主要组织相容性复合体
简称“MHC”。脊椎动物中一组紧密连锁的基因构成的复合座位。包括一些为细胞表面抗原编码的以及为血清补体成分编码的基因,它们控制的免疫应答和疾病易感性。MHC具有高度的多态性。与器官移植和疾病遗传有密切关系,并为遗传学研究提供了极好的遗传标志。
主要组织相容性复合体
主要组织相容性复合体(MHC)是决定主要移植抗原并严重影响移植物排斥的一个遗传区段,或称超基因,如小鼠H-2系统及人类白细胞抗原(HLA)系统。高等动物和人体主要组织相容性复合体是在研究异体移植物的排斥中发现的。如果随机取一个人的皮肤移植于另一个人,移植皮肤只能存活10~15天,如果再次移植同一供者的皮肤,则移植皮肤将更快被排斥。这说明同种异体移植反应是一种特异性免疫反应。受者免疫系统能识别移植物为非己,从而引发一系列免疫反应将移植物排斥。移植物排斥的免疫学性质首为Medawer所确定。
早在1931年,研究红细胞血型的奠基人Landsteiner,就指出输血和移植配型是密切平行的,他提倡应用血清学方法寻找对移植有重要作用的血型。Gorer(1936)开始用小鼠研究与移植有关的抗原,结果发现小鼠的组织相容性H-2系统。此系统揭示的第一个抗原只是小鼠的一个红细胞抗原。不久Medawar用兔做实验,发现用供者白细胞使受者预先致敏,会使移植皮肤象再次移植那样被加速排斥,而红细胞却并无此作用。他推断要找出与移植有关的遗传差别,就应该集中研究白细胞而不是红细胞。后来Gorer和Snell等用小鼠近交系进行研究,Snell把这些与移植有关的白细胞抗原称为组织相容性抗原。现在知道H-2抗原是最重要的移植抗原,而且不只受控于一个基因位点,所以免疫遗传学家们把它称为主要组织相容性复合体。
以小鼠的H-2系统为模式,近二十年来不仅对人体的主要组织相容性复合体HLA系统有相当深入的研究,至少还在其他七种哺乳动物和一种鸟类中鉴定了主要组织相容性系统(表1)。大量数据表明,一卵双生之间移植皮肤或肾脏,其存活期要比一般同胞之间随机移植长得多。通过主要组织相容性复合体作组织配型,确可使临床器官移植,尤其是肾脏移植取得显著进展;但是克服移植物排斥的问题并未解决,因为即使在HLA相同的同胞间作肾脏移植也不能不用免疫抑制剂,否则移植肾将被排斥。免疫遗传学家们发现,主要组织相容性复合体具有高度的多态性,即各个位点上发生多次突变,形成很多等位基因,为遗传学研究提供了极好的遗传标志;他们还发现组织相容性抗原与免疫活性细胞之间的相互作用,从而揭示了主要组织相容性复合体在免疫反应中的重要作用。
表1 人和高等动物的主要组织相容性复合体
| 物种 | 主要组织相容性复合体 |
| 人 黑猩猩 猕猴 狗 猪 兔 豚鼠 大鼠 小鼠 鸡 | HLA ChL-A RhL-A DL-A SL-A RL-A GpL-A AgB H-2 B |
小鼠H-2系统 小鼠H-2系统(H-2复合体)的一系列连锁基因位点在小鼠第17号染色体中部,它的相对长度按所取边界不同而在0.3~1.5厘摩(cM)之间。在着丝粒端H-2K位点和端粒端H-2D位点之间为0.3cM。现知H-2D右侧的若干位点,在功能上和生化上至少与H-2系统有些位点有关,也可视为H-2系统的一部分,这样H-2系统就包括从着丝粒端H-2K位点到端粒端T1a位点(胸腺白血病抗原位点),其相对长度为1.5cM(图1)。
