免疫球蛋白与遗传
免疫球蛋白是一类有抗体活性的糖蛋白分子,是由B淋巴细胞分化而来的浆细胞产生的。免疫球蛋白分子有高度多样性,其遗传机理是遗传学上的一个难题。近年来应用基因克隆增殖技术和核苷酸顺序分析研究免疫球蛋白的基因结构,揭示了免疫球蛋白多样性的遗传机理,并使人们对基因有了新的认识。
免疫球蛋白的基本结构 是由两条相同的轻链(L)和两条相同的重链(H)构成的四聚体。不同的免疫球蛋白分子,其氨基酸顺序,在轻链氨基端1/2部分和重链氨基端1/4部分间有较大变化,分别称为轻链可变区(VL)和重链可变区(VH)。可变区中又有几段氨基酸顺序特别多变,称为高变区。VL有三个高变区,VH有四个高变区。轻链羧基端1/2部分和重链羧基端3/4部分的氨基酸顺序也有变化,但其变化程度较低,分别称为轻链恒定区(CL)和重链恒定区 (CH)。两条轻链分别由二硫键连接在两条重链的氨基端,两条重链约在中间部分再由若干二硫链连接起来,构成免疫球蛋白分子。
免疫球蛋白的同种型 人或高等动物能够产生种类很多的免疫球蛋白分子。这些免疫球蛋白分子可按其血清学特性划分成不同类型。这些类型为同一个体所产生,称为免疫球蛋白的同种型。
重链恒定区的类和亚类 根据CH的抗原特异性可把重链分为五类,分别称为γ、 α、 μ、 δ和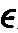 。 由此构成五种免疫球蛋白,分别称为IgG、IgA、IgM、IgD以及IgE。γ重链有γ1、γ2、γ3、γ4四个亚类,构成IgG1、IgG2、IgG3和IgG4。α重链有α1、α2两个亚类,构成IgA1和IgA2。μ重链有μ1和μ2两个亚类,构成IgM1和IgM2。 δ和
。 由此构成五种免疫球蛋白,分别称为IgG、IgA、IgM、IgD以及IgE。γ重链有γ1、γ2、γ3、γ4四个亚类,构成IgG1、IgG2、IgG3和IgG4。α重链有α1、α2两个亚类,构成IgA1和IgA2。μ重链有μ1和μ2两个亚类,构成IgM1和IgM2。 δ和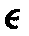 未发现亚类。
未发现亚类。
