜多。内质网膜所含蛋白质,除结构蛋白外,重要的酶类有NADPH—细胞色素c还原酶、细胞色素b、细胞色素p-450、腺苷三磷酸酶(ATP酶)、葡萄糖-6-磷酸酶、以及与合成三酸甘油酯、磷酯和胆固醇等有关的酶类。脂类与蛋白质的比值,滑面内质网膜较高,粗面内质网膜较低。粗面内质网膜还含有大量RNA,这是由于其上结合有核糖体。
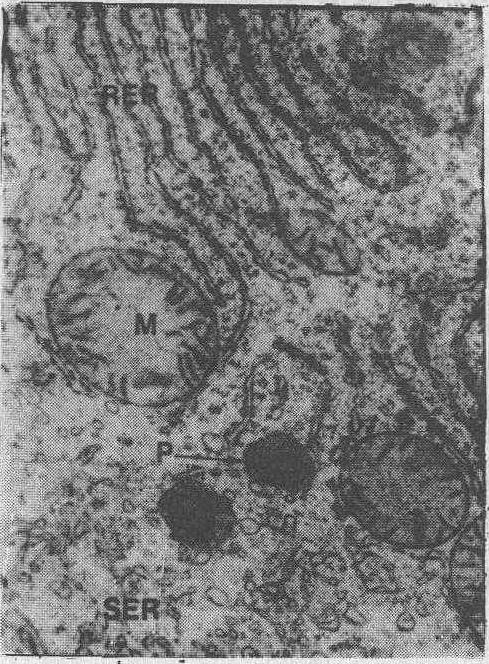
中国仓鼠细胞内质网电镜图象M. 线粒体;RER. 粗面内质网;P. 过氧物酶体;SER. 滑面内质网
粗面内质网
形态结构 大多由相互通连的扁平囊组成,少数呈游离的小泡和小管状。在粗面内质网非常发达的浆细胞和胰腺细胞中,扁平囊排列紧密,在细胞核周围形成同心层板状结构,占细胞体积的大部分(见图)。同一类型细胞的粗面内质网也是可变的细胞器,其数量、大小、囊腔(网池)宽度、核糖体多少、内容物均可发生变化。如蚕豆种子在发育早期,子叶细胞的粗面内质网较少,当累积贮存蛋白质时,粗面内质网变得非常丰富; 胰腺细胞的粗面内质网在分泌旺盛期至静止期明显地由密集变稀少; 浆细胞在免疫球蛋白累积过多或分泌机制发生紊乱时,粗面内质网中出现被称为罗氏小体的蛋白小体; 细胞处于某些病理条件下,如四氧化碳中毒时,粗面内质网膜上的多聚核糖体可发生解聚或脱落等。
功能 粗面内质网膜上结合有附着核糖体,是细胞合成蛋白质的重要场所。主要合成输出蛋白,包括各种肽类激素、酶类和抗体等,也能合成若干种结构蛋白,如构成新膜的蛋白和酶以及溶酶体、微体、液泡和圆球体中的一些酶类。
近年有些学者提出信号学说,认为核糖体之所以能附着于内质网膜,关键在于输出蛋白和粗面内质网上合成的其他蛋白质的信使核糖核酸(mRNA)比较特殊,即在其启动密码后面还有一组编码信号肽链的序列。这类mRNA在核糖体上进行翻译时,必须先合成大约由11~26个氨基酸组成的信号肽链,当其达到一定长度,信号肽链从大亚基暴露出来,被游离在胞质中的信号识别颗粒识别,并与之结合,这时蛋白质合成暂时停止。一旦信号识别颗粒一核糖体复合物附着在内质网膜上的停泊蛋白(信号识别颗粒的受体)上,信号识别颗粒随即脱落,游离于胞质中,并可再次参与识别与停泊过程。与此同时,肽链通过内质网膜上形成的临时孔道进入内质网腔,核糖体也开始重新合成蛋白质,不断伸长的肽链陆续移入内质网池,信号肽链则被附着在内质网膜内侧的信号肽酶切除。当蛋白质合成终止信号出现时,合成停止,新生肽链完全进入内质网腔,核糖体从内质网膜脱落并解聚,进入内质网腔的肽链经修饰和变构,最后形成具有一定空间构像的蛋白质。如不是分泌蛋白,而是结合在内质网膜本身的蛋白质时,新形成的肽链则在合成过程完成之前,即可边合成边从内质网腔转移入膜中(见蛋白质代谢)。