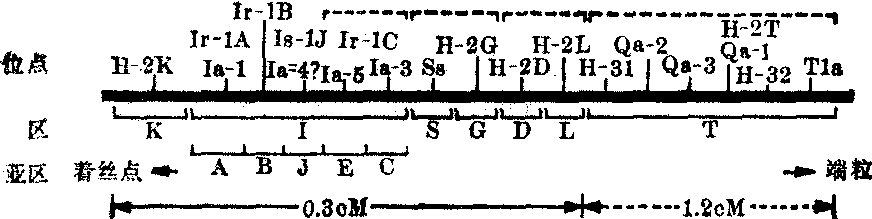
图1 小鼠H-2复合体遗传图
(虚线括号内位点的图序未定)。表示各区及其亚区中各个基因位点的相对位置
免疫遗传学家把至少检出一个位点但可能含有更多位点的染色体节段称为区。一个已鉴定几个位点的区可再分为若干亚区。不过区和亚区的区别相当含糊,有人建议不用亚区这一术语。迄今H-2遗传图分为K、I、S、G、D、T六个区,I区又再分为A、B、J、E、C五个亚区。K区和D区 H-2系统K区和D区的标记位点分别为H-2K和H-2D。H-2K位点和H-2D位点有高度多态性,迄今已发现H-2K位点有56个等位基因,H-2D位点有45个等位基因,可组成2520种基因组合方式。H-2K和H-2D的基因产生经典的移植抗原,这些抗原为糖蛋白,位于细胞膜上。实验证明每一抗原系由若干高度关联的抗原因子或称抗原决定簇组成。这提示为抗原分子编码的H-2K位点或H-2D位点内部可有多处发生突变,各决定一种特异抗原因子的形成。因此一个抗原分子可与二种或二种以上特异性抗血清起反应。这些抗原因子或为若干基因产物所共有,或为某一基因产物所特有,前者称为公共特异性,后者称为私有特异性。H-2K和H-2D抗原赋予效应淋巴细胞以效应特异性,产生一种现象,称H-2限制。小鼠在被病毒感染时可产生效应淋巴细胞,后者可在体内、体外破坏被病毒感染的细胞。效应淋巴细胞具有病毒特异性。例如被淋巴细胞性脉络丛脑膜炎病毒感染的小鼠所产生的效应细胞,只能破坏被淋巴细胞性脉络丛脑膜炎病毒感染的细胞,而不能破坏被其他病毒感染的细胞。效应淋巴细胞还同时有靶细胞特异性。例如一个针对感染了牛痘病毒的H-2a细胞的效应淋巴细胞只会破坏被牛痘病毒感染的H-2a细胞,而不能破坏被牛痘病毒感染的非H-2a细胞。即效应淋巴细胞的效应受H-2K和H-2D的限制。这提示一个正在成熟的淋巴细胞在学会识别病毒抗原的同时,识别了带有病毒抗原细胞的H-2K和H-2D抗原分子。Zinkernagel等利用胸腺上皮为H-2a型而骨髓为H-2b型的嵌合体小鼠,发现效应淋巴细胞学会识别被病毒感染了的H-2a细胞,这就证明H-2K和H-2D限制效应淋巴细胞特异性是由胸腺上皮决定的。
H-2K和H-2D抗原分子均由非共价连接的重链和轻链组成。重链是H-2K或H-2D的基因产物,分子量44,000,有二个多糖侧链,肽链约有350个氨基酸残基,其顺序分析接近完成,血清学特异性决定于肽链的氨基酸顺序。轻链为β2微球蛋白,长约100个氨基酸,分子量12,000,没有多糖侧链,其编码基因不在H-2系统中。β2微球蛋白还以游离状态存在于血清及其他一些体液中。小鼠H-2K和H-2D抗原重链氨基端的顺序分析表明,H-2K和H-2D抗原分子的一级结构有高度同源关系(表2)。这表明这些基因可能系由原始基因重复而来。
表2 小鼠H-2K和H-2D及人的HLA-A和HLA-B抗原重链氨基端的顺序(Klein,1979)

“一”与H-2Kb的氨基酸残基相同; “·”未定: “( )”未肯定。
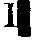 区 Ⅰ区又被划分为A、B、 J、E、C五个亚区(图1)。由于检测表型的指标不同,检出Ⅰ区两种基因,即免疫反应基因(Ir)和免疫反应相关基因(Ia)。诱发免疫反应的过程是很复杂的,遗传调节可发生在不同水平上:Ir基因具有高度特异性,在体液免疫水平上控制动物产生抗体,又在细胞免疫水平上表现为迟发型免疫反应和体外T细胞增殖反应。在某些低反应品系中,发现有些合成多肽抗原会诱导抑制抗体反应的T抑制细胞。这一抑制活性也是特异的,也受Ir基因控制。
