免疫球蛋白分子的遗传标志和血清学类型
免疫球蛋白分子的遗传标志和血清学类型
免疫球蛋白分子具有不同的抗原结合部位,可与相应的抗原发生特异性结合,但其本身又可作为抗原被其特异性抗血清所检出。决定免疫球蛋白分子抗原特异性的化学基因称为抗原决定簇,也称抗原决定基。抗原决定簇的差异,表现在免疫球蛋白分子在血清学上的不均一性,反映了免疫球蛋白分子氨基酸组成和序列的变化。由于免疫球蛋白分子是受基因控制的,因而抗原决定簇也反映了抗体形成细胞遗传性的差异。这种受遗传控制的免疫球蛋白分子上抗原决定簇分子结构上的差异,就称为免疫球蛋白分子的遗传标志。根据不同的遗传标志,可将免疫球蛋白分子的抗原决定簇分为同种型、异型和独特型三个不同的血清学类型。
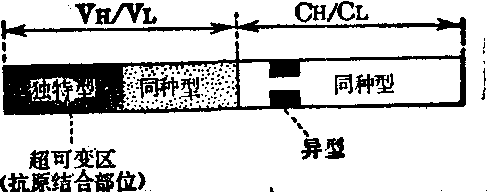
免疫球蛋白分子各种变异型别的抗原决定簇所在位置示意图
同种型 是指同一物种内所有个体都具有的免疫球蛋白的抗原特异性。根据重链或轻链恒定区以及重链和轻链可变区结构的变化分为不同的类、亚类、型、亚型、群和亚群的免疫球蛋白变体。所有同种型变体均存在于每个正常个体的血清中。
类: 根据重链恒定区(CH)的化学性质、分子量、单体数、糖含量及电泳移动率等的特点,将重链特异性抗原分为五类,分别为γ(gamma)、α(alpha)、μ(mu)、δ(delta)及ε (epsilon),其相应免疫球蛋白分别为IgG、IgA、IgM、IgD和IgE。
亚类: 进一步根据各类免疫球蛋白重链恒定区的抗原特异性的差异以及二硫键位置和数目的不同,某类免疫球蛋白再分为亚类。如IgG可分为IgG1、IgG2、IgG3和IgG4四个亚类;IgA可分为IgA1和IgA2两个亚类;IgM可分为IgM1和IgM2两个亚类。
型: 根据各类免疫球蛋白轻链恒定区的抗原特异性分为两型,即κ型和λ型。任何一个免疫球蛋白分子的二条轻链总是同型的,不会同时存在这两型。但在正常人血清中,κ型和λ型免疫球蛋白分子都是存在的,两者的比例约为2:1。每类免疫球蛋白分子中,κ型和λ型的比例也大致如此。
亚型: 根据免疫球蛋白分子λ链恒定区氨基酸序列上的某些变化而将λ链分为四个亚型,即Cλ1(Oz+、Kern-、Mcg-)、Cλ2 (Oz-、Kern+、Mcg+)、Cλ3(Oz-、Kern+、Mcg-)和Cλ4 (Oz-、Kern-、Mcg-)。这几个亚型都是从病人中发现的,Oz、Kern、Mcg等都是病人名字的缩写。Oz+代表氨基酸序列193位置上为赖氨酸,Oz-为精氨酸; Kern+代表156位置上为甘氨酸,Kern-为丝氨酸: Mcg+分别代表116/118/167位置上为天门冬酰胺/苏氨酸/赖氨酸,Mcg-则为丙氨酸/丝氨酸/苏氨酸。K链尚未发现有亚型。
免疫球蛋白分子的遗传标志和血清学类型
| 类 型 | 标志部位 | 特 点 | 举 例 | 分 布 |
| 同种型 | 全部变体存在于正 常个体血清中 | |||
| 类 亚类 | CH CH | 理化性状 细微结构 | IgG,IgA,IgM,IgD,IgE IgG1, IgG2, IgG3, IgG4 IgA1, IgA2 IgM1, IgM2 | |
| 型 亚型 | CL CL | 结构特点 氨基酸种类 | κ,λ Cλ1(Oz+、Kern-、Mcg-) Cλ2(Oz-、Kern+、Mcg+) Cλ3(Oz-、 Kern+、Mcg-) Cλ4(Oz-、Kern-、Mcg-) | |