粗面内质网合成的输出蛋白如何输送到高尔基复合体的说法不一。有人认为内质网与高尔基复合体是连续结构,输出蛋白直接从前者输送至后者。有人认为内质网以出芽方式形成运输小泡,将输出蛋白转运给高尔基复合体。马苏拉(Matsuura)等提出粗面内质网与高尔基复合体之间有一滑面的移行部分,其膜上有输出蛋白的受体,并具有与高尔基复合体膜融合的能力,输出蛋白与之结合即可形成运输小泡,将输出蛋白并入高尔基复合体。
粗面内质网还可将合成的产物(如酶类)与细胞质中其他成分隔开,并将其输送至高尔基复合体。有些可溶性蛋白质则通过粗面内质网膜被直接转运至细胞质基质,粗面内质网还可合成一些脂类。
滑面内质网
形态结构 在常规染色切片标本上,因其与嗜酸性的细胞质染色反应一致,故无法察见。当大量存在时,细胞呈强酸性,如胃底腺的壁细胞、睾丸间质细胞和肾上腺皮质细胞。当其集中存在于细胞的局部区域时,则呈粉红色的嗜酸小体。滑面内质网膜比粗面内质网膜略薄,缺核糖体。通常由具分支的小管或小泡相互吻合而成,扁平囊少见。在同一细胞中,小管的管径比较一致,如在肝细胞中约为100nm,睾丸间质细胞中约为30~40nm。滑面内质网可与粗面内质网和高尔基复合体相连续,它也是一种可随细胞的类型和生理功能状态而变化的细胞器,其膜的化学组成、酶的种类和含量,以及大分子结构等都可能存在差异,因此其功能也各异。
功能
参与合成类固醇激素 睾丸间质细胞、卵巢黄体细胞和肾上腺皮质细胞都是分泌类固醇激素的内分泌细胞。类固醇激素的生物合成是以胆固醇为原料,这类细胞的滑面内质网膜上含有合成胆固醇所必需的酶,还含有将孕烯酮转变为雄激素、雌激素和孕激素所必需的酶系,以及将其转变为11-脱氧皮质醇的酶系,后者进入线粒体经羟化生成氢化可的松。
参与脂类代谢 滑面内质网是细胞脂类合成的主要部位,在脂类运输中也起重要作用。如肝细胞的滑面内质网能将从血液吸收进来的脂肪酸进行β氧化以提供能量,或将其脂化成为三酸甘油脂,一部分以脂滴形式储存于细胞质,大部分则组成脂蛋白复合物,经高尔基复合体,排入狄氏隙。
解毒作用 某些脂溶性药物和代谢产物具有一定的毒性,且不易直接排出。有些细胞的滑面内质网膜上具有进行氧化、还原、水解及结合等生物转化过程所必需的各种酶系,能使其毒性降低、失毒,或将其转化成为易溶于水的物质以利于排出。如肝细胞滑面内质网膜上的葡萄糖醛酸转移酶可使脂溶性巴比妥类药物与葡萄糖醛酸结合,形成水溶性物质; 滑面内质网膜上的酶能将氨基酸代谢过程中产生的有毒的氨转变为无毒的尿素。
参与壁细胞内盐酸的形成 壁细胞的细胞内分泌小管周围分布有大量管泡状滑面内质网,能将血浆中的Cl-传递到细胞内分泌小管,细胞质中从碳酸解离出来的H+也被输送至该处,二者结合遂生成盐酸排出细胞外。有人认为壁细胞的滑面内质网还可能具有浓缩Cl-的作用。
参与横纹肌的收缩活动 骨骼肌和心肌细胞的肌浆网是一种特化的滑面内质网,其膜上镶嵌有大量钙泵蛋白质,在Ca2+和Mg2+存在下,可将肌浆内的Ca2+泵入肌浆网,使肌浆内的Ca2+浓度降低。当神经冲动到达肌纤维膜,引起膜的去极化现象,去极化波沿膜表面传至T小管时,T小管则通过某种机制传给邻近的肌浆网,使其将Ca2+释放到肌浆,从而激活ATP酶,释放出能量,引起肌肉收缩。神经冲动结束,Ca2+又被泵回到肌浆网中,肌纤维松弛。因此,可以认为滑面内质网在肌细胞中起着调节肌浆内钙离子浓度,从而控制肌肉运动的作用。