区 Ⅰ区又被划分为A、B、 J、E、C五个亚区(图1)。由于检测表型的指标不同,检出Ⅰ区两种基因,即免疫反应基因(Ir)和免疫反应相关基因(Ia)。诱发免疫反应的过程是很复杂的,遗传调节可发生在不同水平上:Ir基因具有高度特异性,在体液免疫水平上控制动物产生抗体,又在细胞免疫水平上表现为迟发型免疫反应和体外T细胞增殖反应。在某些低反应品系中,发现有些合成多肽抗原会诱导抑制抗体反应的T抑制细胞。这一抑制活性也是特异的,也受Ir基因控制。小鼠H-2内部重组品系实验表明有四个Ir基因位点,其中三个为增强免疫反应位点,位于A(Ir-1A位点)、B(Ir-1B位点)和E/C(Ir-1C位点)三个亚区内;一个是抑制免疫反应的位点,位于J亚区(Ir-1J)。控制混合淋巴细胞培养 (MLC) 反应的ID-1位点与A亚区密切连锁,或是包含于其中。此外尚有一种或几种有关MLC反应的因子受控于E/C亚区。Ia基因产生B细胞和巨噬细胞上的特异性Ia抗原,至少已检出21种Ia特异性。Ia抗原为糖蛋白,系由两条非共价结合的多肽链构成,即α链(分子量35,000)和β链(分子量28,000),另有数目不详的糖链。是否α链和β链均受控于H-2复合体,尚有待确定。
对胸腺依赖性抗原的抗体反应,需有T辅助细胞参加,因为需要T和B细胞两者的I区具有相同特异性,配合作用后,才能使B细胞增殖分化为浆细胞,从而产生较多的抗体球蛋白。T辅助细胞在胸腺上皮影响下获得识别I抗原和外源抗原(如病毒抗原)的能力。由此可见,H-2限制包括K区和D区限制效应细胞特异性和I区限制辅助细胞特异性。
目前明确的Ia位点有2~3个。Ia-1位点在A亚区。Ia-5和Ia-3位点则分别在E和C亚区,但看法尚不一致。Ia-4位点在J亚区,决定T抑制细胞上的Ia抗原。A亚区Ia位点象D区和K区那样多态,E/C区Ia位点只检出五个等位基因。
S区 S区的基因产物是血清中的一类特殊β球蛋白,由兔抗小鼠血清检出者称为血清蛋白质(Ss),由同种异体抗血清检出者称为
 蛋白质(Slp),两者从未发生重组。事实上Slp仅为Ss上的一个抗原决定簇。现知Ss为补体C4,分子量200,000,由三条共价连接的多肽链组成,即α链(95,000)、β链(78,000)和γ链(33,000),三链均由一条前体链分裂而来,推断为单一基因编码。最近Roos等(1978)报道Ss和Slp的γ链有微小差异,提示或许受控于两个连锁基因。H-2G区基因产物是H-2.7抗原,是小鼠的一种红细胞抗原,但G区是否独立于Ss-Slp尚不肯定。H-2T区中的几个位点尚未肯定,其代表性位点是为小鼠胸腺白血病抗原编码的Tla。综上所述,H-2遗传图据保守的估计有10个位点,据不保守的估计至少有18个位点。