轻链恒定区的型和亚型 根据CL的抗原特异性,轻链有κ和λ两型。正常人血清中的免疫球蛋白分子约65%含有κ链,35%含有λ链。一个免疫球蛋白分子或含两条κ链,或含两条λ链。λ链有λ1、λ2、λ3三种亚型。轻链和重链可变区的组与亚组 可变区有κ链可变区(V )、λ链可变区(Vλ)和重链可变区(VH)三组。尽管这些可变区的氨基酸顺序多变,仍可根据它们的异同程度划分为若干亚组。Vx可分VxⅠ、VxⅡ、VχⅢ三个亚组。Vλ有VλⅠ、VλⅡ、VλⅢ、VλⅣ、VλV五个亚组。VH有VHⅠ、VHⅡ、VHⅢ三个亚组,可能还有VHⅣ。属于同一亚组的可变区仍有很多变异,但比不同亚组的可变区更为相似。
)、λ链可变区(Vλ)和重链可变区(VH)三组。尽管这些可变区的氨基酸顺序多变,仍可根据它们的异同程度划分为若干亚组。Vx可分VxⅠ、VxⅡ、VχⅢ三个亚组。Vλ有VλⅠ、VλⅡ、VλⅢ、VλⅣ、VλV五个亚组。VH有VHⅠ、VHⅡ、VHⅢ三个亚组,可能还有VHⅣ。属于同一亚组的可变区仍有很多变异,但比不同亚组的可变区更为相似。
免疫球蛋白的同种异型 免疫球蛋白分子在种内存在差异,这差异是由遗传决定的,称为同种异型。同种异型特异性可由血凝抑制试验检出。带有一定Rh抗原的红细胞,以已知同种异型特异性的不完全Rh抗体处理,在加入针对此同种异型特异性的抗血清时,将使红细胞发生凝集。这是一种抗人球蛋白试验。如在抗血清中加入待检血清,当此待检血清的同种异型特异性与上述不完全Rh抗体相同时,待检血清将竞争同种异型特异性抗体而抑制红细胞凝集。这时即可鉴定待检血清的同种异型特异性与上述不完全Rh抗体的同种异型特异性相同。Gm系统是γ重链恒定区的遗传标记,至少有23种异型(Gm1~23)。Am系统是α重链的遗传标记,有Am1和Am2两种亚型。Km系统(旧称InV系统)是κ轻链恒定区的遗传标记,有Km(1)、Km(1,2)、Km (3)三种异型。λ轻链的遗传标记有OZ、MZ、hern、Mcg等系统。有些同种异型的氨基酸置换已被揭示。如γ1重链第214位氨基酸,在Gm4型是精氨酸,在Gm17型是赖氨酸。又如κ轻链第153位和第191位氨基酸,在Km(1)型分别是缬氨酸和亮氨酸,在Km(1,2)分别是丙氨酸和亮氨酸,在Km(3)分别是丙氨酸和缬氨酸。免疫球蛋白的同种异型呈孟德尔式遗传。
免疫球蛋白的独特型 Oudin(1963)以沙门杆菌疫苗免疫50只兔子。所有兔子都产生了抗沙门杆菌抗体。从1号兔血清中分离出此抗体,以此免疫另一兔子,得沙门杆菌抗体的抗抗体。此抗抗体只与1号兔沙门杆菌抗体发生沉淀反应,但不与1号兔以沙门杆菌疫苗免疫前的血清反应,不与1号兔针对其他抗原的抗体反应,也不与其他49只兔子中任何一只兔子的抗沙门杆菌抗体发生反应。这一实验说明,50只兔子产生的沙门杆菌抗体都是独特的。针对半抗原抗体的独特型抗体可为该半抗原所抑制,这提示免疫球蛋白独特型的抗原决定簇在它的抗原结合部位或其附近。可以设想,免疫球蛋白独特型的贮备量与其抗原结合部位的贮备量一样大。