参与血小板形成 巨核细胞的滑面内质网非常丰富,能将细胞质分隔成许多小区,内含血小板颗粒。随着巨核细胞逐渐发育成熟,滑面内质网膜不断扩展,相互连接,并与向内折入的细胞膜相连续,使每一细胞质小区都有完整的单位膜包围,最后从巨核细胞上脱落,形成游离的血小板。
与糖原代谢的关系 在肝细胞内,糖原颗粒常与滑面内质网伴存,提示滑面内质网可能与糖原的合成与分解有关。分析经分离的滑面内质网膜碎片时,发现碎片上含有葡萄糖-6-磷酸酶等与糖原代谢有关的酶类。但有人提出这些酶类原存在于细胞质基质,只是在样本制备过程中被沾染在碎片上,滑面内质网实际与糖原代谢无关。这有待进一步研究。
其他作用 滑面内质网在肝细胞中还参与生成胆汁,在视网膜色素细胞中能控制色素迁移,在幼稚的植物细胞中则可为细胞壁提供纤维素合成酶。此外,还有人认为滑面内质网与水和电解质代谢有关,在有丝分裂的末期则可能参与形成新的核被膜。
内质网与其他细胞器的关系 内质网与细胞膜、核被膜和高尔基复合体关系非常密切,而溶酶体、微体、圆球体和液泡等细胞器的形成都与内质网和高尔基复合体有关。一般认为内质网膜可能由细胞膜演化而来,核被膜则可能由内质网膜演化而来。因为在电镜下可以观察到细胞膜的凹陷部分与内质网膜直接相连续,还可见到细胞膜上有时也有附着核糖体,如哺乳动物的网织红细胞缺乏内质网,但仍可依靠附着在其细胞膜上的核糖体合成部分蛋白质。此种现象与原核细胞中核糖体附着于细胞膜及其内折物上合成蛋白质的情况极为相似,提示在细胞进化过程中,内质网有可能起源于细胞膜。电镜下,内质网膜与核被膜外层相连续,内质网池与核周隙直接相通,核被膜外层附有核糖体,其化学组成也与内质网膜明显相似,这些都反映二者在发生上的同源性。此外,在合成旺盛的细胞中,内质网常与线粒体紧密相伴,可能有利于线粒体为内质网进行功能活动提供能量。
内质网的发生 内质网是一种易变的动态结构,常随细胞的生理功能状态而不断生成和解体,其发生过程包括核糖体和内质网膜的形成,一般认为内质网膜起源于游离的多聚核糖体。有人在胚胎、再生和肿瘤细胞中观察到在游离的核糖体之间先出现许多小泡,小泡聚集在一起形成扁平的囊泡,再分化形成内质网。内质网膜上的酶蛋白可能是陆续组装入膜内,而不是一次同步合成。如刚出生的大鼠肝细胞的内质网中,葡萄糖-6-磷酸酶增加很快,一日间即达最高峰,而细胞色素P -450增加缓慢,须持续一周以上。退变的内质网可局部或全部通过细胞自噬作用形成自噬体,然后与溶酶体并合,并在其中被消化分解。
内质网endoplasmic reticulum
真核细胞内细胞器之一。是分布在细胞质内由膜围成的管腔道系统。包括囊状、管状和小泡状3种基本类型。膜为一层单位膜,管腔一般较窄,可随物质充盈而扩大。依其膜外面有无核糖体附着又可分为粗面内质网和滑面内质网两种。功能主要参与细胞内蛋白质、脂类的合成及其他一些重要的代谢活动,且作为内膜系统的主要组分参与膜的定向流动。