蛋白质(Slp),两者从未发生重组。事实上Slp仅为Ss上的一个抗原决定簇。现知Ss为补体C4,分子量200,000,由三条共价连接的多肽链组成,即α链(95,000)、β链(78,000)和γ链(33,000),三链均由一条前体链分裂而来,推断为单一基因编码。最近Roos等(1978)报道Ss和Slp的γ链有微小差异,提示或许受控于两个连锁基因。H-2G区基因产物是H-2.7抗原,是小鼠的一种红细胞抗原,但G区是否独立于Ss-Slp尚不肯定。H-2T区中的几个位点尚未肯定,其代表性位点是为小鼠胸腺白血病抗原编码的Tla。综上所述,H-2遗传图据保守的估计有10个位点,据不保守的估计至少有18个位点。人体HLA系统 人体HLA系统(HLA复合体)是小鼠H-2系统的同源结构,受控于人体第6号染色体短臂的HLA 区域(6p2100—6pter)(图2)。
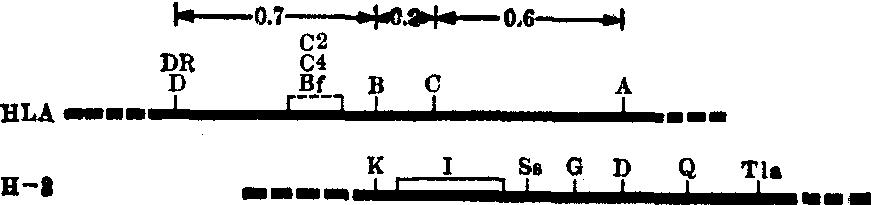
图2 人体HLA和小鼠H-2遗传图比较
1952年,Dausset和Nenna在多次输血病人的血清中发现白细胞凝集素。1958年,Dausset利用病人血清检出第一个白细胞抗原Mac (=HLA-A2+A28)。同年Payne等及van Rood在经产妇血清中,发现了由胎儿和母亲不相容产生的白细胞凝集素,开拓了白细胞抗血清的广阔来源。白细胞抗原研究的迅速发展,要归功于vanRood的重要贡献。他于1962年,将数理统计中的聚类分析和电子计算机引进白细胞抗血清特异性分析,建立了4a/4b等位基因系统,即Four系统。1964年,Payne等进一步发展了这些统计方法,检出了等位抗原LA1(=HLA-A1)和LA2(=HLA-A2),即LA系统。1964年,Walford等将检查小鼠白细胞抗原的补体依赖细胞毒性试验移用于人体,并确定新鲜兔血清为该试验的最佳补体来源。同年,寺畸(Terasaki)和McClelland又将此细胞毒试验微量化,并在经产妇血清中检出了细胞毒抗体。此后在经产妇血清中筛选细胞毒抗体,成为白细胞SD抗原分型血清的主要来源,推动了这一领域的迅速发展。1970年Thorsby等鉴定了AJ抗原。遗传分析表明AJ抗原独立于Four和LA系统。随后检出的许多白细胞抗原均分属于LA、Four和AJ三个分离子系列,分别受控于HLA-A、HLA-B和HLA-C三个连锁的位点,每个位点包括许多个等位基因。
1964年 Bain、Bach和Hirschhorn将两个人的淋巴细胞混合培养,发现淋巴细胞转化为淋巴母细胞样细胞的程度与两个个体的组织相容程度成反比。随后,Bach、Yunis、Amos等人发现MLC反应受控于与SD抗原基因连锁的独立位点,称为MLR-S位点,即HLA-D位点。由MLC检出的特异性可称为细胞学鉴定抗原,即LD抗原。
1971年Ceppellini等发现有些HLA分型血清可抑制MLC反应,此种抑制活性不能由血小板吸收去除。随后van Leeuwen等(1973)系统地寻找能阻断MLC的血清,并由免疫荧光技术证明,此种有阻断作用的血清中的抗体特异地与B细胞反应,而不与T细胞反应,提示人体B细胞上另有一组可由血清学方法检出的小鼠Ia样抗原。
随着检出的白细胞抗原越来越多,就需要统一命名。1967年Allen等建议把决定人体白细胞抗原的主要位点称为HL-A,这为以后成立的世界卫生组织命名委员会所接受。1975年命名委员会规定以HLA表示人体白细胞抗原的遗传区域或系统,下接联字号和拉丁字母以表示位点,再以数字编号表示抗原。凡属专题讨论会临时鉴定的抗原,于所属位点字母后加W(workshop),待其最后被证实时除去W。HLA系统知识的迅速积累是全世界免疫学家通力协作的结果。1964~1980年已召开了八届国际组织相容性试验专题讨论会议。