免疫球蛋白的遗传机理 浆细胞合成免疫球蛋白,首先以相应mRNA为模板合成轻链和重链。免疫球蛋白轻链和重链mRNA的基本结构与其他蛋白质肽链的mRNA相同,有连续的核苷酸顺序为可变区和恒定区编码。免疫球蛋白轻链或重链的特殊结构提示它们有独特的遗传机理。早在1965年,Dreyer和Bennett就设想,为轻链可变区和恒定区编码的DNA顺序是不连续的。七十年代初以来,随着分子生物学和遗传工程的发展,应用分子杂交、DNA建株和核苷酸顺序分析等技术,证明Dreyer和Bennett的设想基本正确,从而揭示了通过体细胞重组和体细胞突变导致免疫球蛋白多样性的遗传机理。
免疫球蛋白轻链的遗传控制 现知人的κ轻链基因在第2号染色体短臂,λ轻链基因在第22号染色体长臂。κ轻链基因及其表达如图1所示。在种系细胞和胚胎细胞中有大量的Vκ顺序,每一个Vκ顺序为κ轻链可变区氨基端95个氨基酸编码。这些Vκ顺序似可分为若干族,每族的核苷酸顺序密切相关。在每个Vκ顺序的上游有一个Lκ顺序(L意为导引),为κ轻链氨基端的嫌水性导引肽段编码。两者之间有一个短的间隔顺序。在最后一个Vκ顺序的下游有五个Jκ顺序 (J意为连接),为可变区的其余13个氨基酸编码,相互间也有间隔顺序隔开。在最后一个Jκ顺序的下游有单一Cκ顺序,为恒定区编码。在淋巴细胞分化过程中首先发生体细胞重组,大量Vκ顺序中的一个和五个Jκ顺序中的一个 (随同其下游Cκ顺序)相接,形成连续的Vκ/Jκ顺序,是为Vκ/Jκ重组。重组后的DNA成为有转录活性的基因。原初转录产物在Lκ顺序和Vκ/Jκ顺序之间、Vκ/Jκ顺序和Cκ顺序之间仍有间隔序列。它们大概在酶的作用下发生拼接,去除间隔顺序,形成Lκ/Vκ/Jκ/Cκ相接的成熟mRNA,继而以此mRNA为模板合成κ轻链前体。前体在其氨基端有导引肽段,在抗体分子通过细胞时被切去。此导引肽段大概有利于抗体分子通过细胞膜(图1)。
据Leder(1982)估计,保守地说Vκ顺序有150个,Jκ顺序有5个,Cκ顺序只有1个。可重组产生750种不同组合。此外,在V/J重组时,其接头处可有灵活性,至少可使组合可能性增加10倍。总共可产生7500种κ轻