内质网
真核细胞的细胞质中折叠的膜系统。1945年由波特(K.R.Porter)发现。内质网膜的厚度约5~6nm。分粗面内质网(膜上附着大量核糖体者)和滑面内质网(膜上无核糖体附着者)2类。主要成分为脂(50%~60%为磷脂)、蛋白和少量RNA。
内质网neizhiwang
细胞质中由膜围成的分支小管、小囊或扁平囊状结构连通而成的管道系统,其周缘常分离出一种小泡状结构。电镜下观察,内质网膜厚度约为5~6微米,按形态结构的不同分为两个区域:一是粗面内质网,多为扁平囊状结构,膜上含有两种核糖体亲合蛋白,因而在膜的细胞质面上附着有核糖体;一是滑面内质网,多呈网状分布的小管,膜的细胞质面上不附着核糖体。滑面内质网不仅在一定部位与粗面内质网相连通,而且有的与质膜或核外膜相联。内质网扩大了细胞质内的膜面积,在内质网膜上附有的多种酶,为生命活动的各种化学反应的正常进行创造有利条件。粗面内质网不仅是核糖体的支架,而且是在核糖体上合成的分泌蛋白的运输通道。此外,能够对核糖体合成的多肽链进行一定的改造,或用于自身的装配和生成。滑面内质网具有解毒、合成脂类和分解糖元的功能,还参与分泌性蛋白的运输过程。
内质网endoplasmic reticulum, ER
系细胞器之一。广泛分布于细胞基质中,为膜性管道或囊泡系统。靠近细胞核处者可与核膜相通,靠近细胞膜处与细胞膜相通。故内质网是细胞质中互相连通的片层性管网结构。内质网有粗糙面内质网及平滑面内质网两种,前者表面上附有许多核蛋白体,后者表面没有。粗糙面内质网的功能与合成输出蛋白有关;平滑面内质网与肝细胞中糖原合成、精巢中雄性激素的合成与分泌有关。
内质网
存在于细胞质中的折叠状的膜系统。多数细胞中存在,在某些位置上,与细胞膜或核膜的外层相联。表面附着有核糖体的称为粗面内质网;无核糖体的称为光滑内质网。
内质网
是细胞质中的细胞器。除成熟的血红细胞外,内质网广泛存在于各种细胞内,尤以腺细胞(胰、肝)的内质网更为发达。内质网是由一系列相互连通的双层扁平囊状膜所组成,扁囊可局部膨大成池。内质网可分为滑面内质网和粗面内质网两种。粗面内质网是在小管或池的外表面附有核糖核酸小颗粒(核蛋白体),它与细胞内蛋白质的合成和运输有关。滑面内质网上没有核糖核酸附着,它与细胞内非蛋白质,如类脂、胆固醇、类固醇激素等的合成和分解有关。滑面内质网在骨骼肌细胞、肝细胞、肾上腺皮质细胞、卵巢黄体细胞、睾丸间质细胞内最为丰富。
内质网endoplasmic reticulum
是细胞质内一些互相通连着的膜管状结构,只有在电镜下才能看到。表面有核蛋白体附着者称粗面内质网,参与蛋白质的合成和转运,故在蛋白合成和分泌旺盛的细胞特别多,如浆细胞和胰腺细胞。表面无核蛋白体附着者称滑面内质网,参与糖、脂类和固醇类物质的转运和代谢过程。在横纹肌中的滑面内质网尚有摄取、贮存和释放钙离子的作用。
内质网