研究HLA的动力主要来自临床器官移植配型的需要。20年来的研究成果表明,HLA配型对于延长移植物存活相当重要,而且HLA系统是迄今所知最多态的免疫遗传系统,故可作为遗传标志,用来研究疾病的遗传机理,从而导致研究HLA与疾病关联的新领域。
HLA-A、B、C位点 HLA-A、B、C位点均为共显性复等位基因位点,它们的基因产物均为SD抗原,各由高度相关的细胞毒血清组按全部或多数血清反应作鉴定。截至1980年2月第八届国际组织相容性专题讨论会,已鉴定A位点特异性20个,B位点特异性42个,C
表3 HLA系统特异性命名表(1980年2月)
| HLA-A | HLA-B | HLA-C | HLA-D | HLA-DR |
| A1 A2 A3 A9 Aw23 Aw24 A10 A25 A26 A11 A19 A29 Aw30 Aw31 Aw32 Aw33 A28 Aw34 Aw36 Aw43 | B5 Bw51 Bw52 B7 B8 B1Z Bw44 Bw45 B13 B14 B15 Bw62 Bw63 Bw16 Bw38 Bw39 Bw17 Bw57 Bw58 B18 Bw21 Bw49 Bw50 Bw22 Bw54 Bw55 Bw56 Bw27 Bw35 Bw37 Bw40 Bw60 Bw61 Bw41 Bw42 Bw46 Bw47 Bw48 Bw53 Bw59 Bw4 Bw6 | Cw1 Cw2 Cw3 Cw4 Cw5 Cw6 Cw7 Cw8 | Dw1 Dw2 Dw3 Dw4 Dw5 Dw6 Dw7 Dw8 Dw9 Dw10 Dw11 Dw12 | DR1 DR2 DR3 DR4 DR5 DRw6 DR7 DRw8 DRw9 DRw10 |
表中缩进一格者是上一抗原的分解抗原
位点特异性8个。A位点和B位点的特异性按位点分类、依数字连续编号: Bw4原来为4a,Bw6原来为4b,编号20原为AJ抗原,现已列入C位点特异性重新编号(表3)。
HLA-A、B、C抗原存在于T细胞、B细胞、血小板、精子及其他所有有核细胞的细胞膜上。HLA-A、B、C抗原均为糖蛋白,由重链和轻链作非共价连接(图3)。重链分子量43,000,具有特异性抗原决定簇。轻链分子量12,000,即β2微球蛋白,无特异性,为HLA-A、B、C抗原所共有,其编码基因在第15号染色体。HLA-A、B抗原重链N端氨基酸顺序有显著同源关系,提示它们起源于共同的原始基因(表2)。
许多HLA-A、B抗原已经被分解,如A9分解为Aw23和Aw24,A10分解为A25和A26,B5分解为Bw51和Bw52等。这意味着A9、A10、A19、B5、B12、Bw16、Bw21等决定簇各为它们相应分解物的抗原分子所共有,前者可称为超型特异性,其分解物属于亚型特异性。Bw4和Bw6即van Rood发现的4a和4b。已有证据表明,Bw4和Bw6特异性存在于有其他B位点特异性的抗原分子上,分别与某些B位点特异性及其分解物相关联,可用于鉴别分解物。如Bw4与Bw38(Bw16)、Bw44(B12)与Bw49(Bw21)相关联,Bw6与Bw39 (Bw16)、Bw45(B12)与Bw50(Bw21)相关联。有些HLA-A、B特异性多见于特殊人群,如Aw36、Aw43和Bw42多见于非洲黑人,Aw34见于一些黑人和东方人,Bw46 (SIN2)和Bw54(Bw22J)见于东方人,Bw48(KSO)见于爱斯基摩人、印第安人及某些东方人。根据上海市免疫研究所调查,汉族人HLA-A、B、C、DR、D位点基因频率见表4。