| 群 亚群 | VH/VL VH VK Vλ | 结构特点 | VH,Vκ,Vλ VHⅠ,VHⅡ, VHⅢ, VHⅣ VκⅠ,VκⅡ,VκⅢ,VκⅣ VλⅠ,VλⅡ,VλⅢ,VλⅣ,VλⅤ,VλⅥ | |
| 异型 | CH(γ1) CH(γ2) CH(γ3) CH(α2) CL(κ) | G1m 1, 2, 3,17 G2m 23 G3m 5, 6,10,11,13,14,15,16, 21, 24, 26, 27 Azm 1, 2 Km(InV) 1,2,3 | 交替形式:遗传控 制并不出现在全 部个体中 | |
| 独特型 | VH/VL | 氨基酸序列 | 可能在形成抗原结合部位的一个或一个以 上高变区 | 每一免疫球蛋白分 子具有个体特性 |
群和亚群: 免疫球蛋白的可变区是非常不均一的。到目前为止,已分析了上千例多发型骨髓瘤病人的IgG,没有发现任何两例IgG的可变区的氨基酸组成及顺序是完全相同的。虽然如此,可变区仍可根据其同源性程度,分为三个主要的群,即VH群、VK群和Vλ群。比较不同来源各群可变区氨基酸的组成和排列顺序,可进一步将各群分为若干亚群。无论哪一类免疫球蛋白的重链可变区都可分为VHI、VHII、VHIII和VHIV四个亚群;k型轻链可变区可分为VKI、VKII、VKIII及VKIV等亚群; λ型轻链可变区可分为VλI、VλII、VλIII、VλIV、VλV及VλVI等亚群。各亚群在每个正常个体中都同时存在。
异型 也称同种异型,是指在同种某些个体共同存在的抗原决定簇。即具有某个抗原决定簇的免疫球蛋白可存在于某人群中,但不同抗原决定簇的异型免疫球蛋白不能同时存在于一个个体内。正如人的血型物质(A、B、O) 不能同时存在于一个个体内一样。异型免疫球蛋白的区别有时只相差一个氨基酸,这个起决定作用的氨基酸称为异型标志。目前已证明在人有4组异型标志,为二组独立的基因连锁群所编码,其一为含有Gm、Am、Mm的连锁群,另一为含Km的连锁群。Gm、Am、Mm因子分别为IgG、IgA和IgM重链上的遗传标志,Km(旧称InV)因子为轻链上的遗传标志。在m前加数字表示该标志与免疫球蛋白亚类的明确关系,而异型标志则以m后用数字加圆括号表示。如IgG1的异型标志有G1m(1)、G1m(2)、G1m(17) ……等; IgA2的异型标志有A2m(1)、A2m(2)等。Gm因子定位于γ链的恒定区,多存在于Fc分段,但G1m(4)、G1m(17)则存在于Fd分段。IgG约有30个遗传标志,有些Gm因子与IgG亚类的关系已明确,如G1m、G2m、G3m。但IgG4尚未发现有异型标志。Am因子是存在于α链上的异型标志,IgA1尚未发现有遗传标记,IgA2有两个标志,即A2m(1)、A2m(2)。必须指出,重链的一个异型标志只存在于一个亚类的免疫球蛋白中,也就是说一个亚类只表现一个Gm因子,不能同时存在两个因子。Km因子存在于k型的轻链上,为各类免疫球蛋白所共有。Km因子有三个异型标志。λ型尚未发现有异型标志。独特型 也称个体基因型。每一抗体形成克隆所产生的免疫球蛋白分子特有的抗原性称为此免疫球蛋白分子的独特型。此抗原特异性定位于免疫球蛋白的可变区,即抗原的结合部位,与高变区相关,亦即存在于轻链及重链可变区的与抗原决定簇结合的特殊部位(参见“免疫球蛋白”)。其单一决定簇称为独特型抗原决定簇。每一免疫球蛋白分子可变区可有多个独特型抗原决定簇,而其总和则称为免疫球蛋白的独特型。