内质网是电镜下发现的一种膜性细胞器,是由扁囊(池)和小管互相通连而成的膜性囊管系统,几乎存在于所有细胞的胞质内,参与细胞内蛋白质、脂类的合成及其它一些重要的代谢活动。内质网的膜也属于单位膜,但比细胞膜薄,厚约5~6nm,膜上有在代谢过程中起重要作用的多种特殊的酶。内质网的腔一般较窄,可随物质充盈而扩大。内质网的多寡随细胞类型及功能状态而异。内质网可依其膜外面(即胞质面)有无核糖体附着分为粗面内质网和滑面内质网(图1)。在分泌蛋白功能旺盛的细胞内,粗面内质网特别丰富,而在分泌类固醇激素的细胞中,滑面内质网特别发达,这种区别反映出这两型内质网功能上的差异。光镜下,由于分辨力低而不能辨认内质网,但在H.E染色的标本上,粗面内质网及游离核糖体丰富的胞质区域呈较强的嗜碱性染色,而滑面内质网丰富区的胞质则显嗜酸性染色。
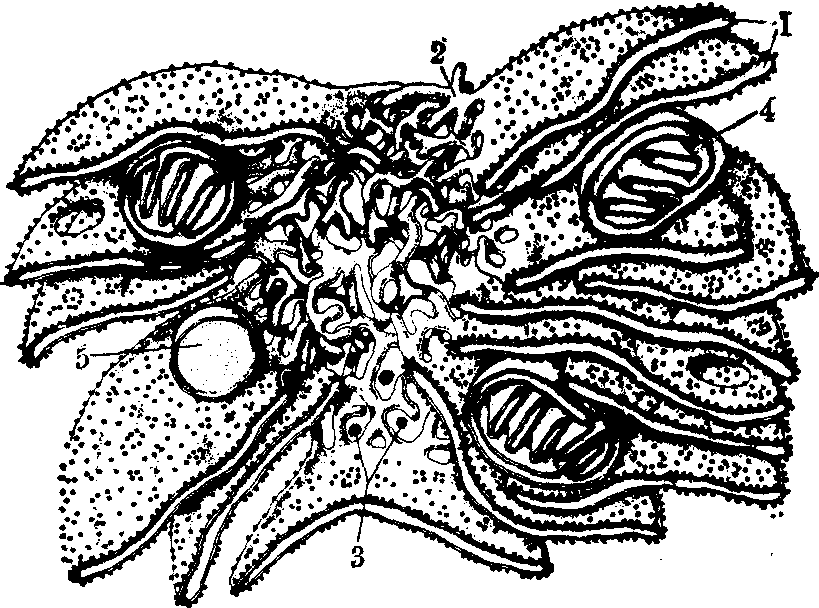
图1 内质网立体示意图
1.粗面内质网 2.滑面内质网 3.糖原颗粒4.线粒体 5.溶酶体
粗面内质网 也称颗粒内质网,一般成自一些平行排列的扁囊,扁囊互相通连,在膜外面附着许多核糖体(图2)。通常认为所附着的是多核糖体,它由一个mRNA与数个至30个以上的核糖体结合而成,通过各个核糖体大亚单位顶部与内质网膜上的受体蛋白结合而附于膜的外面。从膜的表面看,多核糖体排列成行、环或螺旋形(图1)。
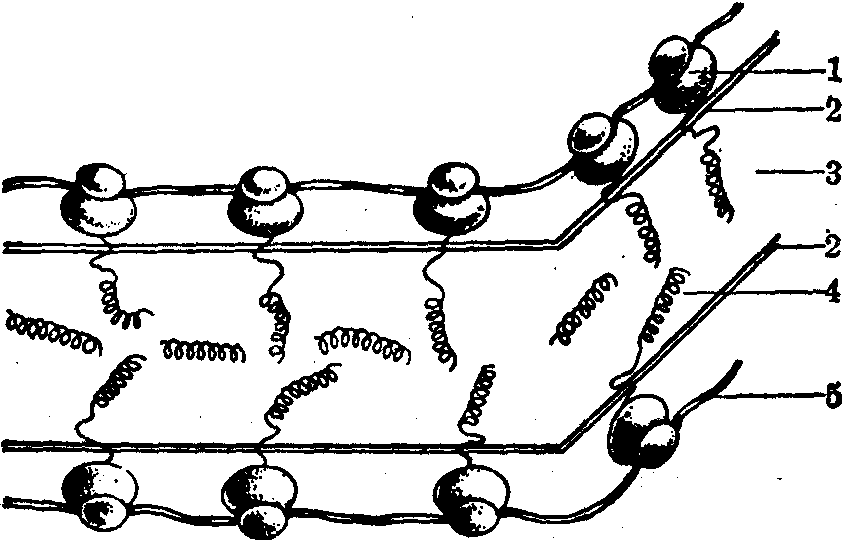
图2 粗面内质网与分泌蛋白合成示意图
1.核糖体以大亚单位附于内质网膜外面(胞质面) 2.粗面内质网的膜 3.粗面内质网的腔 4.形成中的多肽链 5.信使核糖核酸(mRNA)
在不同类型和不同功能状态的细胞内,粗面内质网的量差别很大,以分泌蛋白类旺盛的细胞最为丰富,如活跃的成纤维细胞和成骨细胞、浆细胞、胃底腺主细胞、胰和唾腺的浆液性细胞和肝细胞等。粗面内质网在胞质内的分布往往显出细胞类型的特征,如在胰腺泡细胞和胃底腺主细胞等外分泌腺细胞中,集中在细胞基部和核旁;于肝细胞成许多扁囊群,每群有3~20个平行的扁囊,扁囊群之间为滑面内质网区; 浆细胞的胞质则充满密集的粗面内质网,唯核附近的高尔基复合体区除外。
粗面内质网的功能主要是合成分泌蛋白。以mRNA为模板所形成的多肽链穿过内质网膜进入腔内,经过复杂的修饰、加工与构型,合成具有一定空间结构的蛋白质(图2);糖蛋白前体还在此经糖基转移酶的催化与核心糖基结合。合成的蛋白质输送到高尔基复合体附近,包在从扁囊脱落的运输小泡内,由运输小泡转运到高尔基扁平囊泡内,再经加工浓缩,形成分泌颗粒,移到细胞表面排出。外分泌腺细胞分泌蛋白的合成与分泌过程,已由放射自显影与电镜观察相结合的方法进行追踪研究,予以证实(参见“胰”条)。
粗面内质网还合成细胞自身所需的蛋白,如膜蛋白、溶酶体与微体的酶。有的资料表明,粗面内质网和滑面内质网均能合成脂类,参与脂蛋白合成。