图3 HLA-ABC抗原的分子结构
表4 我国汉族人HLA抗原基因频率(上海市免疫学研究所)
| HLA-A位点 | HLA-B位点 | HLA-C位点 | HLA-DR位点 | HLA-D位点 | |||||
| A1 A2 A3 A9 A10 A11 Aw19 A29 Aw30 Aw31 Aw32 Aw33 合计 空白 | 0.0272 0.2441 0.0180 0.2681 0.0090 0.2094 0.2324 0.0180 0.0457 0.0551 0.0180 0.0893 1.0000 0.0000 | B5 Bw51 Bw52 B7 B8 B12 Bw44 Bw45 B13 B15 Bw62 Bw63 B15/CH B16 | 0.0937 0.0646 0.0272 0.0090 0.0090 0.0180 0.0090 0.0090 0.1136 0.1763 0.1136 0.0090 0.0457 0.0272 | Cw1 Cw2 Cw3 Cw4 Cw5 Cw6 Cw7 Cw8 合计 空白 | 0.1679 0.0000 0.3645 0.1013 0.0000 0.1451 0.1340 0.0000 0.9128 0.0872 | DR1 DR2 DR3 DR4 DR5 DRw6 DR7 DRw8 DRw9 DRw10 合计 空白 | 0.0000 0.1964 0.0871 0.1461 0.0871 0.0000 0.1220 0.0646 0.1964 0.0000 0.8997 0.1003 | Dw1 Dw2 Dw3 Dw4 Dw5 Dw6 Dw7 Dw8 Dw9 Dw10 Dw12 合计 空白 | 0.0093 0.0871 0.0093 0.0187 0.0671 0.0093 0.0572 0.0000 0.0000 0.0000 0.0093 0.2673 0.7327 |
(续表)
| HLA-A位点 | HLA-B位点 | HLA-C位点 | HLA-DR位点 | HLA-D位点 | |
| Bw38 Bw39 B17 Bw58 Bw59 Bw21 Bw22 Bw54 Bw55 Bw56 B27 Bw35 B37 B40 Bw60 Bw61 Bw41 Bw42 Bw46 Bw47 Bw48 Bw59 合计 空白 | 0.0090 0.0180 0.0937 0.0180 0.0742 0.0090 0.1237 0.0551 0.0457 0.0180 0.0090 0.0839 0.0090 0.1872 0.1136 0.0646 0.0000 0.0000 0.0551 0.0000 0.0000 0.0000 1.0000 0.0000 | ||||
某些HLA-A位点基因和HLA-B位点基因组合的单倍型,其频率显著超过随机预期频率,这一现象称为连锁不平衡。最显著的连锁不平衡在白人中见于单倍型A1、B8和A3、B7。