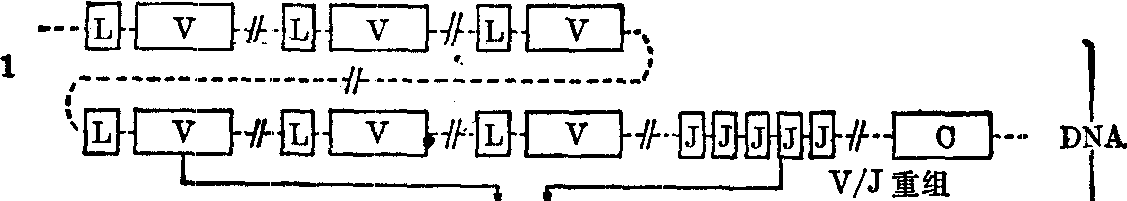
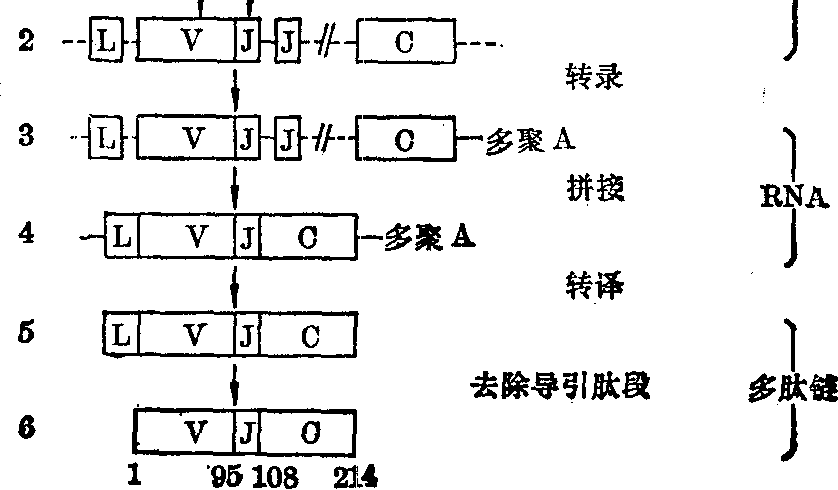
图1 人κ轻链活性基因的形成及其表达
1.种系细胞和胚胎细胞中的κ轻链遗传结构 2.体细胞重组(V/J重组)后形成的活性基因 3. κ轻链原初转录产物 4. κ轻链mRNA 5. κ轻链前体 6. κ轻链
λ轻链的多样性低于κ轻链,但也有很多Vλ顺序。小鼠有4个Cλ顺序,各伴随一个Jλ顺序。人有6个Cλ顺序,也各伴随一个Jλ顺序。
免疫球蛋白重链的遗传控制 人免疫球蛋白的重链基因在第14号染色体长臂。据目前估计,人约有80个VH顺序,在每个VH上游有一个LH顺序。在最后一个VH顺序的下游有一组D顺序(D意为多样性)。目前还不能确定D顺序的数目,据Leder(1982)估计约有50个。在D顺序下游有6个JH顺序。JH顺序下游则为一组CH顺序;在小鼠是Cμ、Cδ、Cγ3、Cγ1、Cγ2b、Cγ2α、Cε和Cα;在人有Cμ(可能有Cμ1和Cμ2)、Cδ、Cγ4、Cγ、2、Cγ3、Cγ1、Cs、Cα1和Cα2,但在Cδ之后的排列次序尚未肯定。重链基因的体细胞重组,是将VH顺序之一、D顺序之一和JH顺序之一(随同其下游CH顺序)连接起来,是为VH/D/JH重组(图2)。重组后仍存在间隔顺序。每个CH顺序也是分割的,各有DNA片段为重链各个功能区和轻链区编码。重链基因的原初转录产物也要经过拼接,剪除间隔顺序,才成为成熟的重链mRNA。
在淋巴细胞分化过程中,首先是重链基因发生VH/D/JH重组,形成重链活性基因,转录后再拼接,产生μ重链
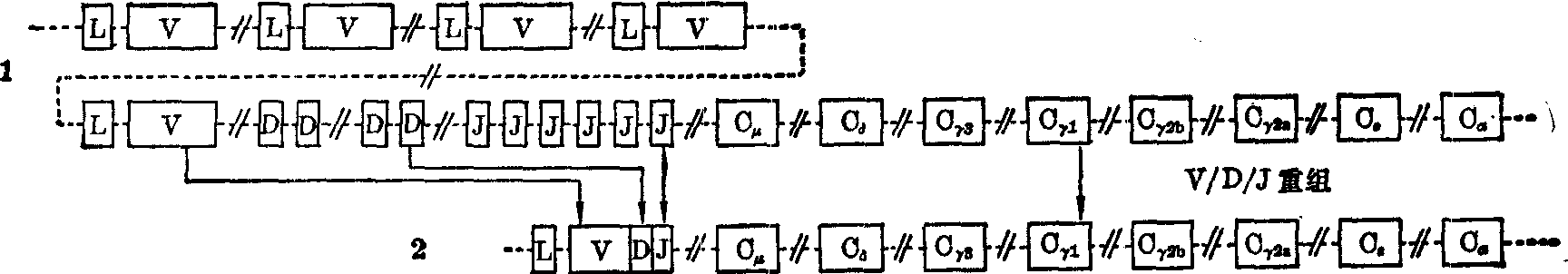
图2 小鼠重链活性基因的形成(各个Cn顺序分割为3~6个片段,图中未示出)
1. 种系细胞和胚胎细胞的重链遗传结构 2. 体细胞重组后形成的重链活性基因
同一个淋巴细胞可以产生附着在细胞膜上的IgM,也可产生分泌型IgM。附着在膜上的IgM,在其μ重链羧基端有一个嫌水性肽段,这在分泌型IgM的μ重链是没有的。现知两种μ重链的mRNA来自同一活性基因的差别转录(图3)。Cμ顺序分割为6个编码片段,为原初转录产物包括这6个编码片段,合成的μ重链其羧基端有嫌水性片段,产生的IgM可附着在细胞膜上。如原初转录产物只包括Cμ顺序前4个编码片段,合成的μ重链其羧基端无嫌水性肽段,产生的是分泌型IgM。很可能其他重链的CH顺序也有类似结构。