滑面内质网 也称无粒内质网,它虽与粗面内质网通连,但有独特的形态,一般为一些形状及直径不一的小管,互相通连成网状,其特点是小管膜的外面光滑,不附有核糖体(图1)。多种细胞的滑面内质网不发达,分布形式各异。类固醇激素分泌细胞则有丰富的滑面内质网,遍布在胞质内。肝细胞滑面内质网也丰富,与肝的许多重要物质代谢有关。横纹肌纤维内,滑面内质网呈特异的形态,称肌质网。
滑面内质网的功能比较复杂,随所在细胞种类而有很大差别,概括如下:
❶小肠吸收细胞和肝细胞的滑面内质网参与脂类的合成与运输,在大多数细胞中也是脂类合成的主要部位(参见“小肠”条,“肝细胞”条);
❷类固醇激素分泌细胞的滑面内质网参与激素的合成,合成过程所需的酶大多位于滑面内质网,少数在线粒体内,故激素的合成是在滑面内质网与线粒体协同参与下进行;
❸肝细胞的滑面内质网膜上有规律地排列着多种酶系如氧化还原酶系、水解酶、合成酶系等,参与胆汁合成、激素代谢及由肠道吸收的有机异物(药物、腐败产物)的生物转化(参见“肝细胞”条);
❹肝细胞中糖原颗粒常与滑面内质网伴存,一般认为滑面内质网参与糖原分解代谢(参见“肝细胞”条);
❺横纹肌纤维的肌质网是特殊形式的滑面内质网,它的膜上有钙泵蛋白等蛋白质,对肌浆内钙离子浓度有调节作用,与传导神经兴奋引起肌纤维收缩有关(参见“骨胳肌”条);
❻骨髓巨核细胞的滑面内质网将胞质分隔成许多小块,形成血小板(参见“血细胞发生”条)。微粒体 微粒体不同于微体,不是细胞固有的细胞器,而是将细胞制备成匀浆时,被打碎的内质网片段愈合成的小泡。微粒体可经梯度离心分离出来,对它进行化学成分分析和电镜观察,以研究内质网的成分和功能。电镜下,微粒体分为粗面和滑面两种,分别来自粗面内质网和滑面内质网。
有孔板层 电镜下,在某些细胞的胞质中见多个平行并列或单个的有孔膜扁囊,多位于细胞核附近,常见于分裂活跃的细胞,如胚胎的细胞、肿瘤细胞及男性和女性的生殖细胞。膜厚7~9nm,两膜间的腔宽30~50nm。孔的直径40~50nm,配布较规律,一般相距100~200nm。孔的结构与核被膜的相似,两层膜在孔边缘处相连,也有隔膜封闭。有孔板层的功能还未了解,或许是将RNA由细胞核输送到细胞质。它们可能由核被膜或内质网发生。
核外染色质 在染色的光镜标本上,多种细胞的胞质含有对碱性染料亲和性强的物质,或弥散分布在胞质内(如淋巴细胞、浆细胞),或集聚成团块状(如神经元、肝细胞),或集中在细胞基部(如胰腺泡细胞)。早年的细胞学家把这种深染的嗜碱性物质称为核外染色质。经组织化学染色及紫外线吸收光谱方法鉴定表明,核外染色质含核糖核酸。电镜下证明,核外染色质是富于游离核糖体和(或)粗面内质网的区域。核外染色质随细胞的功能活动而改变,故早年又称为动质。
内质网
内质网是真核细胞的重要细胞器之一,它是细胞一系列重要物质,如蛋白质(包括酶、抗体与激素等)、糖类与脂类的合成基地。内质网广泛存在于绝大多数动植物细胞内,甚至在原生动物与真菌(如酵母菌)等细胞内亦能观察到较为简单的内质网结构。只有在原核细胞 (支原体、细菌与蓝藻等)与少数高度分化的真核细胞 (如无核红细胞) 内没有内质网。在原核细胞内由细胞膜代行内质网的主要功能。