HLA-D和HLA-DR位点 HLA-D位点的基因产物是LD抗原或称为淋巴细胞激活决定簇,系由单向混合淋巴细胞培养法检出。检测方法有阴性分型和阳性分型两种。阴性分型为经典方法。首先由HLA-A、B分型在近亲婚配子女中找出HLA纯合细胞,这些细胞经交叉单向MLC按阴性反应(一般为低反应,但可与阳性反应鉴别,有时称为分型反应)聚类后作为特异性纯合分型细胞组。凡未知淋巴细胞对经X线照射或丝裂霉素处理的纯合分型细胞呈阴性反应,即表示有此特异性。阳性分型系先以相差一条HLA单倍型的两个人的淋巴细胞分别作单向MLC,得到特异性致敏淋巴细胞。经X线照射或丝裂霉素处理的未知细胞与这些预先致敏的淋巴细胞作单向MLC时,阳性反应表示有特定LD抗原。截至1980年2月检出LD抗原Dw1~Dw12,其中Dwll多少包含于Dw7,可能是Dw7的一个分解物。
HLA-DR抗原是相
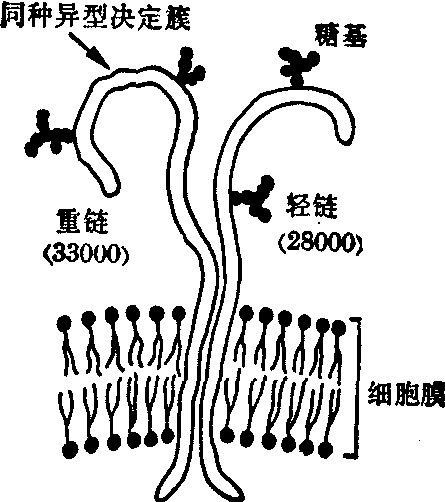
图4 HLA-DR抗原的分子结构
当于小鼠Ia抗原的分子,存在于B细胞和单核细胞,不存在于T细胞、血小板和肝细胞。DR抗原由经血小板吸收而去除A、B、C抗体的特异性血清检出。DR抗原与D抗原高度相关,DR(D-relat-ed)即表示与D有关,两者迄未发现重组,很可能为同一产物,故有合写为D(R)抗原。DR抗原亦为糖蛋白,由重链(分子量33,000)和轻链(分子量28,000)作非共价连接而成。两链均有多糖残基(图4)。重链具有特异性决定簇,其编码顺序在HLA区域,轻链编码顺序可能不在HLA区域,但尚无实验证据。人体补体成分C2、C4和Bf位点也位于HLA区域。通过HLA配型为肾脏移植选择供受组合,在亲属肾移植是简便有效的方法。由于HLA诸位点是密切连锁的(A和B之间的交换率为0.8%,B和C之间的交换率为0.2%,B和D之间的交换率为0.7%),碰到重组体的机会不多。设父母精子和卵子的HLA分别为ab和cd,子女中便有四种可能组合,即ac、ad、bc、bd。如第一胎为ac,则第二胎为HLA ac或bd的概率各为0.25,为HLA ad、bc的概率为0.50。子女的HLA只有一半与父或母相同。因此,在配型时应首选HLA相同同胞,次选HLA半相同的同胞或父母,对于HLA不相同的同胞,没有任何理由去冒险。尸体肾则选择有尽可能多的相同抗原的受者。
骨髓含有免疫活性细胞,移植骨髓可导致移植物抗宿主病,因此骨髓移植迄今只能在HLA相同同胞间进行。HLA与疾病的关联 比较病人组和对照组的HLA抗原频率,发现某些疾病与特定的HLA抗原显著关联,主要是B位点抗原和D (R)抗原。一些最突出的发现见表5。表中相对风险超过1,表示有某抗原者得该种疾病的风险为无此抗原者的若干倍。HLA抗原与疾病关联的机理尚未了解,一般认为并非由于HLA抗原本身,很可能是由于与特定HLA-B、D(R)基因处于连锁不平衡的某种免疫反应基因或疾病易感基因。将病人组有关抗原频率与按显性遗传和隐性遗传所得的群体遗传模型的理论值相比较,Kidd等(1977)得出假设的强直性脊椎炎易感基因为显性基因。同年,Thomson和Bodmer得出假设的青年型糖尿病易感基因为隐性基因(表5)。
表5 HLA与疾病的关联
| 疾 病 | 相关抗原 | 频率(%) | 相对风险 | |
| 对照组 | 病人组 | |||
| 乳糜泻(白人) | B8 Dw3 | 20 27 | 67 96 | 8.1 64.5 |