一个淋巴细胞可在产生IgM的同时产生IgD,也是由
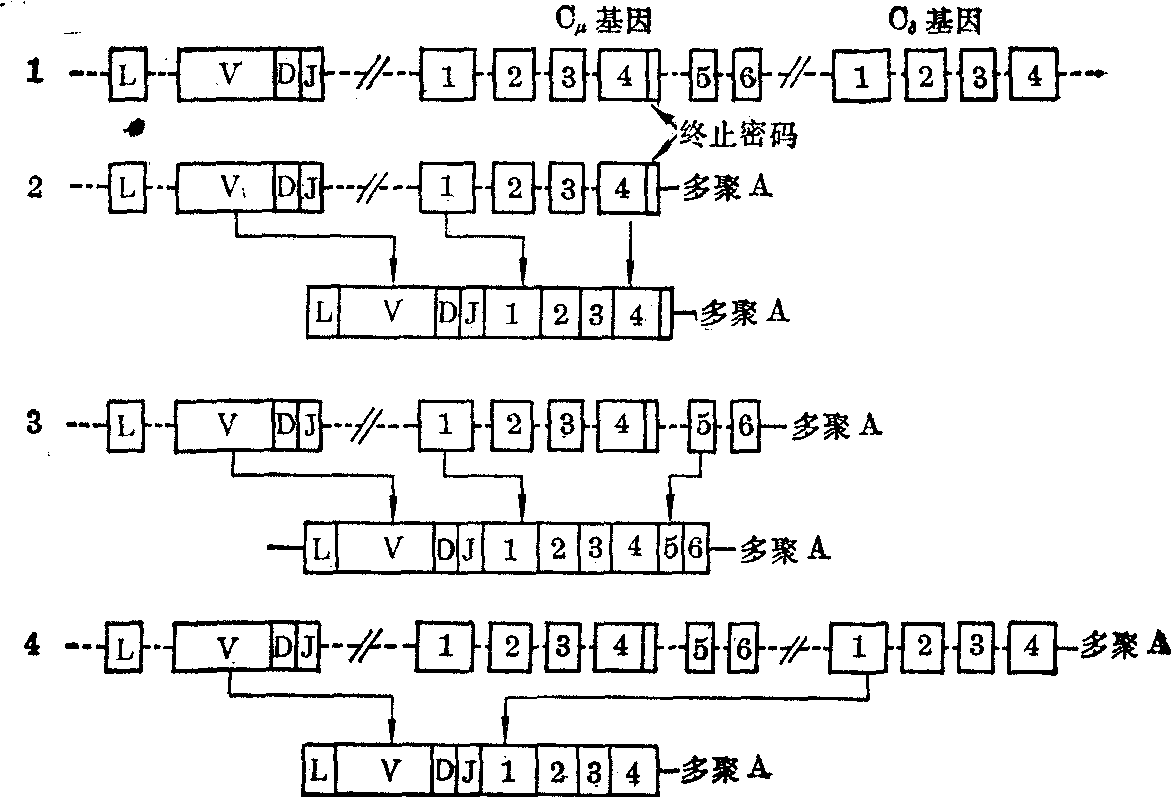
图3 重链活性基因的差别转录和原初转录产物的拼接
1. 重链活性基因 2. 原初转录产物和分泌型μ重链mRNA 3. 原初转录产物和膜附着型μ重链mRNA 4. 原初转录产物和δ重链mRNA
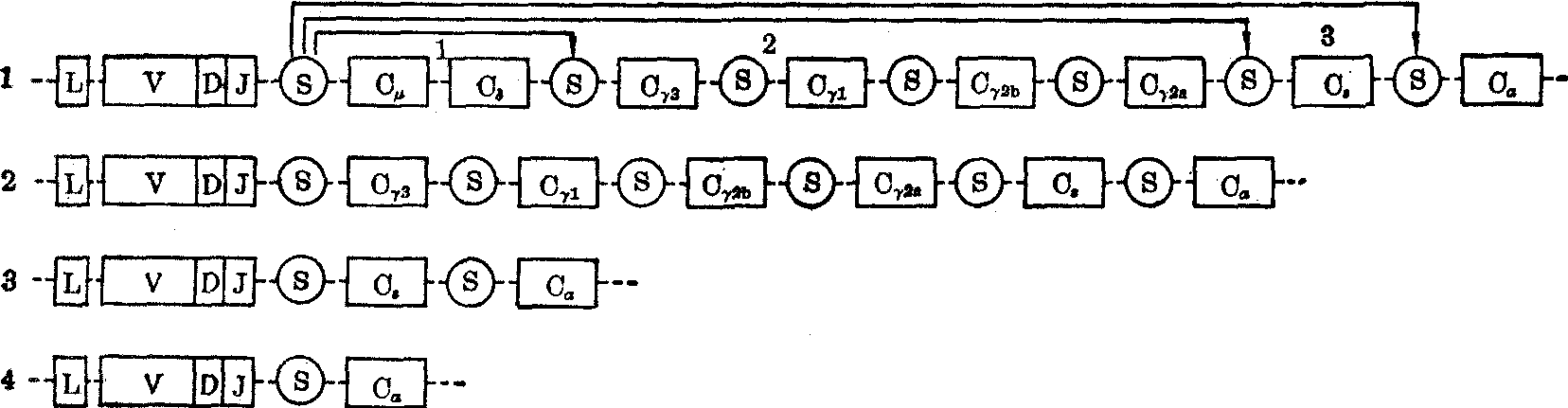
图4 重链活性基因通过有互补关系的信号顺序(S)发生重组,导致重链类别转换
1. 转换前的活性基因,细胞合成μ重链和δ重链 2. 转换后的活性基因,细胞合成γ3重链 3. 转换后的活性基因,细胞合成ε重链 4. 转换后的活性基因,细胞合成α重链
一个淋巴细胞先产生附着在膜上的IgM和IgD。在它被抗原选出后可转而产生分泌型IgM、IgG、IgE和IgA中的一种。这里涉及重链类别转换(图4)。现知重链类别转换是由于重链活性基因再次发生重组。试以小鼠为例,在Cγ3、Cγ1、Cγ2b、Cγ2a、Cε及Cα顺序前面的间隔顺序中都有一个顺序与VH/D/JH和Cμ…Cδ之间的一个顺序有某种互补关系,构成S信号(S意为转换)。其重组机理与VH/D/JH重组相似。因此,一个淋巴细胞克隆只产生一种特定类别的免疫球蛋白分子。
重链比轻链有更高的多样性。据估计,人的VH顺序多达80个,D顺序约有50个,JH顺序有6个,在体细胞重组时VH/D接头和D/JH接头各有10种不同可能性。重链的组合类型将有80×50×6×10×10=2,400,000种。
一个免疫球蛋白分子由两条相同轻链和二条相同重链构成。据Leder(1982)估计,免疫球蛋白分子的组合可能性将有7500×2,400,000=180亿种。
导致免疫球蛋白多样的机理,除上述种系编码顺序的组合机理之外,还有体细胞突变的作用。在成熟淋巴细胞中筛选新的抗原结合特异性,发现免疫球蛋白的遗传结构是高度不稳定的,每代每1万个细胞就有一次突变。还发现,从产生抗体的细胞中分离出来的活性基因,可与胚胎DNA中的相应顺序相差1~2个核苷酸。免疫球蛋白的潜在多样性将超过180亿。