内质网作为一个独立的细胞器,它的发现比线粒体与高尔基体等要晚得多,因为只有在电子显微镜下才能观察到内质网的真正结构。在光学显微镜下,虽然早就描述过在细胞质内的碱性染料区域与蛋白质合成有关,在研究原生质的特性时,有人提出过动质的概念,但只有到40—50年代,由于电子显微镜与超薄切片技术相结合才有可能真正揭示内质网这一重要细胞器。1952年Porter等人在鸡胚成纤维细胞内初次观察到细胞质的内质部位分布有网状结构,因此建议叫做内质网。虽然以后发现内质网不尽在内质部位,但仍习惯沿用此名称。
内质网由膜和膜围成的腔(或称囊池) 组成,内质网的膜具有一般生物膜的共同特点,膜厚约5—6nm,在高倍镜放大下内质网膜又可分为三层。膜的基本成分是脂蛋白,脂类成分可达50%,其中磷脂类约占一半以上,尤以卵磷脂与脑磷脂含量最多,内质网膜上还含有各种酶与一定量的RNA。
内质网的结构 在不同类型的细胞,甚至相同类型细胞的不同生理状态下,内质网呈多形态性。内质网大致可分为三种基本形态:
❶扁囊状内质网:这种内质网中最常见的是整齐排列的扁囊内质网,普遍存在于各种分泌细胞内,在超薄切片上可以观察到两个平行的膜之间有狭窄的腔。在特殊生理条件下或病理状态下,扁囊内质网可以变成扩张内质网,其腔很宽,甚至呈大泡状。
❷小泡状内质网:其立体构型近似小球状,一般认为这是一种过渡型内质网。
❸管道状内质网: 往往由分支状管道形成的较为复杂的立体构型。有人认为即使是在正常生理状态下,内质网的结构也是不稳定的。内质网的扁囊与管道可以随时缢裂成很多小泡。在细胞周期的各个阶段,内质网的变化是极为复杂的,在细胞分裂时内质网要经历解体与重建的过程。
内质网的类型和功能 按基本功能与结构,内质网分为两大类型:粗面内质网和滑面内质网。
粗面内质网 或称颗粒内质网,是内质网与核糖体共同形成的复合功能结构,其主要功能是合成分子量较大的、分泌性的蛋白质,如酶原蛋白、抗体蛋白与激素蛋白等。在分泌细胞与浆细胞内,粗面内质网较为发达,而在一些未分化的细胞与肿瘤细胞内,粗面内质网较为稀少。粗面内质网主要是扁囊状内质网。粗面内质网是因为在其膜表面分布着大量颗粒结构(即核糖体)故名。目前尚未观察到核糖体与膜之间具有直接的结构联系,用核糖核酸酶处理,核糖体很快降解与消失。核糖体在膜表面的分布排列是多样性的,如胰腺外分泌细胞分布均匀整齐,肝细胞的不整齐,浆细胞与成纤维细胞常常是数个颗粒排成环状,有的细胞也有排成花瓣状的,这显然是与多聚核糖体的形态特点有关系。一般认为粗面内质网上合成的蛋白质首先进入内质网的腔内,再由管道输送到高尔基体进行浓缩与包装后分泌到细胞之外。
滑面内质网 与脂类及糖类的合成关系密切。滑面内质网基本是管状内质网。在糖类与脂类代谢旺盛的细胞内,以及色素细胞内滑面内质网转为发达。植物细胞的滑面内质网与壁的形成有关,正在形成壁的细胞内,滑面内质网在胞质外层较为丰富。肝细胞中的滑面内质网分布在肝糖合成的区域,这显然与肝糖的贮存有关系,但至今尚未发现滑面内质网中有糖原合成的酶。在合成脂肪类或固醇类的细胞内,如脂肪细胞,肾上腺皮质细胞,以及植物的胚乳细胞内,滑面内质网较发达,这些均说明它与脂类代谢关系十分密切。