| 强直性脊椎炎 Reiter病 | B27 B27 | 8 9 | 90 80 | 103.5 40.4 |
| 慢性活动性肝炎 | B8 DRw3 | 15 17 | 52 41 | 6.1 3.4 |
| 重症肌无力(白人) | B8 DRw3 | 17 17 | 39 32 | 3.1 2.3 |
| 重症肌无力(日本人) | DRw4 | 35 | 59 | 2.7 |
| Graves病(白人) | B8 DRw3 | 18 18 | 44 53 | 3.6 5.1 |
| 风湿样关节炎(白人) | DRw4 | 15 | 56 | 7.2 |
| 青年型糖尿病(白人) | B8 DRw3 | 16 17 | 32 27 | 2.5 1.8 |
| 青年型糖尿病(日本人) 多发性硬化(白人) 血色素沉着症(白人) | DRw4 DRw2 A3 | 35 22 21 | 65 41 72 | 3.4 2.5 9.7 |
主要组织相容性复合体
主要组织相容性复合体 (MHC)是控制形成主要组织相容性抗原的基因群。MHC由多位点组成。其产物即为分布于有核细胞膜上的主要组织相容性抗原,称为主要组织相容性系统。现已证明,在多种动物体内含有主要组织相容性复合体。在小鼠为H-2,在人为HLA,猩猩为CHLA,恒河猴为RhLA,狗为DLA,猪为SLA,兔为RLA,豚鼠为GPLA,大鼠为AgB位点H-1,鸡为B位点。还有人认为两栖类也有类似系统,看来主要组织相容性复合体在进化过程中是发生较早的。目前对小鼠及人的基因复合体的研究较深入。
- 哥舒翰是什么意思
- 哥舒翰是什么意思
- 哥萨克是什么意思
- 哥谭镇是什么意思
- 哥达大会和《哥达纲领》批判是什么意思
- 哥达纲领批判是什么意思
- 哥达纲领批判是什么意思
- 哥达纲领批判是什么意思
- 哥达纲领批判是什么意思
- 哥达纲领批判是什么意思
- 哥达纲领批判是什么意思
- 哥雅是什么意思
- 哥顿法是什么意思
- 哥顿法是什么意思
- 哦是什么意思
- 哦是什么意思
- 哦热热是什么意思
- 哦罗斯是什么意思
- 哦美苦是什么意思
- 哦,小公马是什么意思
- 哦,香雪是什么意思
- 哦,香雪是什么意思
- 哨是什么意思
- 哨是什么意思
- 哨是什么意思
- 哨兵是什么意思
- 哨兵是什么意思
- 哨呐情话是什么意思
- 哨子是什么意思
- 哨岗社是什么意思
- 哨遍是什么意思
- 哨遍是什么意思
- 哨遍是什么意思
- 哨遍是什么意思
- 哨遍·高祖还乡〔般涉调〕是什么意思
- 哨遍(套数)是什么意思
- 哩是什么意思
- 哩是什么意思
- 哩依儿若是什么意思
- 哩哩是什么意思
- 哩嘎嘟是什么意思
- 哩多是什么意思
- 哩(里 裹)是什么意思
- 哪是什么意思
- 哪是什么意思
- 哪是什么意思
- 哪是什么意思
- 哪是什么意思
- 哪一壶不开提哪一壶是什么意思
- 哪一天庄稼汉不再买布吕赫尔和那愚蠢的英国贵族,而从集市上带回去别林斯基和果戈理是什么意思
- 哪个猫儿不吃腥是什么意思
- 哪咤是什么意思
- 哪咤是什么意思
- 哪咤是什么意思
- 哪咤是什么意思
- 哪咤(1)是什么意思
- 哪咤(2)是什么意思
- 哪咤下凡——一身火是什么意思
- 哪咤令是什么意思
- 哪咤传是什么意思