抗体多样性的遗传控制 机体免疫系统为什么能产生千万种不同的免疫球蛋白分子,即抗体多样性或不均一性是如何形成的,已成为免疫学最重要的课题之一,它涉及一些十分独特的遗传学问题。目前解释抗体多样性的理论是根据免疫球蛋白分子结构和氨基酸序列分析的资料。通过对鼠M蛋白轻链结构的研究,表明免疫球蛋白肽链是由2个不同基因分别编码可变区和恒定区,因此提出2个基因编码一条肽链的学说。认为恒定区基因(C基因)是有限的,它虽可决定免疫球蛋白分子的类别和亚类,是造成免疫球蛋白分子多样性的原因之一,但造成免疫球蛋白分子多样性的主要原因是由可变区的异质性决定的,它是由可变区基因(V基因)编码的。V基因数目仍不清楚。
基于上述认识,对抗体多样性的遗传控制曾提出三种学说。其一为胚系学说又称种系学说,认为抗体形成细胞具有编码免疫球蛋白分子的全部基因 (即有限数量的C基因和未知数量的V基因),通过长期进化形成的,并通过生殖细胞从亲代传给子代。但现有资料提示,在生殖细胞内不可能存在如此大量的V基因。另一学说为体细胞突变学说,认为在生殖细胞内只继承了数量有限的V基因,但免疫球蛋白分子多样性的形成是由于体细胞在发育过程中发生突变或基因重组,因而产生许多不同的V基因。根据小鼠分子杂交的研究表明,V基因的数量可能很少,因而提示体细胞突变可能对免疫球蛋白分子多样性起重要作用。近年来对鼠轻链mRNA分离纯化成功以及体外核酸分子杂交技术的建立,可以直接研究免疫球蛋白分子的遗传学。根据对鼠DNA片段研究表明,免疫球蛋白分子可变区是由一个V基因片段、J基因片段及D基因片段组成,因此V、D、J基因相互连接对免疫球蛋白分子多样性的产生是极为重要的,此即所谓V区基因相互作用学说。
总之,从对免疫球蛋白分子结构和DNA结构的研究表明,免疫球蛋白分子多样性不可能简单地归因于胚系基因、体细胞突变或基因相互作用单一机制加以说明,它可能与上述多种机制有关。此外,还依赖于基因片段连接点上的连接多样性以及VH和VL链的不同配对有关。
☚ HLA复合体 免疫化学 ☛
- 宥是什么意思
- 宥十世是什么意思
- 宥坐之器,虚则欹,中则正,满则覆。是什么意思
- 宥密是什么意思
- 宥情是什么意思
- 宦是什么意思
- 宦是什么意思
- 宦是什么意思
- 宦乡是什么意思
- 宦乡是什么意思
- 宦娘是什么意思
- 宦学吏师制确立是什么意思
- 宦官是什么意思
- 宦官是什么意思
- 宦官是什么意思
- 宦官是什么意思
- 宦官是什么意思
- 宦官弄权是什么意思
- 宦海是什么意思
- 宦海是什么意思
- 宦海升沉录是什么意思
- 宦海升沉录是什么意思
- 宦海日记校注是什么意思
- 宦海潮是什么意思
- 宦海风波是什么意思
- 宦游纪闻是什么意思
- 宦者是什么意思
- 宦门子弟错立身是什么意思
- 宦门子弟错立身是什么意思
- 宦门子弟错立身是什么意思
- 宦门子弟错立身是什么意思
- 宦顺源是什么意思
- 宪是什么意思
- 宪是什么意思
- 宪世编是什么意思
- 宪之魂是什么意思
- 宪之魂是什么意思
- 宪兵是什么意思
- 宪友会湖南支部是什么意思
- 宪台通纪是什么意思
- 宪台通纪续集是什么意思
- 宪圣皇后(吴氏)是什么意思
- 宪宗七年(公元1257年)是什么意思
- 宪宗三年(公元1253年)是什么意思
- 宪宗二年(公元1252年)是什么意思
- 宪宗元年(公元1251年)是什么意思
- 宪宗六年(公元1256年)是什么意思
- 宪宗(段正严)是什么意思
- 宪德是什么意思
- 宪成皇后(野利氏)是什么意思
- 宪政是什么意思
- 宪政是什么意思
- 宪政促进会是什么意思
- 宪政促进会筹备会是什么意思
- 宪政党是什么意思
- 宪政党是什么意思
- 宪政公会是什么意思
- 宪政商榷会是什么意思
- 宪政实施促进委员会是什么意思
- 宪政实施协进会是什么意思