等位基因排斥 等位基因排斥是免疫球蛋白的一种特殊遗传现象。一个淋巴细胞克隆只产生一种免疫球蛋白分子。人体等位基因是成对存在的,因为同源染色体是一对。在一对重链等位基因中只有一个等位基因发生体细胞重组而获得表达,其另一等位基因通常不发生重组也就不表达。但是偶尔也有两个等位基因都发生重组的。这可能是一次重组失效,随即使其同源染色体上的等位基因发生重组而成为活性基因。在轻链基因,κ基因重组在先,如κ基因重组失效,于是λ基因发生重组。人体在一条第2号染色体上有1个Cκ顺序,在一条第22号染色体上有6个Jλ/Cλ顺序,一个细胞共有14个轻链恒定区顺序,即有14次形成一个有活性的轻链基因的机会。
- 体育代表团是什么意思
- 体育代表队是什么意思
- 体育任务是什么意思
- 体育仿生学是什么意思
- 体育传媒是什么意思
- 体育传统项目学校是什么意思
- 体育伦理学是什么意思
- 体育保健学是什么意思
- 体育俱乐部是什么意思
- 体育先进县是什么意思
- 体育先进县市是什么意思
- 体育具体帮助法是什么意思
- 体育出版社是什么意思
- 体育分卷是什么意思
- 体育分篇是什么意思
- 体育分解教学法是什么意思
- 体育刊物是什么意思
- 体育制度是什么意思
- 体育功能是什么意思
- 体育医学检查是什么意思
- 体育医学监督是什么意思
- 体育医生是什么意思
- 体育医疗是什么意思
- 体育协会是什么意思
- 体育博士是什么意思
- 体育卫生是什么意思
- 体育卫生保健是什么意思
- 体育卫生谚语是什么意思
- 体育原理是什么意思
- 体育发展基金是什么意思
- 体育口号是什么意思
- 体育古今是什么意思
- 体育史是什么意思
- 体育名星瓦代尔误入歧途是什么意思
- 体育后备人才培训是什么意思
- 体育周刊是什么意思
- 体育周报是什么意思
- 体育哲学是什么意思
- 体育器材是什么意思
- 体育土器材制作的技能是什么意思
- 体育地理学是什么意思
- 体育场是什么意思
- 体育场地利用率奖是什么意思
- 体育场地器材的管理技能训练是什么意思
- 体育场馆是什么意思
- 体育场馆▷是什么意思
- 体育场馆设施是什么意思
- 体育基础理论是什么意思
- 体育基础理论与学科是什么意思
- 体育增刊是什么意思
- 体育夏令营是什么意思
- 体育大全是什么意思
- 体育大辞典是什么意思
- 体育奖是什么意思
- 体育奖励制度是什么意思
- 体育委员是什么意思
- 体育委员会是什么意思
- 体育学是什么意思
- 体育学习动机是什么意思
- 体育学院是什么意思