内质网的其他功能 内质网的功能是多方面的,除以上所述的重要合成功能外。其扁囊与管道具有积累与运输合成物质的作用,内质网合成的物质通过管道直接输送到细胞质的各部位,而主要是送到高尔基体上“加工”。内质网与吸收和输入物质也有关系,内质网的囊与管道经常和吞饮泡与吞噬泡直接连结。此外,内质网可能还有传递能量,运动与支架等作用。内质网对外界因素的作用(如放射线与化学毒物的作用,病毒感染等)非常敏感,粗面内质网发生的最明显与最普遍的病理变化是内质网腔扩大,并形成空泡,继而发生脱粒现象,即核糖体从内质网膜上脱落与消失,说明蛋白质合成受阻。滑面内质网在激素与药物的作用下常常有增生现象,这是一种病理现象。
内质网与其他细胞器的关系 内质网与其他细胞器关系的研究对阐明细胞内一些生理生化过程以及细胞器的发生与进化是很有意义的,在这一方面已积累了很多资料,也提出了一些有启示性的设想。超微结构研究说明,向内折叠的细胞膜有时与内质网相连结,甚至管道相通。人们考虑的不仅是内质网与细胞在功能上的相关性,更进一步提出原核细胞内虽然没有内质网,但细胞膜却经常向内折叠,伸向胞质深处,形成与内质网颇类似的管道或囊。原核细胞膜内侧有时附着大量核糖体,并兼顾执行着粗面内质网合成蛋白质的功能,从而一些人认为,在细胞进化过程中,内质网可能是细胞膜进一步分化演变而来。在超薄切片上可以经常观察到内质网的膜经常与核膜外层连结,内质网的腔和管道与双层核膜相通,这种结构上的关系明显反映核质交换的关系,这经常发生在代谢极为旺盛的细胞内。而且核膜外层有时也附着大量的核糖体,显然起着粗面内质网的类似作用。由此也就逻辑地提出内质网与核被膜在发生上的同源关系,在进化过程中核被膜可能是由内质网膜演化而来。内质网与高尔基体在结构、功能与发生上的关系更为密切。内质网合成的物质是通过内质网的管道输送到高尔基体的。最常见的形式是充满合成物的内质网管道缢成大量小泡。移向高尔基体的形成面,将合成物注入高尔基体扁囊 甚至可以观察到小泡与高尔基体囊融合。有人也描述过滑面内质网管道直接与高尔基体连通的现象(见图)。

粗面内质网、滑面内质网与高尔基体的关系
滑面内质网与高尔基体在发生与进化上同源性的假设可能易被人们所接受。在合成功能旺盛的细胞内,内质网总是与线粒体紧密相依,因为线粒体是内质网执行功能的能量直接“供应站”。内质网
endoplasmic reticulum(ER)
- 议案的审议是什么意思
- 议案的审议是什么意思
- 议案的审议是什么意思
- 议案的审议是什么意思
- 议案的审议是什么意思
- 议案的审议是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思
- 议案的提出和审议是什么意思